 2019, Vol. 39
2019, Vol. 39文章信息
- 庞碧剑, 蓝文陆, 黎明民, 李天深
- PANG Bijian, LAN Wenlu, LI Mingmin, LI Tianshen
- 北部湾近岸海域浮游动物群落结构特征及季节变化
- Community structure and seasonal variation of zooplankton in coastal Beibu Gulf
- 生态学报. 2019, 39(19): 7014-7024
- Acta Ecologica Sinica. 2019, 39(19): 7014-7024
- http://dx.doi.org/10.5846/stxb201807191553
-
文章历史
- 收稿日期: 2018-07-19
- 网络出版日期: 2019-08-16





北部湾近岸海域是我国重要的海产品增养殖区和优质渔场之一[1], 同时其拥有红树林、珊瑚礁和海草床等多种典型海洋生态系统, 是我国海洋生物多样性最高的海区之一[2]。但由于近十年来北部湾经济的快速发展和沿岸城市现代化的不断推进, 北部湾近岸海域的生态环境正承受着愈来愈大的压力, 局部海域污染问题突出, 环境容量下降, 海湾富营养化加剧, 海洋生态灾害如赤潮、绿潮等开始加重, 海湾生态景观遭受到不同程度的破坏。如何保护好北部湾优良的生态环境, 构建人与海洋和谐共生的蓝色海湾已成为迫切需求。
浮游动物是海洋生态系统中重要的次级生产者, 其群落结构的动态变化通过“下行效应”制约着浮游植物的群落结构, “上行效应”则影响着鱼、虾和贝类等海洋生物资源的结构和总量, 在海洋生态系统的物质循环和能量流动中起着重要的作用[3]。近年来, 有关北部湾海域浮游动物的种类组成和分布等已进行了部分的调查与监测。如郑白雯等研究了北部湾北部浮游动物数量分布及优势种[4];郑挺等研究了北部湾北部浮游动物空间生态位及其分化[5];阙江龙等研究了北部湾西北部饵料浮游动物季节变化及其与鱼卵、仔稚鱼的关系[6]。但这些研究主要集中在北部湾相对靠外的海域, 对于北部湾近岸海域尤其是20 m以内、人为扰动较大的沿岸海域浮游动物群落结构的研究仍很少, 只有个别小海湾如钦州湾有学者作了两次报道[7-8]。为此, 本研究拟聚焦于受人为影响较为严重的北部湾近岸海域, 对浮游动物群落组成、生物量和丰度的时空变化、优势种的季节变化、生物多样性等进行分析, 研究其主要影响要素, 以期为掌握北部湾近岸海域生态环境的变化特征以及生态保护提供基础资料, 同时也为深入研究该海域浮游动物与环境质量的内在关系提供科学依据。
1 材料与方法 1.1 样品采集与分析分别在2017年3月13日—24日(枯水期)、7月15日—31日(丰水期)和10月01日—28日(平水期)对北部湾近岸海域的浮游动物、叶绿素a和理化环境因子等进行了3个航次的综合调查, 共布设了44个调查站位(图 1)。各调查水文时期的站位水深介于0.8—20.0m之间, 其中水深最深出现在平水期的S23站位, 大多数站位水深小于10.0m。

|
| 图 1 采样站位 Fig. 1 Sampling stations in coastal Beibu Gulf |
浮游动物样品用浅水Ⅰ型浮游生物网(网口直径50 cm、筛绢CQ14、孔径0.505 mm)自海底至海面进行垂直拖网采集, 标本以5%福尔马林溶液固定保存, 湿重法称量其生物量, 在显微镜和体视镜下进行种类鉴定和计数。浮游动物样品的采集、保存和分析均按《海洋监测规范—海洋生物调查》(GB17378.7—2007)[9]的规定进行。
水温、盐度采用多参数水质仪(Multi 350)现场测定, 其他水质监测项目样品用8L采水器采集后带回实验室分析。水质样品的采集、保存、分析等均按《海洋监测规范—海水分析》(GB17378.4—2007)[10]提供的方法测定。
1.2 统计方法浮游动物优势度(Y)的计算公式为:
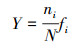
|
式中, N为样品的总丰度;ni为样品中第i种的丰度;fi为该种在各样品中的出现频率。若Y ≥0.02, 则认定该物种为优势种[11]。
浮游动物群落的Shannon-Wiener多样性指数(H′)[12]的计算公式为:

|
式中, H′为种类多样性指数;S为样品中的种类总数;Pi为第i种的个体数与总个体数的比值。
采用SPSS 16.0统计分析软件对浮游动物生物量与主要环境因子进行Pearson相关性分析。浮游动物丰度、生物量以及生物多样性指数分布图采用surfer11.0克里金(Kriging)插值法绘制。
2 结果 2.1 种类组成和生态类群北部湾近岸海域浮游动物种类比较丰富, 2017年已鉴定出终身性浮游动物251种和兼性浮游幼虫24个类群(表 1)。终身性浮游动物以桡足类的种类数最多;其他种类数较多的类群依次是水螅水母类、毛颚类和端足类, 种类数在17—68之间;介形类、被囊类、浮游螺类、糠虾类、管水母类、枝角类、樱虾类等种数较少, 均在8种以下。各水文时期皆有出现的浮游动物种类为55种, 仅占总种数的20%。浮游动物枯水期和丰水期的种类数相近, 平水期大幅度增加, 其种类数最多。浮游动物各类群的种数季节变化因类群不同而有所差异。桡足类、端足类的种类数季节变化幅度较大, 均以平水期种数最多, 而丰水期的种类数较少;浮游幼虫、水螅水母类则是平水期种数最多, 枯水期的种数较少;毛颚类、糠虾类、枝角类、磷虾类等大多数类群的种数季节性差异不大(表 1)。
| 浮游动物类群 Zooplankton group |
种类数Species amount | ||||
| 枯水期 Dry season |
丰水期 Wet season |
平水期 Normal season |
总计 Total |
占比/% Percentage |
|
| 桡足类Copepoda | 47 | 39 | 65 | 85 | 30.91 |
| 水螅水母类Hydromedusae | 17 | 35 | 43 | 68 | 24.73 |
| 毛颚类Chaetognatha | 21 | 18 | 19 | 28 | 10.18 |
| 浮游幼虫Pelagic larvae | 13 | 19 | 20 | 24 | 8.73 |
| 端足类Amphipoda | 6 | 0 | 13 | 17 | 6.18 |
| 介形类Ostracocda | 7 | 1 | 3 | 8 | 2.91 |
| 被囊类Tunicata | 7 | 3 | 4 | 8 | 2.91 |
| 腹足类Gastropoda | 2 | 4 | 6 | 7 | 2.55 |
| 糠虾类Mysidacea | 4 | 3 | 2 | 6 | 2.18 |
| 管水母类Siphonophora | 3 | 1 | 3 | 5 | 1.82 |
| 枝角类Cladocera | 2 | 3 | 2 | 4 | 1.45 |
| 十足类Decapoda | 1 | 4 | 3 | 4 | 1.45 |
| 多毛类Polychaeta | 3 | 1 | 3 | 4 | 1.45 |
| 栉水母类Ctenophora | 2 | 1 | 2 | 2 | 0.73 |
| 磷虾类Euphausiacea | 1 | 1 | 1 | 2 | 0.73 |
| 原生动物Protozoa | 1 | 1 | 1 | 1 | 0.36 |
| 涟虫类Cumacea | 1 | 0 | 0 | 1 | 0.36 |
| 钵水母类Scyphozoa | 0 | 0 | 1 | 1 | 0.36 |
| 合计Total | 138 | 134 | 191 | 275 | 100 |
北部湾近岸海域地处亚热带, 终年水温较高(16.8—33.1℃)。北岸有北仑河、防城江、茅岭江、钦江、南流江、大风江等多条河流淡水注入, 并且海域开阔, 受外海水的影响显著。该海域是咸淡水交汇的水域, 其盐度变化范围较广(0.9—32.6)。因此, 北部湾近岸海域的浮游动物群落结构较为复杂。根据浮游动物的生态习性及地理分布, 北部湾近岸海域的浮游动物可大致划分为下列4个生态类群[3-8]:
河口低盐性类群:受大陆径流的影响, 这片海域生活着一些适应低盐的种类, 主要有中华异水蚤(Misophria sinensis)、火腿伪镖水蚤(Schmackeria poplesia)、刺尾纺锤水蚤(Acartia spinicauda)。这些河口低盐种类在调查海域仅零星出现, 并多出现于盐度较低的丰水期。
近岸暖温性类群:适应相对低温高盐的种类, 种数不多, 主要有中华哲水蚤(Calanus sinicus)、强额孔雀水蚤(Pavocalanus crassirostris)、小拟哲水蚤(Paracalanus parvus)、近缘大眼水蚤(Corycaeus affinis)等, 这些是典型的低温高盐种类, 在调查海域多出现于水温较低的枯水期。
近岸暖水性类群:该类群主要出现在水温较高的丰水期和平水期, 种类数多, 分布广, 数量大, 为北部湾近岸海域浮游动物最主要组成部分, 主要物种有太平洋纺锤水蚤(Acartia pacifica)球型侧腕水母(Pleurobrachia globosa)、鸟喙尖头溞(Penilia avirostris)、肥胖三角溞(Evadne tergestina)、红纺锤水蚤(Acartia erythraea)、亚强次真哲水蚤(Subeucalanus subcrassus)、锥形宽水蚤(Temora turbinata)、中型莹虾(Lucifer intermedius)、百陶箭虫(Sagitta bedoti)、肥胖箭虫(Sagitta enflata)等。这些种类是典型的亚热带沿岸种。
外海暖水性类群:适应高温高盐的种类, 适应范围相对较广, 在调查海区出现的种类也较多。代表种有软拟海樽(Dolioletta gegenbauri)、四叶小舌水母(Liriope tetraphylla)、精致真刺水蚤(Euchaeta concinna)、不列颠高手水母(Bougainvillia britannica)、奥氏胸剌水蚤(Centropages orsinii)、多变箭虫(Sagitta decipiens)等。
2.2 丰度的平面分布与季节变化2017年北部湾近岸海域浮游动物丰度的变化范围为0—17676.67个/m3, 年均值为789.95个/m3。浮游动物丰度有明显季节变化, 枯水期丰度最高, 平均丰度高达1540.19个/m3, 平水期次之, 平均丰度为457.58个/m3, 丰水期最低, 平均丰度为372.08个/m3。枯水期浮游动物丰度高值区主要分布钦州湾外海域和北海市南部涠洲岛海域, 其中最高丰度出现在S24站位;防城港珍珠湾海域及茅尾海海域丰度相对较低, 最低丰度出现在S4站位(图 2枯水期)。丰水期浮游动物的丰度高值区出现在防城港湾海域, 最高丰度出现在S6站位;低值区主要分布在珍珠湾、钦州湾、廉州湾和涠洲岛周边海域, 其中最低丰度出现在S21站位(图 2丰水期)。平水期浮游动物的丰度平面分布相对较为均匀, 高值区主要分布在廉州湾和铁山港海域, 最高丰度出现在S35站位, 最低丰度出现在钦州湾茅尾海海域的S15站位(图 2平水期)。

|
| 图 2 北部湾近岸海域浮游动物丰度的平面分布/(个/m3) Fig. 2 Horizontal distribution of abundance of zooplankton in coastal Beibu Gulf |
2017年北部湾近岸海域浮游动物生物量的变化范围为0—5112.22mg/m3, 年均值为252.40mg/m3。北部湾近岸海域浮游动物生物量同样具有明显季节变化, 平水期最高, 平均生物量为385.01mg/m3, 枯水次之, 平均值为221.41mg/m3, 丰水期最低, 平均值为150.78mg/m3。枯水期以北海市南部涠洲岛海域生物量最高, 最高生物量出现在S42站位;生物量低值区分布在钦州湾内湾茅尾海海域以及珍珠湾海域, 最低生物量出现在S4站位(图 3枯水期)。丰水期生物量平面分布以铁山港海域最高, 最高生物量出现在S39站位;钦州湾海域的生物量较低, 最低生物量出现在S21站位(图 3丰水期)。平水期生物量高值区主要集中在北海市北岸廉州湾海域、南岸近岸海域以及钦州湾外湾海域, 最高生物量出现在S29站位;钦州湾内湾茅尾海海域的生物量较低, 最低生物量出现在S15站位(图 3平水期)。

|
| 图 3 北部湾近岸海域浮游动物生物量的平面分布/(mg/m3) Fig. 3 Horizontal distribution of biomass of zooplankton in coastal Beibu Gulf |
北部湾近岸海域浮游动物优势种(包括浮游幼体)共有9种, 其中枯水期1种, 丰水期6种, 平水期5种(表 2)。枯水期以夜光虫为绝对优势种, 也是枯水期仅有的1种优势种, 其丰度占总丰度的50.80%;丰水期浮游动物优势种类最多, 优势度最高的种类为肥胖三角溞, 其次为蛇尾类长腕幼虫, 它们的丰度共占总丰度的35.2%;平水期优势度最高的种类为中型莹虾, 其次为太平洋纺锤水蚤, 它们的丰度共占总丰度的21.2%。
| 时间 Time |
优势种 Dominance species |
优势度(Y) Degree of dominance |
平均丰度/(个/m3) Average abundance |
| 枯水期Dry season | 夜光虫Noctiluca scientillans | 0.196 | 782.50 |
| 丰水期Wet season | 太平洋纺锤水蚤Acarti pacifica | 0.032 | 19.23 |
| 亚强次真哲水蚤Eucalanus subcrassus | 0.021 | 20.04 | |
| 肥胖三角溞Evadn etergestina | 0.061 | 82.87 | |
| 蛇尾类长腕幼虫Echinodermata bipinnaria larva | 0.047 | 48.01 | |
| 长尾类幼虫Mccruran larva | 0.041 | 19.92 | |
| 短尾类幼虫Zoea larva(Brachyura) | 0.024 | 18.51 | |
| 平水期Normal season | 太平洋纺锤水蚤Acarti pacifica | 0.057 | 33.49 |
| 中型莹虾Lucifer intermedius | 0.095 | 63.65 | |
| 长尾类幼虫Mccruran larva | 0.042 | 20.42 | |
| 短尾类幼虫Zoea larva(Brachyura) | 0.040 | 19.11 | |
| 蔓足类无节幼虫Nauplius larva(Cirripdia) | 0.048 | 38.37 |
2017年北部湾近岸海域浮游动物生物多样性指数的变化范围为0—4.48, 均值为2.58。北部湾近岸海域浮游动物生物多样性指数以平水期最高, 均值为3.16, 丰水期次之, 均值为2.35, 枯水期最低, 均值为2.22。其中, 枯水期生物多样性指数平面分布以铁山港以及北海南岸近岸海域为较高值区, 最高值出现在S32站位, 防城港湾海域生物多样性指数较低, 最低值出现S4站位(图 4枯水期);丰水期生物多样性指数较高值区主要分布在铁山港、廉州湾以及防城港湾海域, 最高值出现在S29站位, 钦州湾海域生物多样性指数较低, 最低值出现S21站位(图 4丰水期);平水期生物多样性指数分布相对较均匀, 最高值出现在S6站位, 最低值出现S26站位(图 4平水期)。

|
| 图 4 北部湾近岸海域浮游动物生物多样性的平面分布 Fig. 4 Horizontal distribution of diversity index of zooplankton in coastal Beibu Gulf |
采用SPSS 16.0统计分析软件对浮游动物生物量与主要环境因子进行Pearson相关性分析, 当P<0.05, 表明相关关系显著;当P < 0.01, 表明相关关系极显著, 结果如表 3所示。浮游动物生物量在枯水期和丰水期与丰度有明显的正相关, 枯水期相关性最高丰水期次之, 但在平水期生物量与丰度之间没有相关性。枯水期浮游动物生物量与pH存在极显著的正相关(P<0.01), 与悬浮物、叶绿素a及营养盐(PO43-、NO2-、NO3-、SiO32-、NH4+)存在明显的负相关(P<0.05);丰水期浮游动物生物量与pH、叶绿素a存在明显的正相关(P<0.05), 与营养盐(NO2-、NO3-、SiO32-)存在明显的负相关(P<0.05);平水期浮游动物生物量与各环境因子没有明显的相关性。
| 生物量 Zooplankton biomass |
丰度 Zooplankton abundance |
盐度 Salinity |
悬浮物 SPM |
PO43- | COD | pH | NO2- | NO3- | NH4- | SiO32- | 叶绿素a Chl-a |
| 枯水期Dry season | 0.583* | 0.191 | -0.379* | -0.323* | -0.296 | 0.635** | -0.346* | -.368* | -0.464** | -0.335* | -0.350* |
| 丰水期Wet season | 0.328* | 0.172 | -0.181 | -0.293 | 0.119 | 0.367* | -0.346* | -0.303* | 0.069 | -0.310* | 0.305* |
| 平水期Normal season | 0.043 | 0.169 | -0.093 | -0.065 | -0.082 | 0.251 | -0.141 | -0.086 | -0.128 | -0.116 | 0.195 |
| “**”表示极显著相关, “*”表示显著相关; SPM:Suspended solids | |||||||||||
北部湾近岸海域浮游动物丰度的平面分布特征在枯水期、丰水期和平水期之间均存在较大差异。枯水期浮游动物丰度的高值区主要分布在北海南部涠洲岛周边以及犀牛脚西南部附近海域, 低值区则出现在防城港以及钦州湾内湾近岸海域, 总体上略呈现为河口近岸低、外海高的分布特征(图 2枯水期)。枯水期的唯一优势种夜光虫(Noctiluca scintillans)是对该季节浮游动物丰度贡献率最大的物种, 也决定了枯水期浮游动物丰度及生物量的分布特征。夜光虫是引起我国赤潮频发的重要种类[13]。夜光虫和营养盐的分布密切相关, 其出现数量的多寡可作为指示水环境污染程度的一个依据[14-15]。有研究认为夜光虫赤潮是富营养化的结果之一[16], 水体富营养程度的不断加重是导致夜光虫赤潮日益频繁的根本原因[17]。夜光虫丰度较高的站点主要集中在远离大陆近岸的涠洲岛周边海域, 该片海域水质状况良好, 大部分符合国家第一类海水水质标准, 全年海水营养盐浓度偏低。但自1999至2008年的10年期间, 涠洲岛附近海域就发生了数次赤潮[18]。研究表明在枯水期, 粤西沿岸流经由琼州海峡的侵入在涠洲岛附近海域形成上升流[19-20], 因此枯水期涠洲岛海域夜光虫等赤潮物种大量生长可能与该海区独特的营养盐补充机制有关。在涠洲岛发生的赤潮中, 2008年以前主要以束毛藻等具有较低营养盐适应机制的种类, 而在2008年开始出现夜光虫赤潮, 这可能与涠洲岛周边海域营养盐增加有关。近年来随着涠洲岛上的旅游业不断发展, 游客增多带来的入海排污量增加会直接影响周边海域水体环境的变化, 水体营养盐水平上升可能会增加夜光虫赤潮爆发的风险。
丰水期浮游动物丰度和生物量的分布特征与枯水期相似(图 2丰水期和图 3丰水期), 主要呈现出由近岸向外海增高趋势。丰水期浮游动物优势种较多, 以肥胖三角溞的优势度最大。该物种是亚热带近海种, 具有广温广盐的生态特征, 最适温度高于25℃, 其高丰度分布区是沿岸水团的标志[21]。丰水期由于水温较高有利于肥胖三角溞形成最优势种类, 而丰水期径流大量输入使沿岸水团往外推移可能是导致肥胖三角溞在相对靠外站点数量较高从而决定了浮游动物丰度的分布特征。北部湾位于亚热带, 平水期10月份的水温仍较高, 因此在本次调查中发现平水期浮游动物的优势种仍是以适宜水温较高的近岸暖水性类群为主, 其中的太平洋纺锤水蚤和中型莹虾都是典型的亚热带沿岸种。相对于丰水期, 平水期径流减弱, 沿岸水回退到河口近岸海区, 因此浮游动物丰度的分布较为均匀, 即使是廉州湾及铁山港内浮游动物的丰度也较高(图 2平水期)。平水期浮游动物种类最丰富, 而且丰度分布相对均匀, 没有呈现出与枯水期及丰水期较为一致的明显分布特征, 甚至略显从近岸到离岸丰度降低的相反分布特征, 表明了浮游动物的种类组成结构以及优势种的演替对浮游动物的分布特征有着重要的决定作用, 也是导致分布特征季节变化的主要原因。
北部湾近岸海域浮游动物的生物量在枯水期总体呈现出从河口近岸到离岸海区逐渐增高的趋势特征, 但在丰水期呈现出从北仑河口至英罗港一线较高而河口及外海较高, 平水期呈现出自钦州港至北海南部生物量较高而其他河口及外海较低的特征, 表明各季节之间的生物量分布特征也具有明显的季节变化。在分析中发现浮游动物的丰度分布特征与优势种类数量分布有着密切关系, 但生物量的分布主要是受一些个体较大种类的影响, 如丰水期浮游动物生物量的空间分布主要由丰度很低(6.45个/m3)的球型侧腕水母决定。与肥胖三角溞相比, 含水量大的水母类即使数量很少, 其湿重生物量也远比个体小的枝角类重, 因此出现了丰度与生物量不完全一致的现象。类似的情况在平水期更为显著, 平水期浮游动物生物量的空间分布与丰度的分布并不一致, 浮游动物生物量的主要贡献者仍是含水量较大的水母类, 浮游动物生物量与丰度之间没有相关性(表 3), 表明了优势种类主要决定了浮游动物丰度的分布特征及变化, 而含水量较大或个体较大的种类主要决定着浮游动物生物量的分布特征及变化, 也由此导致了丰度和生物量分布特征的不一致性。
3.2 影响浮游动物分布及季节变化的关键环境因子北部湾近岸海域枯水期和丰水期浮游动物丰度、生物量的分布特征主要呈现为自河口近岸海域往外海递增的趋势特征, 浮游动物的生物量虽然没有与盐度呈现出显著正相关性, 但和悬浮物及营养盐等环境因子呈显著负相关(表 3)。北部湾近岸海域河口海湾众多, 水文环境复杂, 同时受到北部湾沿岸水、混合水及外海水的影响[22]。各项物理化学指标由近岸向外海都存在浓度梯度, 表明沿岸径流的污染物输入对近岸海域水质变化起着主导作用。其中影响近岸海域水质的污染物主要是无机氮和无机磷[23], 氮、磷的持续输入增加了近岸海域水体富营养化的风险。近几年的监测数据显示局部海域已经出现富营养化状况[24-25], 海水富营养化已然成为北部湾近岸海域突出的环境问题。北部湾近岸海域枯水期尽管径流量减少, 但营养盐浓度较高, 而丰水期径流输入的营养盐大量增加, 在河口海湾局部海域形成营养盐浓度较高的海域。营养盐的持续输入改变水体中营养盐的组成和比率, 从而影响浮游植物群落组成和数量, 由此导致浮游动物丰度和生物量的下降[26]。此外, 枯水期地表径流量减少, 沿岸水势力减弱, 外海水势力向内推进作用增强, 在河口海湾形成水体浑浊的混合区。丰水期径流增加不仅使河口近岸悬浮泥沙大量增加, 而且会导致水体悬浊的混合区范围扩大。水体透明度下降不利于浮游植物进行光合作用[11, 27], 进而影响浮游动物的生长繁殖。同步观测的结果表明北部湾近岸海域无论是枯水期还是丰水期营养盐、悬浮物空间分布都呈现从河口向外湾递减的趋势, 这与浮游动物生物量的分布特征相反, 相关性分析也表明二者呈显著负相关, 由此表明径流导致的悬浮物、营养盐等的变化可能是决定北部湾近岸海域浮游动物丰度和生物量空间分布的主要因素。
除了悬浮物和营养盐, 水体的pH值以及叶绿素a对浮游动物也具有较大的影响。枯水期浮游动物生物量受pH值变化的影响较大, 两者呈极显著的正相关(P<0.01)。丰水期浮游动物生物量也与pH值有显著正相关性(P<0.05)。这反映出水域的水质环境变化能直接或间接影响浮游动物密度的变化, 一定程度上成为浮游动物生长繁殖的限制因子[28]。枯水期浮游动物的优势种为夜光虫, 大量的夜光虫捕食浮游植物会直接导致水体中叶绿素的骤减[29]。枯水期浮游动物的生物量与叶绿素a呈显著的负相关关系(P<0.05), 暗示了夜光虫大量生长给浮游植物带来了较大摄食压力, 可能抑制了其他浮游动物的生长, 因而夜光虫成为了枯水期的绝对优势种, 也决定了枯水期浮游动物丰度和生物量的分布特征。而丰水期浮游动物生物量与叶绿素a值有显著正相关性(P<0.05), 这可能跟丰水期浮游动物优势种种类较多有关, 各物种之间相互制衡没有形成绝对优势种, 捕食对象的多样化也减轻了对浮游植物的摄食压力。浮游动物丰度与叶绿素a的正相关关系也可能与浮游动物的选择性摄食有关, 大多数以浮游植物为食的浮游动物与叶绿素a具有正相关关系[30], 即叶绿素a通过影响浮游动物的食物而影响其群落结构。
本研究发现北部湾近岸海域浮游动物的种类数为平水期远高于枯水期、丰水期, 丰度呈现出枯水期>平水期>丰水期的季节变化特征, 生物量呈现出平水期>枯水期>丰水期的季节变化特征, 由此导致了生物多样性呈现出平水期远高于枯水期、丰水期的特征。研究表明稳定的环境有利于“机会种类”形成绝对优势却不利于形成较高生物多样性的条件, 剧烈变动的环境也不利于形成较高的生物多样性, 而中度稳定的环境最利于保持较高的生物多样性。在三沙湾、福宁湾、椒江口以及珠江口[31-34]等海域的研究中同样发现在环境相对稳定的枯水期, 浮游动物丰度较高但生物多样性指数最低, 而在环境处于较稳定的平水期往往形成各优势种丰度较小, 生物多样性指数最高的结构格局。北部湾近岸海域入海河流众多, 枯水期径流输入最小, 沿岸水势力减弱, 外海水势力向内推进, 整个近岸海域基本上被相对高盐的海水所占据, 环境相对最稳定, 同时营养盐浓度也不低, 透明度增加, 因此非常有利于一些机会种类包括浮游植物和浮游动物形成绝对优势种类, 如北部湾近岸海域发生的赤潮主要爆发在春、冬季节即枯水期[18], 导致了夜光虫等机会种类爆发, 其他在稳定环境中不具备竞争力的种类减少, 因而枯水期浮游动物的丰度最高, 但种类数量及生物多样性却相对较低。丰水期沿岸江河入海径流很强, 沿岸水向外扩张, 同时也是外海水势力最强时期[22], 两股强势的水系混合导致水流湍急, 浮游生物在受激烈的径流和水质变动等影响较大的河口海湾生长必须要付出更多的能量[7]。复杂多变的水体环境不利于浮游生物生长。此外, 混合水面积的扩大将河流污染物的影响范围扩大, 从而导致丰水期的浮游动物生物量和丰度均较低, 种类数量及生物多样性也比较低。相对而言, 平水期北部湾近岸海域的环境处于中度扰动状态, 虽然江河径流量已减弱但仍较强, 环境的扰动度没有枯水期那么轻也没有丰水期如此剧烈, 此时水体变清, 营养丰富, 浮游植物生长迅速, 但机会种类也难以爆发, 食物及其他小型种类的增加也促进了较大浮游动物的生长, 因此浮游动物种类、生物量和生物多样性指数相较于丰水期均有明显的上升, 是三个水期中生物多样性最丰富的季节, 浮游动物群落结构趋于稳定。由此表明径流强度以及近岸海域水团环境的稳定性可能是决定浮游动物多样性季节变化的主要因素。
3.3 北部湾浮游动物群落结构特征及与其他海区比较北部湾近岸海域浮游动物群落中以近岸暖水性种类占大多数, 根据调查海域的环境条件和浮游动物群落结构特征, 北部湾近岸海域的浮游动物属于亚热带沿岸群落。此外, 调查海区浮游动物群落一个显著特点是阶段性浮游动物占有重要的位置。优势种中出现多类浮游幼体, 如长尾类幼体、短尾类幼体、蔓足类幼体和蛇尾类幼虫。长尾类幼体和短尾类幼体是丰水期和平水期共有的优势类群, 丰水期、平水期浮游幼体的优势类较多, 优势度较高, 这反映了北部湾近岸海域在较低水温的春、冬季大多数动物不繁殖而夏、秋季随着水温升高各类动物大量繁殖这一规律。这与北部湾相对外海海域的浮游动物群落结构特征具有明显的区别:外海海域浮游动物种类更为丰富, 且终年以质体较重的中型浮游动物(肥胖箭虫、亚强次真哲水蚤和中型莹虾)占显著优势, 因此优势种数量的消长决定了外海域浮游动物的丰度和生物量的分布变化[4]。
本次调查共鉴定出浮游动物275种(类), 比东南沿海几个典型河口海湾[33-38]的种类数都要多, 全年平均生物多样性指数为2.58, 也高于其他几个海湾, 说明北部湾近岸海域浮游动物种类组成复杂丰富(表 4)。北部湾近岸海域浮游动物丰度和生物量均比纬度相近的珠江口和湛江港高;北部湾近岸海域浮游动物丰度与高纬度的椒江口和长江口丰度相比差异不大, 但生物量明显较低;其浮游动物丰度远高于瓯江口, 但浮游动物生物量反而低于瓯江口, 这主要是因为长江口、椒江口和瓯江口海域的优势种以大型浮游动物(水母类、毛颚类、磷虾类、糠虾类、体长>2.0mm桡足类)为主, 而北部湾近岸海域浮游动物优势种中除亚强次真哲水蚤和中型莹虾为中型浮游动物外, 其他大部分优势种如夜光虫、长尾类幼虫、短尾类幼虫、蔓足类无节幼虫和棘皮动物长腕幼虫均为小型浮游动物。小型浮游动物对生物量的贡献不大, 这也是北部湾近岸海域浮游动物生物量相对偏低的原因。从本次调查结果来看, 北部湾近岸海域浮游动物群落结构整体有小型化的趋势。事实上, 笔者在对2014—2015年钦州湾海域浮游动物群落结构进行研究时已经发现趋于小型化[8], 这很可能是整个北部湾近岸海域都存在的问题。已有研究表明, 浮游动物群落结构小型化可能与水体富营养化有关[39]。水体富营养化的加重和营养盐结构的变化使得近岸海域浮游植物群落构成趋于小型化[40-41]并通过食物链的传递效应, 进一步引起主要摄食者浮游动物的小型化[42]。因此, 北部湾近岸海域水体富营养化加重可能是引起浮游动物群落结构也趋于小型化的一个重要因素。浮游动物小型化也会驱使捕食浮游动物的鱼类的小型化, 而北部湾海域渔业资源结构也确实已趋于小型化[43]。随着时间的推移和海域环境的变化, 北部湾近岸海域浮游动物群落结构小型化可能会逐渐加重, 因此这一现象需引起广泛关注。
| 海域 Area |
调查时间 Time |
总个体数/ (个/m3) Total number |
总生物量/ (mg/m3) Total biomass |
总种类数 Species amount |
多样性指数 Diversity index |
文献来源 Literature sources |
| 椒江口 Jiaojiang Estuary |
2009 | 887.74 | 531.48 | 106 | 2.20 | [34] |
| 珠江口 Pearl River Estuary |
2015 | 298.30 | 74.81 | 73 | 2.24 | [35] |
| 长江口 Changjiang River Estuary |
2009—2010 | 624.77 | 718.63 | 178 | 2.12 | [36] |
| 湛江港 Zhanjiang Bay |
2010 | 498.73 | 190.20 | 51 | 2.10 | [37] |
| 瓯江口 Oujiang Estuary |
2007 | 172.65 | 365.58 | 94 | 2.56 | [38-39] |
| 北部湾近岸海域 Beibu Gulf |
2017 | 789.95 | 244.81 | 275 | 2.58 | 本调查 |
| [1] |
徐姗楠, 林华剑, 戴明, 肖雅元, 陈作志. 广西近岸海域浮游植物群落的生态特征. 生态学杂志, 2014, 33(10): 2733-2739. |
| [2] |
陈兰, 蒋清华, 石相阳, 蓝文陆. 北部湾近岸海域环境质量状况、环境问题分析以及环境保护建议. 海洋开发与管理, 2016, 33(6): 28-32. DOI:10.3969/j.issn.1005-9857.2016.06.006 |
| [3] |
张才学, 周凯, 孙省利, 陈春亮, 张际标, 张瑜斌. 深圳湾浮游动物的群落结构及季节变化. 生态环境学报, 2010, 19(11): 2686-2692. DOI:10.3969/j.issn.1674-5906.2010.11.029 |
| [4] |
郑白雯, 曹文清, 林元烧, 郑连明, 张文静, 杨位迪, 王宇杰. 北部湾北部生态系统结构与功能研究Ⅱ.浮游动物数量分布及优势种. 海洋学报, 2014, 36(4): 82-90. DOI:10.3969/j.issn.0253-4193.2014.04.004 |
| [5] |
郑挺, 林元烧, 曹文清, 张文静, 郑连明, 王宇杰, 杨位迪. 北部湾北部生态系统结构与功能——浮游动物空间生态位及其分化. 生态学报, 2014, 34(13): 3635-3649. |
| [6] |
阙江龙, 徐兆礼, 孙鲁峰. 北部湾西北部饵料浮游动物季节变化及其与鱼卵、仔稚鱼的关系. 中国水产科学, 2015, 22(5): 1027-1035. |
| [7] |
蓝文陆, 李天深, 刘勐伶, 庞碧剑, 刘昕明. 钦州湾丰水期和枯水期浮游动物群落特征. 海洋学报, 2015, 37(4): 124-132. DOI:10.3969/j.issn.0253-4193.2015.04.012 |
| [8] |
庞碧剑, 李天深, 蓝文陆, 黎明民, 骆鑫, 陈莹. 钦州湾秋季和春季浮游动物分布特征及影响因素. 生态学报, 2018, 38(17): 6204-6216. |
| [9] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB 17378.7-2007海洋监测规范第7部分: 近海污染生态调查和生物监测.北京: 中国标准出版社, 2008.
|
| [10] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB 17378.7-2007海洋监测规范第4部分: 海水分析.北京: 中国标准出版社, 2008.
|
| [11] |
徐兆礼, 王云龙, 陈亚瞿, 沈焕庭. 长江口最大浑浊带区浮游动物的生态研究. 中国水产科学, 1995, 2(1): 39-48. DOI:10.3321/j.issn:1005-8737.1995.01.002 |
| [12] |
Shannon C E, Weaver W. The Mathematical Theory of Communication. Urbana, USA: University of Illinois Press, 1949.
|
| [13] |
王丽, 王保栋, 陈求稳, 汤新武, 韩瑞. 三峡三期蓄水后长江口海域浮游动物群落特征及影响因子. 生态学报, 2016, 36(9): 2505-2512. |
| [14] |
徐家铸, 苏翠荣. 海州湾浮游动物的种类组成和分布(Ⅰ)——夜光虫和砂壳纤毛虫. 南京师大学报:自然科学版, 1995, 18(4): 125-129. |
| [15] |
张天文, 朱丽岩, 徐培培, 周浩, 戚本金. 胶州湾夜光虫丰度的季节变化和分布特征. 中国海洋大学学报, 2009, 39(S1): 89-93. |
| [16] |
徐兆礼. 长江口夜光藻(Noctiluca scintillans)年间变化和水域富营养化趋势. 海洋与湖沼, 2009, 40(6): 793-798. DOI:10.3321/j.issn:0029-814X.2009.06.019 |
| [17] |
黄长江, 齐雨藻, 黄奕华, 林小涛. 南海大鹏湾夜光藻种群生态及其赤潮成因分析. 海洋与湖沼, 1997, 28(3): 245-255. DOI:10.3321/j.issn:0029-814X.1997.03.004 |
| [18] |
罗金福, 李天深, 蓝文陆. 北部湾海域赤潮演变趋势及防控思路. 环境保护, 2016, 44(20): 40-42. |
| [19] |
侍茂崇. 北部湾环流研究述评. 广西科学, 2014, 21(4): 313-324. DOI:10.3969/j.issn.1005-9164.2014.04.001 |
| [20] |
谭光华. 北部湾海区水文结构及其特征的初步分析. 海洋湖沼通报, 1987(4): 7-15. |
| [21] |
徐兆礼, 高倩, 陈华, 陈佳杰, 蔡萌. 东海浮游枝角类和涟虫类生态适应性. 生态学杂志, 2007, 26(11): 1782-1787. |
| [22] |
陈波. 北部湾水系形成及其性质的初步探讨. 广西科学院学报, 1986, 2(2): 92-95. |
| [23] |
李斌, 谭趣孜, 李蕾鲜, 巫冷蝉, 刘成辉, 戴培建, 范航清. 2014年北部湾主要河流污染状况及污染物入海通量. 广西科学, 2018, 25(2): 172-180. |
| [24] |
蓝文陆. 钦州湾枯水期富营养化评价及其近5年变化趋势. 中国环境监测, 2012, 28(5): 40-44. DOI:10.3969/j.issn.1002-6002.2012.05.009 |
| [25] |
杨斌, 鲁栋梁, 钟秋平, 张志清, 李尚平. 钦州湾近岸海域水质状况及富营养化分析. 中国环境监测, 2014, 30(3): 60-64. DOI:10.3969/j.issn.1002-6002.2014.03.013 |
| [26] |
徐晓群, 曾江宁, 陈全震, 刘晶晶, 杜萍, 王桂忠. 乐清湾海域浮游动物群落分布的季节变化特征及其环境影响因子. 海洋学研究, 2012, 30(1): 34-40. DOI:10.3969/j.issn.1001-909X.2012.01.005 |
| [27] |
徐兆礼, 王云龙, 白雪梅, 陈亚瞿. 长江口浮游动物生态研究. 中国水产科学, 1999, 6(5): 51-58. |
| [28] |
张光星, 吴钟解, 陈石泉, 蔡泽富, 王道儒. 春季三亚湾浮游动物生态特征与环境因子的关系. 福建水产, 2014, 36(3): 176-184. DOI:10.3969/j.issn.1006-5601.2014.03.002 |
| [29] |
孙东, 赵冬至, 文世勇, 邢小罡, 王林. 夜光藻赤潮与环境因子关系的模糊分析. 海洋环境科学, 2010, 29(1): 70-75. DOI:10.3969/j.issn.1007-6336.2010.01.015 |
| [30] |
徐晓群, 曾江宁, 陈全震, 刘晶晶, 杜萍, 王桂忠. 浙江三门湾浮游动物优势种空间生态位. 应用生态学报, 2013, 24(3): 818-824. |
| [31] |
徐佳奕, 徐兆礼. 三沙湾浮游动物生态类群演替特征. 生态学报, 2013, 33(5): 1413-1424. |
| [32] |
王兴春. 福建福宁湾尖刀蛏保护区中小型浮游动物群落结构的季节变化. 海洋环境科学, 2014, 33(6): 870-875. |
| [33] |
杜萍, 徐晓群, 刘晶晶, 曾江宁, 陈全震, 寿鹿, 廖一波, 周青松. 椒江口春、秋季浮游动物分布特征及与主要环境因子的关系. 应用与环境生物学报, 2011, 17(4): 486-494. |
| [34] |
徐姗楠, 龚玉艳, 詹凤娉, 许友伟, 陈作志. 珠江口海域南沙段浮游动物群落生态特征. 海洋湖沼通报, 2017(6): 82-90. |
| [35] |
殷晓龙, 徐兆礼. 长江口不同水域浮游动物群落结构比较. 应用与环境生物学报, 2015, 21(1): 88-95. |
| [36] |
姚少慧, 于红兵, 王霞, 孙宗勋. 湛江港夏秋季浮游动物群落结构的分布与变化. 广东农业科学, 2012, 39(13): 192-196. DOI:10.3969/j.issn.1004-874X.2012.13.064 |
| [37] |
高倩, 徐兆礼. 瓯江口夏、秋季浮游动物种类组成及其多样性. 生态学杂志, 2009, 28(10): 2048-2055. |
| [38] |
高倩, 徐兆礼. 瓯江口水域夏、秋季浮游动物数量时空分布特征. 中国水产科学, 2009, 16(3): 372-380. DOI:10.3321/j.issn:1005-8737.2009.03.009 |
| [39] |
张才学, 龚玉艳, 王学锋, 孙省利. 湛江港湾浮游桡足类群落结构的季节变化和影响因素. 生态学报, 2011, 31(23): 7086-7096. |
| [40] |
Pan Y L, Rao D V S. Impacts of domestic sewage effluent on Phytoplankton from Bedford Basin, eastern Canada. Marine Pollution Bulletin, 1997, 34(12): 1001-1005. DOI:10.1016/S0025-326X(97)00115-X |
| [41] |
Dippner J W. Competition between different groups of phytoplankton for nutrients in the Southern North Sea. Journal of Marine Systems, 1998, 14(1/2): 181-198. |
| [42] |
龚玉艳, 张才学, 陈作志, 叶丽娣, 陈伟杰, 王伟荣, 孙省利. 湛江湾浮游动物群落结构特征及其周年变化. 海洋科学, 2015, 39(12): 46-55. DOI:10.11759/hykx20141027001 |
| [43] |
王雪辉, 邱永松, 杜飞雁, 林昭进, 孙典荣, 黄硕琳. 北部湾秋季底层鱼类多样性和优势种数量的变动趋势. 生态学报, 2012, 32(2): 333-342. |