 2019, Vol. 39
2019, Vol. 39文章信息
- 潘玉梅, 唐赛春, 韦春强, 李象钦, 吕仕洪, 王云波
- PAN Yumei, TANG Saichun, WEI Chunqiang, LI Xiangqin, Lü Shihong, WANG Yunbo
- 两种本地植物种子萌发对飞机草的化感耐受性及其相互竞争
- Tolerance of seed germination in two native species to the allelopathic effect of Chromolaena odorata and the competition between them
- 生态学报. 2019, 39(19): 7302-7310
- Acta Ecologica Sinica. 2019, 39(19): 7302-7310
- http://dx.doi.org/10.5846/stxb201806221374
-
文章历史
- 收稿日期: 2018-06-22
- 网络出版日期: 2019-08-16





2. 广西国有六万林场, 玉林 537000
2. Guangxi State-owned Liuwan Forest Farm, Yulin 537000, China
生物入侵常使原有的生态平衡系统遭到破坏, 造成环境公害爆发, 给入侵地的农业、林业、畜牧业及人类健康带来严重损失, 如何控制外来生物的继续入侵, 消除或降低生物入侵造成的危害, 是入侵生态学领域一直关注的热点问题。尽管目前已对外来入侵植物开展了较多控制方法的研究, 如人工或机械清除、化学防治、天敌生物防治等, 但这些方法在应用上或效果维持时间短暂, 或污染生态环境, 或存在二次入侵的风险, 具有一定的局限性, 因此, 根据不同入侵生物的入侵特性及危害程度, 寻找相应的安全有效且持续的控制方法是目前有效管理外来入侵生物亟待解决的问题。
替代控制是根据植物种间竞争关系, 利用有生态效益或经济价值的植物取代入侵植物的一种生态控制方法, 具有持效久、生态和经济效果良好、利于生态修复等特性, 是目前比较理想的外来入侵植物控制方法之一, 也是防控入侵植物的新方向[1-2], 但替代目标植物的筛选是其关键。近年来, 一些学者针对我国一些重要入侵植物已筛选了部分替代植物, 如利用柱花草(Stylosanthes humilis)、东非狼尾草(Pennisetum clandestinum)和勃氏甜龙竹(Dendrocalamus brandisi)混合种植或利用红薯(Ipomoea batatas)对入侵植物薇甘菊(Mikania micrantha)的控制[3-4];利用紫穗槐(Amarpha fruticosa)、草地早熟禾(Poa pratensis)及菊芋(Helianthus tuberosus)等对公路边豚草(Ambrosia artemisiifolia)的替代控制[5];利用白蝶合果芋(Syngonium podophllum)和细叶萼距花(Cuphea hyssopifolia)对入侵植物猫爪藤(Macfadyena unguis-cati)的替代控制[6];利用宽叶雀稗(Paspalum wetsfeteini)对紫茎泽兰(Eupatorium adenophorum)的替代控制等[7]。但目前筛选的目标植物多为牧草或经济作物, 对生境要求较高, 有些还是外来物种, 有可能产生新的生物入侵。外来物种入侵的生境往往地形复杂, 很多地区土壤贫瘠、水分不足, 经济作物或牧草很难适应。因此, 需要开展更多的适应不同入侵生境的本地替代植物的筛选及替代控制模式的相关研究。
飞机草[Chromolaena odorata (L.) R. King et H.Rob.]是菊科(Compositae)香泽兰属(Chromolaena)的多年生草本或亚灌木, 原产于中、南美洲, 现已广泛分布于亚洲、非洲、大洋洲和西太平洋群岛的大部分热带及亚热带地区, 是世界上危害最严重的外来入侵物种之一[8-9]。该植物自从1934年在我国云南被首次发现以后, 在南方地区迅速扩散, 现已广泛分布于台湾、云南、海南、贵州、四川、广东、广西等多个省区[10], 是国家环保总局与中科院联合发布的第一批入侵最严重的外来入侵物种之一[11]。近年来, 许多学者对其生物学特性[8, 11]、化感作用[12-13]、生理生态适应性[14-15]及控制方法[16-17]等多方面展开了研究, 对其生物学特性及其入侵机制等方面有了初步了解, 但由于该植物繁殖能力强、种子数量多、易密集成丛排挤其他植物及其入侵生境复杂多样, 目前对其管理和控制方面仍未找到切实可行的有效途径或可持续长久的控制方法。
化感作用是多数外来入侵植物共有的特性, 也被认为是飞机草成功入侵的主要机制之一[18-19]。研究表明入侵植物的化感物质可以通过雨水淋溶、根部渗出、植株腐烂分解等方式释放到周围环境抑制其他植物生长, 替代目标植物是否能够耐受这些化感物质从而成功萌发和再生是替代植物种群能够长期维持的关键。卢向阳等[7]和高尚宾等[20]研究认为替代控制入侵植物的目标物种选择的关键之一就是可以耐受入侵植物的化感作用。另外, 替代目标植物是否对入侵种具有竞争抗性也是其能否替代成功的关键因素之一。
在广西对飞机草的野外调查中发现, 在飞机草入侵的岩溶山地、弃荒地、山脚路边等区域常有本地植物假地豆(Desmodium heterocarpon)和白饭树(Flueggea virosa)伴生, 因此设想这两种本地植物可能对飞机草的入侵具有一定的抗性, 可以作为其替代控制的目标植物。假地豆是蝶形花科山蚂蟥属亚灌木植物, 茎直立或平卧斜生, 茎长约90cm, 分枝多, 叶茂盛, 果多而密, 生长快。广泛分布于我国南部各个地区, 常生于山坡、灌丛或林中。全株药用, 可清热解毒、治跌打骨折、蛇伤;韧皮纤维可造纸和制绳索, 具较高经济和药用价值。白饭树是大戟科白饭树属多年生灌木植物, 植株高约2m, 分枝多, 叶茂密, 果实较多, 常生于丘陵地及石灰岩石山中下部地区, 在我国西南和东南广大地区均有分布。它是我国传统中药, 具有清热解毒、消肿止痛、止痒止血等功效, 还可以外用治疗水痘、皮炎、湿疹等多种皮肤病, 也有用于治疗多种关节炎疾病的报道[21]。
因此, 基于是否对入侵植物具有化感耐受性和竞争力两个因素, 我们以本地灌木假地豆和白饭树为研究对象, 开展了以下实验:1)两种本地植物的种子萌发对不同浓度飞机草根、茎、叶提取液的化感耐受性实验;2)飞机草与两个本地植物的生长竞争关系。拟通过以上实验探讨两种本地植物替代控制飞机草的潜力, 为飞机草的生态控制及其入侵地的生态修复提供科学依据。
1 材料与方法 1.1 实验材料实验中所需的飞机草植株和种子均采自广西百色市平果县飞机草入侵地(23°22′N, 107°22′E, 240 m a.s.l.)。飞机草植株采自飞机草入侵多年的山坡弃荒地和山脚路边等不同生境, 采集处于生长旺盛期的飞机草全株, 采集时每个植株间隔一定距离。同样, 从这些不同的生境采集飞机草成熟饱满的种子, 采集时每个植株间隔至少20m;本地植物假地豆和白饭树的种子均采自百色市平果县飞机草入侵地周边山坡和路边, 采集时每个植株间隔至少20m。所采集植物种子分别装入信封带回实验室保存备用。
1.2 实验方法 1.2.1 本地种种子萌发对飞机草水提液的耐受性实验(1) 飞机草水提液的制备
将采集的飞机草植株根、茎、叶分离后, 分别装袋带回实验室, 为方便存储备用, 用自来水将其灰尘和泥土洗净、荫干后, 用植物粉碎机分别将它们粉碎成粉末状保存备用。根据郑丽等[22]的方法, 按每1000mL蒸馏水25g干物质的比例浸泡, 室温下浸提48h后, 用双层纱布过滤2次, 得到浓度为2.5%的飞机草根、茎、叶水浸提液母液, 然后将一部分母液分别用蒸馏水稀释成浓度为1.25%、0.25%的根、茎、叶提取液, 放置于4℃冰箱内保存备用。
(2) 本地种种子萌发实验
采用培养皿滤纸法进行实验。成熟饱满的本地植物假地豆和白饭树的种子分别用75%的酒精消毒后, 将其分别均匀点播在铺有两层滤纸的直径为11cm的培养皿中, 每皿均匀放置本地植物种子100粒。飞机草根、茎、叶母提取液及其2个不同浓度稀释液处理分别设置4个重复, 点播种子前每个培养皿先用相应的提取液将滤纸润湿, 并以蒸馏水为对照, 置于昼夜温度为25℃/20℃, 光照周期12h/12h的光照培养箱内进行种子萌发。每个处理每隔2天喷洒一次相应浓度的溶液以保持滤纸湿润, 同时记录种子发芽数, 以胚根冲破种皮为发芽标准, 直至连续3天不再有种子萌发时实验结束, 实验总萌发时间持续30天, 统计各处理下本地种的种子萌发率, 计算其与对照相比的相对萌发率。
种子萌发率(G)=供试种子发芽数/供试种子总数×100%;
相对萌发率(RG)=TG/CG×100%, 其中, TG为处理的种子萌发率, CG为对照的种子萌发率。
1.2.2 飞机草与两个本地植物的竞争实验飞机草与两个本地种的竞争实验在广西植物研究所实验基地露天空旷处采用盆栽法进行(位于桂林市雁山区雁山镇, 25°04′49.6″ N, 110°18′01.8″ E, 170m a.s.l.)。该地属中亚热带季风气候, 年平均气温17.8℃, 最冷月1月平均温度5.8℃, 最热月7月平均温度28℃;年平均降水量1949.5 mm。2017年4月中旬, 在温室大棚内将飞机草、假地豆和白饭树成熟饱满的种子分别种在装有相同土质的塑料盆内进行育苗, 待植物幼苗长至2—4叶期, 选择株高、基茎基本一致的幼苗移植到装有相同壤土的塑料盆内(内径24cm, 底径16cm, 高18cm), 每盆装土约4kg。入侵植物与两个本地植物分别设置单种和混种处理, 单种为每种植物每盆种植2株植物, 混种处理也为每盆种植2株植物, 即入侵植物与本地植物各1株, 每种处理设置7个重复。实验期间每天浇足量的水, 保持盆内土壤湿润, 及时清除杂草和防治病虫害, 并每周随机移动花盆1次以避免位置效应。植物生长4个月左右进行收获, 记录每种植物的株高, 将地上茎叶和地下根分开并分别在60℃下烘干至恒重用电子天平称重(精确度为0.001), 计算其总生物量、地上生物量比(地上生物量/总生物量)、地下生物量比(地下生物量/总生物量)和根冠比(地下生物量/地上生物量)。采用相对产量(RY)和竞争攻击力系数(A)[23]衡量物种间的竞争力。相对产量(RY)和竞争攻击力系数(A)根据以下公式计算:
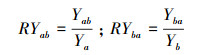
|
(1) |
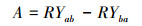
|
(2) |
式中:Yab为飞机草与本地种混种时单株生物量, Ya为飞机草单种时单株生物量;Yba为本地种与飞机草混种时单株生物量, Yb为本地种单种时单株生物量。
RYab < 1时, 飞机草对本地种的竞争力小于对飞机草本身的竞争力, 飞机草受本地物种的竞争影响比其本身大, 即种间竞争大于种内竞争;RYab=1时, 2个种的竞争力水平相当, 即种间竞争等于种内竞争;RYab>1时, 飞机草对本地物种的竞争力大于对飞机草本身的竞争力, 飞机草受本地植物的竞争影响比其本身小, 即种内竞争大于种间竞争。
A < 0表明飞机草比本地植物竞争力弱;A=0表明飞机草与本地种竞争力相当;A>0表明飞机草比本地植物竞争力强。本地植物对飞机草的竞争攻击力系数与飞机草对本地植物的竞争攻击力系数相反, 在此不再作详细说明。
1.3 数据处理所有数据分析均在SPSS 17.0软件中进行, 利用One-way ANOVA方差分析本地植物种子相对萌发率在飞机草各部位不同浓度提取液处理间的差异性及入侵植物与本地植物竞争时生长参数与对照的差异性;利用单样本T测验分析相对产量(RY)与1及攻击力系数A与0之间的差异显著性, 显著水平均为0.05。利用EXCEL 2007软件进行作图。
2 研究结果 2.1 本地种在飞机草的不同部位提取液处理下的种子萌发率研究结果表明, 本地植物假地豆种子萌发率在飞机草根部各浓度浸提液处理下均与对照无显著差异, 且在各浓度处理间也无显著差异(P>0.05);在飞机草根部各浓度提取液处理下, 假地豆的相对萌发率由低浓度到高浓度分别为67.26%、82.14%和103.57%。假地豆种子相对萌发率在飞机草茎部各浓度提取液处理下与对照相比也无显著差异(P>0.05), 在1.25%茎浓度提取液处理下相对萌发率为114.29%, 显著高于0.25%(82.14%)和2.5%浓度(78.57%)处理(P < 0.05)。与对照相比, 假地豆种子萌发率在飞机草2.5%叶提取液处理下显著受到抑制(P < 0.05), 相对萌发率为50%;在0.25%和1.25%叶提取液处理下未受到显著影响(P>0.05), 相对萌发率分别为117.86%和64.29%。
与对照相比, 本地植物白饭树种子相对萌发率在飞机草0.25%根提取液处理下受到显著促进作用(P < 0.05), 相对萌发率为107.02%;在1.25%和2.5%根提取液处理下未受到显著影响(P>0.05), 其相对萌发率分别为101.4%和102.25%。白饭树种子萌发率在飞机草茎和叶各浓度提取液处理下均未受到显著影响(P>0.05), 在各浓度茎提取液处理下, 其相对萌发率由低浓度到高浓度分别为104.49%、105.06%和101.12%;在各浓度叶提取液处理下, 其相对萌发率由低浓度到高浓度分别为97.75%、99.72%和93.54%。

|
| 图 1 飞机草提取液对假地豆和白饭树的种子相对萌发率的影响(平均值±标准误) Fig. 1 Effects of Chromolaena odorata water leachate on the relative seed germination of Desmodium heterocarpon and Flueggea virosa (means±SE) 不同小写字母表示相同部位不同浓度提取液处理间的差异显著, P < 0.05 |
飞机草与假地豆混种时, 与单种相比, 飞机草的株高、地下生物量比和根冠比均显著降低(P < 0.05), 地上生物量比显著增加(P < 0.05), 而总生物量无显著变化(P>0.05)(图 2);本地植物假地豆的株高、地上生物量比、地下生物量比和根冠比与单种相比均无显著变化, 而总生物量却显著高于单种(P < 0.05)(图 3)。飞机草与白饭树混种时, 与单种相比, 飞机草的株高、总生物量、地上生物量比和地下生物量比均显著增加(P < 0.05), 根冠比显著降低(P < 0.05)(图 2);本地植物白饭树的株高、总生物量、地上生物量比均显著低于单种, 而地下生物量比和根冠比显著高于单种(P < 0.05)(图 3)。同时, 除了地上生物量比外, 飞机草的其他参数在与假地豆竞争处理下均显著低于与白饭树竞争处理的(P < 0.05)(图 2)。

|
| 图 2 飞机草单种与混种时的生长变量(平均值±标准误) Fig. 2 The growth variables of Chromolaena odorata when planted separately and grown with the two native species (means±SE) “F+J”表示飞机草与假地豆混种, “F+B”表示飞机草与白饭树混种; 不同小写字母表示飞机草的生长变量在不同处理间的差异性, P < 0.05 |

|
| 图 3 本地物种单种与混种时的生长变量(平均值±标准误) Fig. 3 The growth variables of the native species when planted separately and grown with C. odorata (means±SE) 不同字母表示本地物种的生长变量在混种处理时与对照间的差异性, P < 0.05 |
飞机草与假地豆混种时, 飞机草的竞争参数相对产量RY小于1, 表明飞机草与假地豆竞争时, 种间竞争大于种内竞争;同时, 飞机草对假地豆的竞争攻击力系数A显著小于0 (P < 0.05), 说明飞机草的竞争力弱于假地豆。本地种假地豆的相对产量RY显著大于1(P < 0.05), 表明假地豆与飞机草竞争时表现为种内竞争大于种间竞争, 其竞争攻击力系数A则与飞机草的相反, 显著大于0, 其竞争力强于飞机草(图 4)。

|
| 图 4 混种时飞机草与本地种的相对产量及其相互攻击力系数 Fig. 4 The relative yield and competitive aggressivity coefficient of Chromolaena odoratum and native species when they grown with each other “F+J”表示飞机草与假地豆混种, “F+B”表示飞机草与白饭树混种。*表示物种竞争时相对产量和竞争攻击力系数分别与1和0相比有显著性差异, P < 0.05 |
飞机草与白饭树混种时, 飞机草的竞争参数相对产量RY显著大于1(P < 0.05), 表明飞机草与白饭树竞争时, 种内竞争大于种间竞争;飞机草对白饭树的竞争攻击力系数A显著大于0, 表明飞机草的竞争力强于白饭树;白饭树的相对产量RY显著小于1(P < 0.05), 表明白饭树与飞机草竞争时表现为种间竞争大于种内竞争, 其竞争攻击力系数则与飞机草的相反, 显著小于0, 表明其竞争力弱于飞机草(图 4)。
3 结论与讨论 3.1 本地植物种子萌发对飞机草的化感耐受性种子萌发是植物生命周期中的关键环节, 对植物生长发育至关重要[24-25]。在入侵植物替代控制过程中, 替代植物在被入侵群落中的种子萌发能力是其在被入侵群落中能否成功繁殖后代, 从而长期持续生长的关键。研究表明多数外来入侵植物可以通过分泌化感物质抑制其他植物种子萌发, 在被入侵植物群落中, 替代目标植物可能会因其种子不能耐受入侵植物的化感作用而影响幼苗和植株的产生, 这可能是很多替代防治不能长期持久的原因之一。因此, 替代目标植物的种子萌发能力是否可以耐受入侵植物的化感作用是筛选替代目标植物的重要条件。郇树乾[26]为探讨能替代控制飞机草的牧草, 研究了热研二号柱花草(Stylosanthes guianensis cv.ReyanⅡ)种子萌发及幼苗生长对飞机草水提液的化感耐受性, 但结果显示飞机草各部位提取液对其有不同程度的化感抑制作用, 因此不能将其作为替代目标牧草。殷洁等[27]以选种合适的牧草为目的, 研究了苜蓿(Medicago sativa)、菊苣(Cichoriu mintybus)和燕麦(Avena sativa)种子萌发和幼苗生长对入侵植物空心莲子草(Alternanthera philoxeroides)的化感耐受性, 结果显示几种牧草均受到不同程度的化感抑制, 建议选种受抑制程度较小的菊苣和苜蓿以减少草场的危害和损失。李苇洁等[28]也通过研究乡土植物白刺花(Sophora davidii)的种子萌发及幼苗生长对中、低浓度紫茎泽兰水提液具有一定的化感耐受性, 揭示了其能与紫茎泽兰伴生的可能机理。本研究中除了飞机草的高浓度的叶提取液对本地植物假地豆的种子萌发率有抑制作用外, 飞机草各部位不同浓度的提取液对假地豆和白饭树的种子相对萌发率均无显著影响, 甚至在多数处理下它们的相对萌发率均有被促进的趋势, 说明这两种本地植物的种子萌发能力对飞机草的化感物质均具有一定的耐受性, 这就为其能够在飞机草入侵群落中成功繁殖后代或建立种群奠定了初步基础。
3.2 飞机草与本地植物的竞争植物的竞争是决定群落性质的主要因素[29], 研究入侵物种与本地种的竞争关系不仅是探讨外来植物成功入侵机制的一种途径, 也是寻找控制方法的重要原理和依据。在替代控制技术中, 入侵种与本地种的竞争能力研究已成为竞争取代的研究焦点之一[4, 23, 30]。不少学者根据物种的竞争原理筛选出一些具有替代控制入侵植物潜力的目标物种, 如蒋智林等[31]发现非洲狗尾草(Setaria sphacelata)能与入侵种紫茎泽兰竞争, 认为可以用其进行替代控制;马杰等[32]研究了入侵植物黄顶菊(Flaveria bidentis)与三种牧草的竞争, 认为高丹草(Sorghum bicolor×Sorghum sudanense)可以作为黄顶菊的生物替代材料;岳茂峰等[33]通过杂交狼尾草(Pennisetum americanum×P. purpureum)能与入侵种豚草竞争, 认为该草能够替代控制豚草;Shen等[34]认为甘薯(Ipomoea batatas)有较高的生长速率和与薇甘菊竞争土壤养分的能力, 可以作为其替代控制植物。最近, 岳茂峰等[9]研究了杂交狼尾草和木豆(Cajanus cajan)与飞机草的竞争, 认为它们可以替代控制飞机草。
植物的大小特征如株高和生物量是反映植物竞争能力的重要指标[35-36]。较高的株高有利于植物抢占光资源, 更好的进行光合作用, 从而积累更多的能量物质, 促进植物生长;较高生物量的积累是植物对资源吸收利用能力的体现。张震等[37]认为在替代控制中, 替代植物是否能够显著降低入侵植物的株高或生物量是取得替代成功的重要条件。本研究中, 在飞机草与本地植物假地豆混种时, 飞机草的株高显著降低, 而本地植物假地豆的株高无显著变化, 生物量却显著增加, 体现出假地豆对资源竞争的优势;混种时飞机草的相对产量显著小于1, 表现出种内竞争小于种间竞争, 其竞争攻击力系数显著小于零, 说明飞机草的竞争力弱于本地植物假地豆。在飞机草与本地植物白饭树混种时, 飞机草的株高、总生物量、地上及地下生物量均显著增加, 而白饭树的株高和生物量则显著降低, 表现出飞机草对资源利用的优势;同时, 混种时飞机草的相对产量显著大于1, 表现出种内竞争大于种间竞争, 其竞争攻击力系数显著大于零, 说明飞机草的竞争力强于本地植物白饭树。可见, 本地植物假地豆在与飞机草的竞争中表现出较强的资源竞争优势, 可以竞争抵御飞机草的生长, 而另一本地植物白饭树对飞机草的竞争力较弱, 不能竞争影响飞机草的生长。
3.3 两种本地植物替代控制飞机草的潜力除了竞争能力和具有化感耐受性外, 替代植物的覆盖性、可持续性及经济或生态价值也是替代植物筛选的重要指标[7]。覆盖性关系着替代植物是否能够占据入侵植物的生态位, 从而影响入侵植物的生长, 可持续生长的特性对于长期控制入侵植物有重要意义。假地豆是一种灌木或亚灌木, 具直立和平卧生长的特性, 叶片茂盛、枝条多而密集, 对地面的覆盖面积较大, 且种子产量较多, 易形成稳定的种群。结合假地豆对飞机草较强的生长竞争力及种子萌发对飞机草化感物质的耐受性, 在飞机草入侵地种植该植物, 一旦其种群建立就会占据大面积生态位, 从而影响飞机草生长和种群扩散, 可以长期持续阻止飞机草入侵。此外, 该植物还是一种重要的药用植物, 内服可以治疗乙型脑炎, 具一定开发价值。综合以上结果, 可以认为假地豆能够在飞机草入侵地较好的繁殖和生长, 且易形成稳定的种群, 具有较好的替代控制飞机草的潜力。
本地植物白饭树的竞争力弱于飞机草, 这可能因为飞机草是草本或木质化程度相对较低的亚灌木植物, 短时间内的资源吸收利用率较高, 植株生长较快, 而白饭树是一种木质化程度相对较高的灌木植物, 在短时间内生长相对较慢, 从而在生长初期竞争力弱于飞机草。但白饭树成株以后冠幅较大, 可形成较大荫蔽作用, 种子产量多, 易形成稳定的种群, 这在一定程度上可以影响飞机草正常开花结果, 因为飞机草生长受光照影响较大, 在光照率小于38%时便不能正常开花结果[38]。同时, 白饭树的种子萌发力不受飞机草化感物质的抑制, 甚至还会被其微弱促进, 因此, 如果在飞机草已被清除的入侵地种植该本地植物, 不会因飞机草的残留化感物质而影响其萌发繁殖, 这样一方面可以促进本地植物的恢复, 增加生物多样性, 另一方面待该植物成功建立种群, 可以通过其荫蔽作用在一定程度上预防飞机草的二次入侵。
3.4 飞机草可能的竞争策略与替代控制植物表型可塑性对植物入侵性的影响主要表现在定居、建群和扩散入侵阶段, 因为表型可塑性能增强植物对多样环境的耐受性和适应性, 增强其资源捕获与利用能力, 使之表现出快速生长和繁殖的特性, 提高其竞争力[39]。本研究中, 飞机草与两个本地种竞争时均表现为地上生物量比增加, 地下生物量比和根冠比显著降低, 在与竞争力较强的假地豆竞争时, 这些变量增加或降低的更多, 说明当存在其他物种竞争压力时, 飞机草能改变自身资源分配, 通过降低地下资源、增加地上资源投入的形式, 与物种竞争空间资源, 且种间竞争力越大, 这种趋势表现的越强, 表现出较大的表型可塑性。全国明等[40]的研究也表明飞机草能够通过植株形态、结构及生物量分配的调整来适应多变的环境, 具较大的表型可塑性, 可见, 表型可塑性可以在一定程度上协同促进飞机草的竞争。同时, 飞机草与物种竞争时的资源分配方式与入侵植物紫茎泽兰在与物种竞争时将更多的资源投入到地下部分的分配方式不同[29], 这可能是飞机草与其他物种竞争时的一种策略。针对这种策略, 在对飞机草进行防治时应该考虑主要抑制其地上部分生长, 因此, 在进行替代控制飞机草的过程中除了选用一些能与飞机草竞争的物种外, 还应再配置一些冠幅较大的阔叶灌木或乔木, 通过荫蔽影响其地上部分生长达到控制效果。
本研究结果表明, 多年生本地植物假地豆和白饭树种子萌发对飞机草的化感物质均具有一定的耐受性, 其中假地豆与飞机草混种, 可使飞机草株高显著降低, 而其自身生物量显著增加, 表现出较大竞争优势, 具有一定的替代控制飞机草的潜力;白饭树竞争能力弱于飞机草, 不能竞争抵御飞机草的生长, 但可以种植在清除飞机草后的入侵地, 促进本地植物多样性恢复和预防飞机草的二次入侵;另外, 飞机草在竞争时往往增加地上资源的投入, 这种表型可塑性可能协同促进飞机草的竞争, 对其替代控制时应考虑抑制其地上部分生长。在自然生态系统中, 替代控制的目的不仅要消除入侵物种的危害, 更重要的是修复被入侵植物破坏的生态系统, 因此, 替代控制不宜采用单一物种, 应根据实地条件选择不同的物种和替代模式。此外, 对于实验筛选的目标植物是否能够替代成功还需要进一步对其进行野外监测和评估, 这也是替代控制过程的一个重要环节。
| [1] |
马杰, 皇甫超河, 易津, 杨殿林. 4种替代植物对黄顶菊入侵土壤养分和酶活性的影响. 生态环境学报, 2011, 20(5): 805-812. DOI:10.3969/j.issn.1674-5906.2011.05.003 |
| [2] |
卢向阳, 王秋霞, 刘冰, 俞晶山, 杨万吉. 紫穗槐替代控制对撂荒山地紫茎泽兰的影响. 西南农业学报, 2013, 26(5): 1893-1898. |
| [3] |
张知晓, 泽桑梓, 季梅, 刘凌, 户连荣. 薇甘菊替代控制技术研究. 西部林业科学, 2018, 47(3): 123-128. |
| [4] |
申时才, 徐高峰, 张付斗, 李天林, 张玉华. 红薯对薇甘菊的竞争效应. 生态学杂志, 2012, 31(4): 850-855. |
| [5] |
关广清, 韩亚光, 尹睿, 瞿强, 苗青, 王文和, 王淑芝, 高东昌, 李素德, 崔松, 金会军. 经济植物替代控制豚草的研究. 沈阳农业大学学报, 1995, 26(3): 277-283. |
| [6] |
卢昌义, 胡宏友, 张明强, 钟跃庭, 郑逢中. 人工清除结合生态替代根除外来入侵植物猫爪藤. 植物保护, 2005, 31(3): 53-56. DOI:10.3969/j.issn.0529-1542.2005.03.013 |
| [7] |
卢向阳, 张锦华, 左相兵, 刘冰, 郑雅婧. 几种替代植物对入侵杂草紫茎泽兰的防控效果. 贵州农业科学, 2012, 40(6): 103-106, 109-109. DOI:10.3969/j.issn.1001-3601.2012.06.028 |
| [8] |
李委涛, 郑玉龙, 冯玉龙. 飞机草入侵种群与原产地种群生长性状的差异. 生态学报, 2014, 34(23): 6890-6897. |
| [9] |
岳茂峰, 崔烨, 冯莉, 张纯, 田兴山. 入侵植物飞机草与4种牧草的竞争效应. 生物安全学报, 2016, 25(4): 270-274. DOI:10.3969/j.issn.2095-1787.2016.04.006 |
| [10] |
黄榕娣, 蔡笃程. 飞机草的生物防除与综合利用研究进展. 杂草科学, 2009(1): 4-6. DOI:10.3969/j.issn.1003-935X.2009.01.002 |
| [11] |
全国明, 章家恩, 徐华勤, 毛丹鹃, 谢俊芳. 外来入侵植物飞机草的生物学特性及控制策略. 中国农学通报, 2009, 25(9): 236-243. |
| [12] |
凌冰, 张茂新, 孔垂华, 庞雄飞, 梁广文. 飞机草挥发油的化学组成及其对植物、真菌和昆虫生长的影响. 应用生态学报, 2003, 14(5): 744-746. DOI:10.3321/j.issn:1001-9332.2003.05.022 |
| [13] |
Mangla S, Inderjit, Callaway R M. Exotic invasive plant accumulates native soil pathogens which inhibit native plants. Journal of Ecology, 2008, 96(1): 58-67. |
| [14] |
王俊峰, 冯玉龙, 李志. 飞机草和兰花菊三七光合作用对生长光强的适应. 植物生理与分子生物学学报, 2003, 29(6): 542-548. |
| [15] |
鲁萍, 桑卫国, 马克平. 外来入侵种飞机草在不同环境胁迫下抗氧化酶系统的变化. 生态学报, 2006, 26(11): 3578-3585. DOI:10.3321/j.issn:1000-0933.2006.11.008 |
| [16] |
张黎华, 冯玉龙. 飞机草的生防作用物. 中国生物防治, 2007, 23(1): 83-88. |
| [17] |
潘玉梅, 唐赛春, 岑艳喜, 吕仕洪, 韦春强, 蒲高忠. 岩溶地区飞机草化学防治研究. 中国岩溶, 2011, 30(1): 53-58. DOI:10.3969/j.issn.1001-4810.2011.01.009 |
| [18] |
Qin R M, Zheng Y L, Valiente-Banuet A, Callaway R M, Barclay G F, Pereyra C S, Feng Y L. The evolution of increased competitive ability, innate competitive advantages, and novel biochemical weapons act in concert for a tropical invader. New Phytologist, 2013, 197(3): 979-988. DOI:10.1111/nph.12071 |
| [19] |
Zheng Y L, Feng Y L, Zhang L K, Callaway R M, Valiente-Banuet A, Luo D Q, Liao Z Y, Lei Y B, Barclay G F, Silva-Pereyra C. Integrating novel chemical weapons and evolutionarily increased competitive ability in success of a tropical invader. New Phytologist, 2015, 205(3): 1350-1359. DOI:10.1111/nph.13135 |
| [20] |
高尚宾, 张宏斌, 孙玉芳, 张国良. 植物替代控制3种入侵杂草技术的研究与应用进展. 生物安全学报, 2017, 26(1): 18-22, 102-102. DOI:10.3969/j.issn.2095-1787.2017.01.003 |
| [21] |
梁业飞, 周有旺. 白饭树提取物抗炎镇痛作用研究. 内科, 2015, 10(6): 845-847. |
| [22] |
郑丽, 冯玉龙. 紫茎泽兰叶片化感作用对10种草本植物种子萌发和幼苗生长的影响. 生态学报, 2005, 25(10): 2782-2787. DOI:10.3321/j.issn:1000-0933.2005.10.046 |
| [23] |
吕远, 王贵启, 郑丽, 倪汉文. 入侵植物黄顶菊与本地植物的竞争. 生态学杂志, 2011, 30(4): 677-681. |
| [24] |
Ishibashi Y, Koda Y, Zheng S H, Yuasa T, Iwaya-Inoue M. Regulation of soybean seed germination through ethylene production in response to reactive oxygen species. Annals of Botany, 2012, 111(1): 95-102. |
| [25] |
Barrero J M, Downie A B, Xu Q, Gubler F. A role for barley CRYPTOCHROME1 in light regulation of grain dormancy and germination. The Plant Cell, 2014, 26(3): 1094-1104. DOI:10.1105/tpc.113.121830 |
| [26] |
郇树乾. 飞机草对热研二号柱花草化感作用初步研究. 广东农业科学, 2014(1): 86-89. DOI:10.3969/j.issn.1004-874X.2014.01.021 |
| [27] |
殷洁, 隋宗明, 魏立本, 黄建国. 空心莲子草浸提液对苜蓿、菊苣和燕麦的潜在化感效应. 草业科学, 2018, 35(1): 146-156. |
| [28] |
李苇洁, 罗开源, 吴迪, 罗充. 乡土植物白刺花对紫茎泽兰化感作用的响应. 生态学报, 2017, 37(16): 5361-5367. |
| [29] |
蒋智林, 刘万学, 万方浩, 李正跃. 植物竞争能力测度方法及其应用评价. 生态学杂志, 2008, 27(6): 985-992. |
| [30] |
彭恒, 桂富荣, 李正跃, 李隽, 万方浩. 白茅对紫茎泽兰的竞争效应. 生态学杂志, 2010, 29(10): 1931-1936. |
| [31] |
蒋智林, 刘万学, 万方浩, 李正跃. 非洲狗尾草与紫茎泽兰的竞争效应. 中国农业科学, 2008, 41(5): 1347-1354. DOI:10.3864/j.issn.0578-1752.2008.05.012 |
| [32] |
马杰, 易津, 皇甫超河, 杨殿林. 入侵植物黄顶菊与3种牧草竞争效应研究. 西北植物学报, 2010, 30(5): 1020-1028. |
| [33] |
岳茂峰, 冯莉, 田兴山, 杨彩宏. 杂交狼尾草与豚草的竞争效应. 广东农业科学, 2013, 40(8): 83-86. DOI:10.3969/j.issn.1004-874X.2013.08.026 |
| [34] |
Shen S C, Xu G F, Clements D R, Jin G M, Chen A D, Zhang F Z, Kato-Noguchi H. Suppression of the invasive plant mile-a-minute (Mikaniamicrantha) by local crop sweet potato (Ipomoea batatas) by means of higher growth rate and competition for soil nutrients. BMC Ecology, 2015, 15(1): 1. DOI:10.1186/s12898-014-0033-5 |
| [35] |
Keddy P, Nielsen K, Weiher E, Lawson R. Relative competitive performance of 63 species of terrestrial herbaceous plants. Journal of Vegetation Science, 2002, 13(1): 5-16. DOI:10.1111/j.1654-1103.2002.tb02018.x |
| [36] |
李慧燕, 陈冬青, 王慧, 杨殿林, 皇甫超河. 不同混种密度下杀真菌剂对黄顶菊与反枝苋种间竞争的影响. 生态学杂志, 2015, 34(4): 1013-1018. |
| [37] |
张震, 徐丽, 王育鹏, 李丹. 葎草水浸提物对外来入侵植物喜旱莲子草营养生长的影响. 西北植物学报, 2012, 32(9): 1844-1849. DOI:10.3969/j.issn.1000-4025.2012.09.019 |
| [38] |
张建华, 范志伟, 沈奕德, 刘丽珍. 不同光照率对飞机草生长的影响. 安徽农业科学, 2009, 37(21): 9967-9968, 9975-9975. DOI:10.3969/j.issn.0517-6611.2009.21.058 |
| [39] |
Prentis P J, Wilson J R U, Dormontt E E, Richardson D M, Lowe A J. Adaptive evolution in invasive species. Trends in Plant Science, 2008, 13(6): 288-294. DOI:10.1016/j.tplants.2008.03.004 |
| [40] |
全国明, 谢俊芳, 章家恩, 毛丹鹃. 氮、磷养分对飞机草营养器官表型可塑性的影响. 生态学杂志, 2014, 33(10): 2625-2632. |