 2019, Vol. 39
2019, Vol. 39文章信息
- 王泽西, 陈倩妹, 黄尤优, 邓慧妮, 谌贤, 唐实玉, 张健, 刘洋
- WANG Zexi, CHEN Qianmei, HUANG Youyou, DENG Huini, SHEN Xian, TANG Shiyu, ZHANG Jian, LIU Yang
- 川西亚高山森林土壤呼吸和微生物生物量碳氮对施氮的响应
- Response of soil respiration and microbial biomass carbon and nitrogen to nitrogen application in subalpine forests of western Sichuan
- 生态学报. 2019, 39(19): 7197-7207
- Acta Ecologica Sinica. 2019, 39(19): 7197-7207
- http://dx.doi.org/10.5846/stxb201805271164
-
文章历史
- 收稿日期: 2018-05-27
- 网络出版日期: 2019-08-16





2. 西南野生动植物资源保护教育部重点实验室, 西华师范大学, 南充 637009
2. Key Laboratory of Southwest China Wildlife Resources Conservation(China West Normal University), Ministry of Education, Nanchong 637009, China
土壤呼吸是指土壤释放CO2的所有代谢过程, 包括3个生物学过程(土壤微生物呼吸, 土壤动物呼吸和根呼吸)和一个化学氧化过程(土壤氧化)[1]。土壤呼吸是陆地生态系统中仅次于植物光合作用的碳通量过程, 占整个生态系统呼吸量的60%—90%, 是生态系统碳循环的重要组成部分[2-3], 更是调控全球碳循环和气候变化的关键过程。森林土壤呼吸是陆地生态系统土壤呼吸的重要组成部分, 其动态变化将对全球碳平衡产生深远的影响, 全球森林过度采伐和其他土地利用变化导致土壤CO2释放的增加, 占过去两个世纪来人类活动释放的CO2总量的一半[4]。森林生态系统土壤呼吸的微小变化不仅会引起大气中CO2浓度的明显改变, 更会影响森林贮存碳能力[4], 因而在整个森林生态系统碳平衡估算中具有重要意义。土壤微生物是森林生态系统物质循环和能量流动的重要参与者[5-6]。土壤微生物在维持生态系统整体服务功能方面发挥着重要作用, 在C、N、P等元素的周转, 生物地球化学循环, 有机质的形成和土壤的碳蓄积、气体交换、污染物的降解起作重要作用[7]。土壤微生物通过分解作用把动植物和自身残体以及根系分泌物的有机C、N矿化释放到土壤中, 为植物提供生长的养分和固定到自身, 从而推动生物化学循环[8-10]。由人为活动导致的大气氮沉降, 20世纪增长了3—5倍, 目前, 约有200 Tg N/a活性氮输入陆地和海洋生态系统[11]。我国已经成为全球氮沉降最为严重的地区之一。有研究表明, 中国在21世纪最初10年平均氮沉降量为21.1kgN hm-2 a-1[12]。据估计, 在21世纪, 氮沉降绝对增加量最多最快的地区将为东亚和南亚[13]。在氮沉降增加条件下土壤呼吸表现出升高[14]、降低[15]或无变化[16] 3种响应方式。Richard等[17]通过对美国哈佛森林长达13年的模拟氮沉降试验发现, 氮沉降初期土壤呼吸速率升高, 但在长期氮沉降条件下, 土壤呼吸受到抑制。涂利华等[18]发现施氮能增加土壤微生物生物量碳氮。Bowben等[19]、Mo等[20]和李仁洪等[21]则发现施氮抑制了土壤呼吸, 这可能是因为氮沉降改变了土壤微环境, 导致了土壤微生物活性降低, 进而抑制了土壤呼吸。因此, 在不同区域、不同生态系统和氮沉降背景下, 施氮对土壤呼吸和微生物生物量的影响仍有很多不确定性。
前期团队对川西亚高山森林土壤微生物及其代谢做了相关研究, 发现川西高山森林-苔原交错带植被类型和季节动态对土壤可培养微生物数量和微生物生物量碳氮都有显著影响[22], 但海拔梯度导致的模拟增温并未对雪被期亚高山森林土壤微生物生物量碳氮及其比值产生显著影响[23]。另外, 林窗效应显著促进了川西亚高山森林土壤呼吸且有明显的季节动态[24]。而凋落物去除对土壤温度、水分以及土壤呼吸速率产生的差异均不显著[25]。目前还缺乏亚高山森林土壤微生物代谢对施氮的响应方面的研究。川西亚高山区域普遍受氮限制, 是模拟氮沉降的天然实验平台, 在此区域进行施氮是否会促进亚高山森林生态系统中土壤呼吸和微生物生物量, 未来氮沉降是否会增加微生物对土壤氮的固定, 从而改变亚高山森林生态系统的物质循环?因此, 本研究以川西亚高山森林为研究对象, 以人工施氮模拟氮沉降变化, 研究施氮对川西高山森林土壤的土壤微生物和土壤呼吸的影响, 以期为深入理解全球氮沉降背景下高山生态系统物质循环过程及其影响机制提供基础数据。
1 材料与方法 1.1 研究区域和研究样地概况研究区位于四川省理县米亚罗自然保护区, 高山森林生态系统定位研究站, 地理位置为31°43′N—31°51′N, 102°40′E—103°02′E, 地处青藏高原东缘褶皱带最外缘, 杂谷脑河上游, 是大渡河与岷江的分水岭, 为重要的江河源区。研究区所在山体具有明显的山地垂直地带性, 自河谷至山顶依次分布的植被类型有针阔混交林、暗针叶林、高山疏林灌丛、高山草甸, 4500m以上为高山荒漠和积雪带气候属于冬寒夏凉的高寒气候, 年平均气温6—12℃, 1月平均气温—8℃, 7月平均气温12.6℃, 年积温1200—1400℃, 年降水量600—1100mm, 年蒸发量1000—1900mm冬季雪被期明显, 从10月到次年4月, 长达6—7个月。试验地位于山体海拔3865m左右的四川红杉林内。
1.2 试验样地设置四川红杉(Larix mastersiana)仅分布于岷江流域高山峡谷地区, 为我国特有种。2017年4月底, 选择60年左右的四川红杉林, 在林中选取地势较为平坦的50m×50m区域作为研究样地。样地中0—10 cm土壤全碳为30.97 g/kg、全氮2. 94g/kg、全磷1. 32 g/kg、C /N 10. 53。在样地中取4个2.5m×2.5m的小区组成一个区组, 整个研究样地由3个这样的区组组成, 每个小区之间间隔约5m, 区组之间间隔约10m, 各小区, 区组间立地条件相似。每个区组的4个小区分别随机由空白对照CK、低氮N1、中氮N2、高氮N3组成。根据Zhu等[26]对中国陆地生态系统大气湿N沉降的组成、空间分布格局和影响因素的研究, 青藏高原东缘的大气湿氮年沉降大约为7.55kgN hm-2 a-1。该区域的大气氮沉降量相比东部地区较低, 我们设计了4个水平的土壤施氮控制试验CK:0 g m-2 a-1、N1:2 g m-2 a-1、N2:5 g m-2 a-1、N3:10 g m-2 a-1(CK只喷洒清水, N1喷洒浓度为0.86g/L的硝酸铵溶液, N2喷洒浓度为2.14g/L的硝酸铵溶液, N3喷洒浓度为4.27g/L的硝酸铵溶液)。从2017年4月底开始使用太阳能自动喷洒装置在每月中旬和每月底对样地进行施氮处理, 每个小区每次喷施1.2L溶液, 施氮量分别为CK:0g, N1:1.03g, N2:2.57g, N3:5.12g。每个小区中挖掘长宽各1m, 深50cm的去根样地, 方法为:将直径1m和深度分别为50 cm的土壤分层取出; 将土壤中可见根系去除; 在土坑壁及底部贴上数层塑料薄膜, 以阻隔根系的侵入; 翻动的土壤按照土壤层次顺序填满, 以减少扰动对土壤结构的影响, 然后尽可能将土壤中可见根系去除, 排除根系分解对土壤异样呼吸的影响, 压实, 恢复1个月后进行土壤呼吸的土壤全呼吸和土壤异样呼吸的测量。每个小区设置4个土壤呼吸环, 去根样地两个来测量土壤异养呼吸, 去根样地外两个来测量土壤全呼吸, 土壤呼吸PVC环规格为:内径20 cm, 高10 cm, 插入土壤深度为5cm。在土壤呼吸测量的前一天整齐地剪去环内所有植物的地上部分, 不干扰试验内的土壤。
1.3 土壤呼吸、土壤温度、土壤水分和微生物生物量的测定土壤呼吸通量采用Li-8100土壤碳通量自动测量系统(Li-Cor, Inc, NE, USA)测定。同时, 采用Li-8100所附带的土壤温度探针和水分传感器测定地下5 cm土壤温度和含水量(体积含水量, %)。土壤呼吸测定从2017年6月中旬开始至2017年10月中旬, 平均每个月中旬观测一次, 每次测量时间在10:00—14:00之间完成。采用微生物生物量的测定使用氯仿熏蒸提取法测定土壤微生物量[18]。
1.4 数据分析Q10值计算方法为:
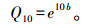
|
式中, Q10值是土壤温度敏感性系数, 即土壤温度每升高10摄氏度, 土壤呼吸速率变为未增温前呼吸速率的倍数, b是土壤呼吸与温度单因素指数曲线模型Rs=aebt中的温度反应常数(Rs为土壤呼吸速率, a为温度0 ℃时的土壤呼吸速率, b为温度反应常数)。具体方法是, 将2007年11月至2008年10月土壤呼吸速率及相对应的土壤温度进行指数分布曲线回归, 将所得的b值代入上述公式计算出各水平的Q10值[18]。
微生物生物量C、N, 微生物代谢商计算方法为:
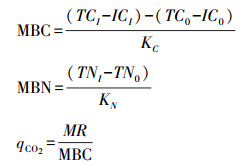
|
式中, MBC为微生物生物量碳(Microbial biomass carbon), TC1为熏蒸样品全碳, IC1为熏蒸样品无机碳, TC0为未熏蒸样品全碳, IC0为未熏蒸样品无机碳, KC为微生物生物量碳转换系数(0.45), MBN为微生物生物量氮(Microbial biomass nitrogen), TN1为熏蒸样品全氮, TN0为未熏蒸样品全氮, KN为微生物生物量氮转换系数(0.54)[18], MR为微生物呼吸。
同一采样时间不同施氮梯度对土壤各指标的影响采用单因素方差分析(one-way ANOVA), 施氮处理和采样时间对土壤呼吸、土壤微生物量、土壤温度、土壤含水率和土壤pH值的影响采用重复测量方差分析, 土壤微生物指标与环境因子的关系采用Pearson相关分析, 采用一般线性回归分析土壤呼吸通量与土壤温度和湿度的关系。利用Origin 8.5软件作图。
2 结果与分析 2.1 土壤温湿度和pH值动态从图 1可以看出, 施氮对于土壤的温度、土壤含水率、土壤pH值都没有显著的影响, 土壤温度呈现先升高后降低的趋势, 7月的时候达到最高, 10月的时候达到最低, 土壤含水率则是先降低后升高, 10月的时候最高, 7月最低。土壤pH值在时间上没有明显变化。

|
| 图 1 川西亚高山森林土壤温湿度和pH值动态 Fig. 1 Dynamics of Soil Temperature, Moisture Content, pH Valuein the Larix mastersiana of the Western Sichuan Province Time effect表示时间效应, N effect表示施氮效应, T×N effect表示时间与施氮处理的交互效应(重复测量方差分析); CK:对照组; N1:低氮组; N2:中氮组; N3:高氮组; |
从图 2可以发现, 施氮促进了生长季MBC、MBN的增长, 但变化趋势有差异。MBC随施氮浓度增加而增加, 在10月MBC的增长表现显著。随着施氮浓度的增加MBN呈现一个先增加后降低的趋势, 整个生长季MBN的增长都十分显著, 各个施氮水平MBN都显著高于对照。而微生物量碳氮比在各施氮水平在7、8月份都显著低于对照。重复测量方差分析结果表明, 施氮对于微生物生物量碳氮都有显著的影响, 对微生物量碳氮比没有显著的影响, 时间对微生物生物量碳氮及其比值都有显著的影响, 两者交互作用对微生物生物量碳氮有显著的影响, 对其比值没有显著的影响。

|
| 图 2 土壤微生物生物量碳氮对施氮的响应 Fig. 2 Soil microbial biomass carbon and nitrogen response to nitrogen application Time effect表示时间效应, N effect表示施氮效应, T*N effect表示时间与施氮处理的交互效应(重复测量方差分析);*表示施氮处理与对照组之间差异显著, **表示施氮处理与对照组之间差异极显著(单因素方差分析); CK:对照组; LN:低氮组; MN:中氮组; HN:高氮组 |
从图 3可以发现, 施氮促进了土壤全呼吸、自养呼吸、异养呼吸和微生物代谢商, 土壤全呼吸、自养呼吸、异养呼吸和微生物代谢商的变化基本一致, 随着施氮浓度的增加呈现一个先增加后降低的趋势, 其中在施加中氮水平时达到一个最大值, 土壤总呼吸在施加中氮时都显著高于对照组。在10月的时候土壤全呼吸、自养呼吸、异养呼吸显著降低, 施氮各水平与对照组相比都没有显著差异, 而微生物代谢商显著低于对照。重复测量方差分析结果表明, 施氮对于土壤全呼吸、自养呼吸、异养呼吸都有极显著的影响, 对于微生物代谢商的影响不显著, 时间对这些指标的影响也很显著, 其中对于全呼吸和微生物代谢商影响极显著, 而施氮和时间的交互效应对土壤呼吸的3个指标影响显著, 对微生物代谢商影响不显著。

|
| 图 3 土壤呼吸和土壤微生物代谢商对施氮的响应 Fig. 3 Soil Respiration and Soil Microbial Metabolizers′ Response to Nitrogen Application Time effect表示时间效应, N effect表示氮效应, T×N effect表示时间与氮处理的交互效应(重复测量方差分析);*表示氮处理与对照组之间差异显著, **表示氮处理与对照组之间差异极显著(单因素方差分析) |
Pearson相关分析表明(表 1), 土壤呼吸、微生物代谢商、微生物生物量碳氮与土壤温度呈极显著正相关, 与土壤湿度呈极显著负相关。土壤微生物氮与土壤pH值呈极显著正相关, 土壤呼吸与微生物代谢商、土壤微生物碳氮呈极显著的正相关, 微生物代谢商与土壤微生物氮呈显著的正相关。
| 项目 Project |
土壤呼吸 Soil respiration |
土壤微生物代谢熵 Soil microbial metabolic entropy |
土壤微生物生物量碳 Soil microbial biomass C |
土壤微生物生物量氮 Soil microbial biomass N |
土壤微生物生物量碳氮比 Soil microbial biomass C/N |
土壤温度 Soil temperature |
土壤湿度 Soil moisture |
土壤 pH Soil acidity |
| 土壤呼吸 Soil respiration |
1.00 | |||||||
| 土壤微生物代谢熵 Soil microbial metabolic entropy |
0.780** | 1.00 | ||||||
| 土壤微生物生物量碳 Soil microbial biomass C |
0.316** | -0.043 | 1.00 | |||||
| 土壤微生物生物量氮 Soil microbial biomass N |
0.321** | 0.191* | 0.262** | 1.00 | ||||
| 土壤微生物生物量碳氮比 Soil microbial biomass C/N |
-0.108 | -0.178 | -0.104 | -0.689** | 1.00 | |||
| 土壤温度 Soil temperature |
0.762** | 0.548** | 0.209* | 0.375** | 0.161 | 1.00 | ||
| 土壤湿度 Soil moisture |
-0.364** | -0.268** | -0.174 | -0.338** | -0.232* | -0.416** | 1.00 | |
| 土壤 pH Soil acidity |
0.001 | -0.095 | 0.247* | 0.033 | 0.060 | 0.051 | -0.30** | 1.00 |
| n=180, *表示相关性显著, **表示相关性极显著 | ||||||||
在施氮各处理下, 土壤呼吸与土壤温度之间存在呈极显著的指数回归关系(P < 0.001), 决定系数R2在0.61—0.78。根据土壤呼吸速率与土壤温度的指数回归方程, 可以计算出土壤呼吸温度敏感系数Q10值, 对照、低、中、高氮下的Q10值分别为4.26, 3.19, 7.10和4.23, Q10在中氮条件下增加最明显。对照、低、中氮处理下的土壤呼吸速率与土壤10cm含水量之间均呈显著直线回归关系( p<0.001), 对应的方程可决系数R2分别为0.41, 0.21, 0.20 (图 4)。

|
| 图 4 模型分别拟合土壤呼吸速率与土壤10cm温湿度的关系 Fig. 4 Model for relationships between soil respiration (R, μmol m-2 s-1) and soil temperature and moisture |
本研究发现, 施氮对于土壤微生物量碳氮有显著影响, 能够增加土壤微生物量碳氮, 但是微生物量碳氮在各施氮水平下的变化趋势则有差异。一般来说, 土壤的碳氮比平均值为25 : 1, 比较适合土壤微生物的生长, 因为微生物每合成1份有机物质需要利用5份碳素和1份氮素, 同时还需要利用20份碳素作为能量来源。当土壤的C/N大于25 : 1时, 微生物不能大量繁殖, 而且从有机物中释放同的氮素全部为微生物自身生长所利用。当土壤的C/N小于25 : 1时, 微生物繁殖快, 堆肥材料分解也快, 而且有多余的氮素释放, 施到土壤后供作物利用, 也有利于腐殖质形成。本研究地的土壤C/N为10.53 : 1, 土壤碳氮比对土壤微生物的生长的促进作用多发生在外源碳输入量较低的情况下, 大概土壤C/N在9.5—14左右[27],一般认为低质量的土壤有机碳限制了土壤微生物生长所需的能量来源, 从而限制了土壤有机碳的矿化速率, 新鲜有机碳的加入为微生物提供了能量和营养元素, 提高了微生物活性, 从而加速了土壤有机碳的矿化, 产生了激发效应[28]。施氮由于降低了土壤C/N比会加速土壤有机碳的矿化, 或作为能量来源而产生正激发效应[29], 因此各施氮水平都促进了土壤微生物生物量碳氮的增加。外源有机质总量对激发效应的影响更多取决于碳输入量和土壤微生物生物量碳之间的比例关系, 当碳输入量少于MBC的50%时, 矿质化反应随外源碳输入量线性增长, 当碳输入量相当于MBC的50%—200%时, 激发效应强度与碳输入量呈负指数相关, 进一步增加碳输入量则对激发效应影响较小[30]。与单独土壤有机碳输入产生的激发效应相比, 土壤碳氮比对微生物的需求在适宜、较高、较低都可抑制激发效应, 而该研究区的四川红杉林林分密度小, 郁闭度为0.7且年凋落物输入量少, 导致土壤中有机碳比其他低海拔地区含量偏低30%左右, 土壤有机氮也比低海拔地区低, 因此土壤微生物可利用的碳源偏少, 导致土壤微生物量容易受到外源氮添加的影响。
根据已有的研究发现单独施碳、施氮和碳氮复合添加都能不同程度增加微生物生物量碳氮[31]。在本实验中单独施氮使土壤微生物量碳提高了5%—9%, 土壤微生物量氮提高了23%—34%。这是因为在华中地区和华南地区氮沉降量都超过了35kgN hm-2 a-1, 而在西北、东北、内蒙古和青藏高原地区氮沉降量为7.55—12.84kgN hm-2 a-1, 川西亚高山的氮沉降量仅为7.55kgN hm-2 a-1[26], 川西亚高山土壤属于氮限制的地区, 土壤微生物量碳氮增加的原因可能是施氮增加了土壤中的可利用氮元素, 解除了微生物的氮限制, 增加了微生物对氮元素的固定, 从而显著增加了微生物生物量氮。10月份土壤微生物碳开始显著增加, 说明施氮后期对微生物碳的影响更加明显。根据方熊等的结果[32], 施氮对MBC没有明显的影响, 这是因为研究区域的氮沉降量已经与欧洲和北美一些高氮沉降区的量相当.在长期的高N沉降背景下, 尽管土壤有机物含量增加, 但不易被土壤微生物分解利用成分增多, 从而抑制了微生物生物量的增长, 这与低氮沉降量区的川西亚高山区域情况有所不同。还有人认为施氮抑制土壤微生物生物量碳是因为微生物分布广泛, 不同林地土壤微生物种类差异巨大, 氮沉降对氮饱和值响应菌种数量和种类的差异, 很可能导致施氮后微生物生物量的变化有差异, 超过微生物耐受范围的施氮必然导致微生物生物量的减少[33], 而Pregitzer等[34]在美国密歇根进行的实验表明, 长达10年的模拟氮沉降导致MBC增加;同样地, Cusack等[35]在热带进行了5年的施N实验, 发现施N显著增加了土壤中的有效氮含量, 为土壤微生物的生长和繁殖提供了丰富的营养来源, 导致MBC升高[36]。
3.2 施氮对土壤呼吸的影响本研究发现, 施氮使土壤呼吸通量提高了11%—15%, 对土壤总呼吸通量有显著影响, 随着施氮浓度的升高先增加后降低, 在中氮水平达到最高, 这也与土壤自养与异养呼吸变化趋势相似, 也与许多短期模拟氮沉降试验的结果一致[37], 短期的氮添加都能不同程度的增加土壤呼吸通量。这是因为川西亚高山地区相对处于高海拔的区域[26], 土壤普遍处于氮限制性的状态, 且目前的年氮沉降量还处于较低水平。施氮使得土壤中可用性氮增加, 缓解了亚高山森林土壤氮的限制条件, 刺激土壤微生物生物量的增长, 使其土壤微生物生物量N的固定增加[31], 这使得异养呼吸得到了增加。外源凋落物和土壤N含量的提高都会导致分解者可用性N的增加, 都会促进有机质的分解[38], 从而表现为土壤异养呼吸的增强。自养呼吸增加的原因是由于微生物活性的升高, 土壤N矿化速率加快, 促进了植物根系对N的吸收[39], 可能使根系呼吸增强, 特别是在本试验区中, 土壤碳氮比更适合土壤矿质化的发生, 另外, 氮沉降普遍可以导致衰老叶片中N元素的再分配减少[40], 从而使凋落物中的N含量增高, 并加快分解速率。施氮对土壤呼吸的影响与土壤本身的N含量有关, 土壤中可用性N的增加可能会使微生物对N的固定和植物根系对N的吸收增加, 但土壤中含N量超过一定程度, 如达到饱和状态时, N不再成为植物和微生物的限制因子, 施氮的增加促进土壤呼吸的作用将会很小, 甚至产生相反的作用, 高剂量的N施入后可能影响微生物群落, 从而产生了抑制性物质或抑制了酶的活性, 这可能是导致土壤呼吸速率下降的原因;还有可能是由于一些非生物学过程, 使一部分氮固定在土壤有机质中, 而使土壤碳释放速率减慢。这也是为什么在川西亚高山施氮对土壤呼吸呈现一个先增加后减少的影响趋势, 而在其他氮沉降水平较高的地区施氮会对土壤呼吸产生抑制作用, 本实验发现在本地区中氮(5 g m-2 a-1)为影响土壤呼吸的临界值, 低于中氮时, 施氮会刺激微生物的生长, 从而导致土壤呼吸的加强, 高于中氮时会抑制土壤微生物的活性, 从而导致土壤呼吸的降低。当然, 喷洒氮素方法和大气自然沉降氮素存在一定的差别, 喷洒氮素只是针对土壤进行处理, 大气氮沉降对整个森林生态系统都有影响, 对地上的植物和土壤都有作用, 且对整个生态系统过程的影响更为复杂, 难以通过人工控制的方式对其进行模拟实验。
3.3 土壤微生物生物量和土壤呼吸的相互关系微生微生物量的变化会导致土壤呼吸的变化, 微生物生物量的增加会导致微生物呼吸的增加, 因此施氮对土壤呼吸和微生物生物量的变化趋势类似[41]。异养呼吸实质上反映的是土壤微生物在酶系统的作用下对有机物的分解释放CO2过程, 土壤中微生物分解有机质释放CO2 的过程是一个受温度、土壤含水量、有机质含量以及氮可利用性等诸多因子影响的生物化学过程。当大量的氮素持续进入生态系统后, 可以通过改变土壤中微生物生物量、群落结构、组成以及微生物功能和活性, 从而引起土壤异养呼吸作用的改变[17]。对于N限制地区, 由于微生物受到可用性N的限制, 对有机质的分解会受到抑制。在施氮的情况下通过微生物生物量的增殖和提高微生物活性, 微生物呼吸得到增强; 温度作为土壤呼吸的重要影响因素, 在土壤呼吸研究中受到普遍关注, Q10所代表的土壤呼吸温度敏感性在模拟和预测全球碳循环过程中非常重要, 已有学者研究了氮沉降对土壤呼吸温度敏感性的影响, 在氮沉降持续增加及全球温暖化的背景下, 探索森林土壤呼吸对氮沉降的响应及其内在机理显得非常重要。本试验通过一般线性回归拟合土壤呼吸速率与土壤10 cm温湿度的关系, 发现土壤呼吸速率与土壤温度极显著的正相关, 与土壤湿度极显著负相关, 中氮水平下土壤温度敏感性系数Q10值高于对照组的Q10值。这可能是在温度升高时, 土壤微生物和土壤酶活性增强, 微生物对N的利用效率提高, 土壤有机质分解加速[42], 从而表现出土壤呼吸的增强。氮沉降和温度均可通过刺激微生物的活动而影响森林土壤CO2的排放。
季节的变化也对土壤呼吸量有显著的影响, 这是因为季节的变化造成了温度和湿度的变化, 这两个因素是影响土壤呼吸的最主要的两个因素, 很高和很低的土壤含水量会抑制土壤微生物活性, 而导致微生物呼吸的降低, 而温度通过直接影响微生物活动和间接的改变土壤湿度、输入土壤有机物的数量和质量而对土壤呼吸造成影响, 所以季节的变化对土壤呼吸也有明显的影响[43], 这也符合上面相关性分析的结果。
4 结论从短期施氮来看, 土壤的温度、土壤含水率、土壤pH值都没有显著变化;而施氮对于土壤微生物量碳氮、土壤呼吸、异养呼吸、自养呼吸通量有显著影响, 能够增加土壤微生物量碳氮;随着施氮量增加土壤呼吸先升高后降低, 在中氮水平达到最高。相关分析发现, 土壤呼吸与微生物生物量碳氮和微生物代谢商呈极显著正相关(P < 0.01), 微生物量碳氮与温度呈极显著的正相关, 与湿度呈极显著的负相关。通过一般线性回归拟合土壤呼吸速率与土壤10 cm温湿度的关系, 发现土壤呼吸速率与土壤温度呈极显著的正相关, 与土壤湿度呈极显著负相关(P < 0.001), 中氮水平下土壤温度敏感性系数Q10值(7.10)明显高于对照Q10值(4.26)。因此, 随着大气氮沉降的增加, 将显著促进西部地区受氮限制的亚高山森林生态系统土壤呼吸和微生物生物量碳氮。
| [1] |
何立平, 田茂平, 吴红, 林俊杰, 兰波. 大气氮沉降对三峡库区消落带土壤呼吸的影响. 中国环境科学, 2019, 39(3): 1132-1138. DOI:10.3969/j.issn.1000-6923.2019.03.029 |
| [2] |
陈磊, 朱广宇, 刘玉林, 上官周平, 邓蕾. 黄土高原人工油松林土壤碳氮对短期氮添加的响应. 水土保持学报, 2018, 32(4): 346-352. |
| [3] |
Hibbard K A, Law B E, Reichstein M, Sulzman J. An analysis of soil respiration across northern hemisphere temperate ecosystems. Biogeochemistry, 2005, 73(1): 29-70. DOI:10.1007/s10533-004-2946-0 |
| [4] |
李奕霏, 肖谋良, 袁红朝, 祝贞科, 王久荣, 李科林, 葛体达, 吴金水. CO2倍增对稻田土壤碳氮水解酶活性的影响. 中国环境科学, 2018, 38(9): 3474-3480. DOI:10.3969/j.issn.1000-6923.2018.09.034 |
| [5] |
陈平, 赵博, 闫子超, 杨璐, 赵秀海, 张春雨. 太岳山油松人工林土壤呼吸对模拟氮沉降的短期响应. 生态学报, 2018, 38(22): 8184-8193. |
| [6] |
闫翠侠, 杨国亮, 李典鹏, 孙约兵, 韩东亮, 徐晓龙, 朱新萍, 贾宏涛. 生物炭对干旱区绿洲农田土壤呼吸的影响. 中国农业气象, 2018, 39(9): 575-584. DOI:10.3969/j.issn.1000-6362.2018.09.003 |
| [7] |
顾峰雪, 黄玫, 张远东, 李洁, 郭瑞, 严昌荣. 氮输入对东北土壤碳蓄积氮素利用效率的影响. 生态学报, 2017, 37(8): 2770-2778. |
| [8] |
张乃莉, 郭继勋, 王晓宇, 马克平. 土壤微生物对气候变暖和大气N沉降的响应. 植物生态学报, 2007, 31(2): 252-261. DOI:10.3321/j.issn:1005-264X.2007.02.008 |
| [9] |
李超, 刘苑秋, 王翰琨, 陈琦, 邓邦良, 刘晓君, 董星宇, 张令, 郑翔, 曹雯. 庐山毛竹扩张及模拟氮沉降对土壤N2O和CO2排放的影响. 土壤学报, 2019, 561: 146-155. |
| [10] |
许淼平, 任成杰, 张伟, 陈正兴, 付淑月, 刘伟超, 杨改河, 韩新辉. 土壤微生物生物量碳氮磷与土壤酶化学计量对气候变化的响应机制. 应用生态学报, 2018, 29(7): 2445-2454. |
| [11] |
Galloway J N, Townsend A R, Erisman J W, Bekunda M, Cai Z C, Freney J R, Martinelli L A, Seitzinger S P, Sutton M A. Transformation of the nitrogen cycle:recent trends, questions, and potential solutions. Science, 2008, 320(5878): 889-892. DOI:10.1126/science.1136674 |
| [12] |
Liu X J, Zhang Y, Han W X, Tang A H, Shen J L, Cui Z L, Vitousek P, Erisman J W, Goulding K, Christie P, Fangmeier A, Zhang F S. Enhanced nitrogen deposition over China. Nature, 2013, 494(7438): 459-462. DOI:10.1038/nature11917 |
| [13] |
Reay D S, Dentener F, Smith P, Grace J, Feely R A. Global nitrogen deposition and carbon sinks. Nature Geoscience, 2008, 1(7): 430-437. DOI:10.1038/ngeo230 |
| [14] |
宋学贵, 胡庭兴, 鲜骏仁, 肖春莲, 刘文婷. 川西南常绿阔叶林土壤呼吸及其对氮沉降的响应. 水土保持学报, 2007, 21(4): 168-172, 192-192. DOI:10.3321/j.issn:1009-2242.2007.04.039 |
| [15] |
陈毅, 闫文德, 郑威, 廖菊阳, 盘昱良, 梁小翠, 杨坤. 模拟氮沉降凋落物管理对樟树人工林土壤呼吸的影响. 生态学报, 2018, 38(21): 7830-7839. |
| [16] |
朱天鸿, 程淑兰, 方华军, 于贵瑞, 郑娇娇, 李英年. 青藏高原高寒草甸土壤CO2排放对模拟氮沉降的早期响应. 生态学报, 2011, 31(10): 2687-2696. |
| [17] |
Bowden R D, Davidson E, Savage K, Arabia C, Steudler P. Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest. Forest Ecology and Management, 2004, 196(1): 43-56. DOI:10.1016/j.foreco.2004.03.011 |
| [18] |
涂利华, 胡庭兴, 黄立华, 李仁洪, 戴洪忠, 雒守华, 向元彬. 华西雨屏区苦竹林土壤呼吸对模拟氮沉降的响应. 植物生态学报, 2009, 33(4): 728-738. DOI:10.3773/j.issn.1005-264x.2009.04.011 |
| [19] |
Mo J M, Zhang W, Zhu W X, Gundersen P, Fang Y T, Li D J, Wang H. Nitrogen addition reduces soil respiration in a mature tropical forest in southern China. Global Change Biology, 2008, 14(2): 403-412. |
| [20] |
李仁洪, 涂利华, 胡庭兴, 张健, 鲁洋, 刘文婷, 雒守华, 向元彬. 模拟氮沉降对华西雨屏区慈竹林土壤呼吸的影响. 应用生态学报, 2010, 21(7): 1649-1655. |
| [21] |
Madritch M D, Hunter M D. Intraspecific litter diversity and nitrogen deposition affect nutrient dynamics and soil respiration. Oecologia, 2003, 136(1): 124-128. DOI:10.1007/s00442-003-1253-0 |
| [22] |
刘洋, 张健, 闫帮国, 黄旭, 徐振锋, 吴福忠. 青藏高原东缘高山森林-苔原交错带土壤微生物生物量碳、氮和可培养微生物数量的季节动态. 植物生态学报, 2012, 36(5): 382-392. |
| [23] |
李洪杰, 刘军伟, 杨林, 郑海峰, 刘洋, 杨万勤, 张健. 海拔梯度模拟气候变暖对高山森林土壤微生物生物量碳氮磷的影响. 应用与环境生物学报, 2016, 22(4): 599-605. |
| [24] |
杨开军, 熊莉, 杨万勤, 贺若阳, 李志杰, 庄丽燕, 谭波, 徐振锋. 林窗对川西亚高山云杉人工林土壤呼吸的影响. 应用与环境生物学报, 2016, 22(3): 505-509. |
| [25] |
熊莉, 徐振锋, 杨万勤, 殷睿, 唐仕姗, 王滨, 徐李亚, 常晨晖. 川西亚高山粗枝云杉人工林地上凋落物对土壤呼吸的贡献. 生态学报, 2015, 35(14): 4678-4686. |
| [26] |
Zhu J X, He N P, Wang Q F, Yuan G F, Wen D, Yu G R, Jia Y L. The composition, spatial patterns, and influencing factors of atmospheric wet nitrogen deposition in Chinese terrestrial ecosystems. Science of the Total Environment, 2015, 511: 777-785. DOI:10.1016/j.scitotenv.2014.12.038 |
| [27] |
陈颖, 刘玉学, 陈重军, 吕豪豪, 汪玉瑛, 何莉莉, 杨生茂. 生物炭对土壤有机碳矿化的激发效应及其机理研究进展. 应用生态学报, 2018, 29(1): 314-320. |
| [28] |
陈春梅, 谢祖彬, 朱建国. 土壤有机碳激发效应研究进展. 土壤, 2006, 38(4): 359-365. DOI:10.3321/j.issn:0253-9829.2006.04.002 |
| [29] |
Kuzyakov Y, Friedel J K, Stahr K. Review of mechanisms and quantification of priming effects. Soil Biology and Biochemistry, 2000, 32(11/12): 1485-1498. |
| [30] |
Blagodatskaya Е, Kuzyakov Y. Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure:critical review. Biology and Fertility of Soils, 2008, 45(2): 115-131. DOI:10.1007/s00374-008-0334-y |
| [31] |
陶朋闯, 陈效民, 靳泽文, 李秋霞, 黄欠如, 张佳宝. 生物质炭与氮肥配施对旱地红壤微生物量碳、氮和碳氮比的影响. 水土保持学报, 2016, 30(1): 231-235. |
| [32] |
方熊, 刘菊秀, 张德强, 刘世忠, 褚国伟, 赵亮. 降水变化、氮添加对鼎湖山主要森林土壤有机碳矿化和土壤微生物碳的影响. 应用与环境生物学报, 2012, 18(4): 531-538. |
| [33] |
Wallenstein M D, McNulty S, Fernandez I J, Boggs J, Schlesinger W H. Nitrogen fertilization decreases forest soil fungal and bacterial biomass in three long-term experiments. Forest Ecology and Management, 2006, 222(1/3): 459-468. |
| [34] |
Pregitzer K S, Burton A J, Zak D R, Talhelm A F. Simulated chronic nitrogen deposition increases carbon storage in Northern Temperate forests. Global Change Biology, 2008, 14(1): 142-153. |
| [35] |
Cusack D F, Silver W L, Torn M S, Burton S D, Firestone M K. Changes in microbial community characteristics and soil organic matter with nitrogen additions in two tropical forests. Ecology, 2011, 92(3): 621-632. DOI:10.1890/10-0459.1 |
| [36] |
张胜男, 闫德仁, 袁立敏. 森林土壤微生物分布及其功能特征研究进展. 世界林业研究, 2018, 31(5): 19-25. |
| [37] |
Hooker T D, Stark J M. Soil C and N cycling in three semiarid vegetation types:Response to an in situ pulse of plant detritus. Soil Biology and Biochemistry, 2008, 40(10): 2678-2685. DOI:10.1016/j.soilbio.2008.07.015 |
| [38] |
曾清苹, 何丙辉, 李源, 夏力文, 杨龙龙, 邓雪梅, 李川. 模拟氮沉降对重庆缙云山马尾松林土壤呼吸和酶活性的季节性影响. 环境科学, 2016, 37(10): 3971-3978. |
| [39] |
曾清苹, 何丙辉, 毛巧芝, 秦华军, 李源, 黄祺. 重庆缙云山两种林分土壤呼吸对模拟氮沉降的季节响应差异性. 生态学报, 2016, 36(11): 3244-3252. |
| [40] |
彭勇, 陈刚, 陈冠陶, 梁政, 涂利华. 模拟氮沉降对瓦屋山常绿阔叶次生林土壤呼吸的影响. 应用与环境生物学报, 2015, 21(4): 733-739. |
| [41] |
刘静, 孙涛, 程云云, 王清奎. 氮沉降和土壤线虫对落叶松人工林土壤有机碳矿化的影响. 生态学杂志, 2017, 36(8): 2085-2093. |
| [42] |
李东, 罗旭鹏, 曹广民, 吴琴, 卓玛措, 李惠梅, 杨永梅, 庞炳坤. 高寒草甸土壤异养呼吸对气候变化和氮沉降响应的模拟. 草业学报, 2015, 24(7): 1-11. |
| [43] |
田艳琴, 曾清苹, 何丙辉, 罗松平, 李源. 氮沉降对缙云山柑橘林不同季节土壤微生物群落结构的影响. 环境科学学报, 2017, 37(10): 3977-3983. |