 2019, Vol. 39
2019, Vol. 39文章信息
- 陈婷, 黄中豪, 黄乘明, 周岐海, 韦华
- CHEN Ting, HUANG Zhonghao, HUANG Chengming, ZHOU Qihai, WEI Hua
- 广西弄岗黑叶猴栖息地选择与利用
- Habitat selection and patterns of habitat use in a group of François' langurs (Trachypithecus francoisi) in Nonggang, Guangxi, China
- 生态学报. 2019, 39(18): 6908-6915
- Acta Ecologica Sinica. 2019, 39(18): 6908-6915
- http://dx.doi.org/10.5846/stxb201807011444
-
文章历史
- 收稿日期: 2018-07-01
- 网络出版日期: 2019-07-04





2. 广西珍稀濒危动物生态学重点实验室, 广西师范大学, 桂林 541006;
3. 中国科学院动物研究所动物生态与保护生物学重点实验室, 北京 100008
2. Guangxi Key Laboratory of Rare and Endangered Animal Ecology, Guangxi Normal University, Guilin 541006, China;
3. Key Laboratory of Animal Ecology and Conservation Biology, Institute of Zoology, Chinese Academy of Sciences, Beijing 100008, China
栖息地选择是动物长期适应生存环境的结果, 动物总是偏向选择最能满足自身需求的场所作为栖息地[1]。动物对栖息地的选择与利用受到诸多生态因子的影响, 如气候条件、生境特征、食物可获得性、捕食与竞争等[2]。其中, 食物的空间分布和数量对动物栖息地的选择具有重要作用[3-6]。具体来说, 动物偏向选择食物丰富的地方作为栖息地, 表现为动物对特定区域的利用频率与食物资源的数量和质量显著相关。例如Huang等[7]在比较同域分布的猕猴(Macaca mulatta)与熊猴(M. assamensis)的栖息地利用时发现, 两种动物均偏向选择喜食食物较多的区域。捕食风险也是动物栖息地利用的重要影响因素, 即避免选择捕食风险高的区域, 尽管这些区域食物分布非常丰富[8, 9]。如神农架川金丝猴(Rhinopithecus roxellana)倾向于选择原始森林作为栖息地, 因为原始森林的植被高大而密集, 更有利于躲避捕食者[4]。弄岗熊猴更喜欢待在崖壁和山顶等人类难以接近的地方, 即使在食物缺乏时期, 它们也不会下到食物相对丰富的低洼地带觅食[10]。另外, 温度调节也会对动物的生境选择产生影响, 比如太行山的猕猴在冬季倾向待在阳光充足、气温较高的阳坡面, 这可能与动物的体温调节行为有关[11]。
黑叶猴(Trachypithecus francoisi), 又名乌猿、乌叶猴, 属灵长目(Primates)、猴科(Cercopithecidae)、疣猴亚科(Colobinae)、乌叶猴属(Trachypithecus), 是国家Ⅰ级重点保护野生动物, 被中国濒危动物红皮书列为濒危种, CITES列入附录Ⅱ, 被IUCN列为濒危种, 分布于中国的广西、贵州、重庆及越南北部、老挝等地的石山地区[12]。黑叶猴的栖息地大都存在破碎化的现象。如扶绥的黑叶猴栖息地受人为活动(开垦、伐薪、放牛羊等)影响较大, 食物种类也因破碎化而变少和分散[13]。贵州麻阳河自然保护区的黑叶猴栖息地被耕地分割, 食物资源减少, 黑叶猴盗食农作物的现象时有发生[14]。而弄岗自然保护区的黑叶猴栖息地的植被保护较好, 虽然气候条件与扶绥类似, 但大部分地区植被仍保持着原始状态, 人为干扰较少[15]。
近年来, 黑叶猴的生态学研究涉及很多方面, 主要集中在食物组成与觅食行为[16-19]、活动节律和活动时间分配[20-24]、家域[20, 25, 26]、夜宿地利用[14, 20, 27-29]等。关于弄岗黑叶猴的栖息地利用也有相关报导。黄中豪等[30]发现黑叶猴对坡向的利用可能受温度调节的影响, 表现为旱季低温时, 黑叶猴会在阳光充足的东向坡面晒太阳; 另外, 黑叶猴对山体部位的利用受食物分布的影响, 猴群偏向选择喜食食物充足的山脚和山坡。Zhou等[13]还发现黑叶猴对栖息地的利用是在觅食利益和捕食风险间进行权衡的结果, 表现为频繁地利用食物少但风险低的山顶和崖壁, 但是在食物贫乏时期, 它们也会到食物丰富而风险高的山坡和山脚觅食。然而, 已有研究尚不能完整解释生态因素对黑叶猴栖息地利用的影响, 主要原因如下:Zhou等[13]研究期间与本研究期间黑叶猴的食物组成有较大差异, 而且两个时期降雨量也差异显著。降雨量与嫩叶和果实等食物部位的数量成正相关关系[31, 32], 这说明两个时期的食物可获得性可能存在较大差异。此外, Zhou等[13]没有考察温度等生态因素对栖息地利用的影响。黄中豪等[30]仅仅研究了旱季时期猴群的栖息地利用情况。上述研究对我们深刻理解该物种对石山生境的适应策略是不足够的。因此, 本文对弄岗黑叶猴栖息地的利用进行研究, 探讨黑叶猴生境利用规律及其与生态因子、食物组成的关系, 为深入分析黑叶猴对石山森林的适应策略提供依据。
1 研究地点、对象与方法 1.1 研究地点与对象本研究地点在弄岗自然保护区的弄岗片西北部。弄岗自然保护区位于广西西南部(106°42′28″—107°4′54″E, 22°13′56″—22°33′9″N), 由东(陇山)、中(弄岗)、西(陇呼)3片组成, 总面积约100 km2。保护区的地貌类型为裸露岩溶, 以峰丛洼地和峰丛谷地为主, 峰丛海拔约300—700 m。山体多悬崖峭壁, 分布有形态各异的石洞[33]。植被类型主要为石灰岩山地季雨林, 多分布热带性较强的高大乔木和藤本植物[34]。研究区域面积约100 hm2, 石山海拔为190—470 m, 碳酸盐岩广布, 水热条件较好, 除少数山尖和陡壁外, 几乎全为植被覆盖, 森林覆盖率达到96%以上[15]。研究期间(2005年9月至2006年8月), 借助温度计和量雨筒收集每天的温度与降雨量数据。结果表明研究期间平均温度为21.6℃; 平均最高温度达31.9℃, 出现在6月; 平均最低温度为8.1℃, 出现在12月。总降雨量为1372.8 mm, 具有明显的雨季和旱季。雨季为4月至9月(月降雨量> 80 mm), 降雨量占年总降雨量的87.5%;旱季为10月至3月(月降雨量 < 80 mm), 降雨量占12.5%。研究对象是一群由9个个体组成的黑叶猴, 包括1只成年雄性、5只成年雌性和3只婴幼猴。
1.2 研究方法把喀斯特石山自上而下划分为山顶(Hill-top)、崖壁(Cliff)、山坡(Hill slope)、山脚(Hill bottom)及平地(Flat zone)5个部分。山顶部分指石山的最高处, 植被稀疏, 多为矮小灌木; 崖壁部分指山体坡度为80°—90°的部分, 陡峭, 植被稀少, 仅有少量灌木; 山坡部分指山体中部, 崖壁的下部, 坡度范围在20°— 60°左右, 土壤丰富, 植被覆盖率较高, 多乔木; 山脚部分指坡度为20°左右的山体部位, 植物种类丰富, 多高大乔木、藤本植物等; 平地部分为峰林或峰丛底部, 坡度几乎等于零的地带[15, 34]。
采用瞬时扫描法(Instantaneous scan sampling)[35]每隔30 min对猴群进行一次扫描取样, 记录猴群每个个体所在山体部位, 以多数个体所处的山体部位代表该猴群所处的山体部位; 记录每个个体活动类型, 以多数个体发生的活动类型来定义该时段猴群的主要活动类型。活动类型包括休息(Resting)、移动(Moving)和觅食(Feeding)[36, 37]。由于理毛等社会行为多发生在休息时, 因此本研究中将社会行为归类为休息。用食物可获得性指数(Food Availability Index, FAI)表示环境中猴群食物的丰富程度, 计算公式为FAI=Ni/N, Ni为样方内长有嫩叶、成熟叶、花或果实等可食用部位的树木的株数, N为样方内树木的总株树。食物多样性指样方内所有可食植物种类和部位的总和, 用Shannon-Wiener指数H来表示, 计算公式为: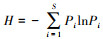
将每次扫描取样作为一个独立样本, 以每月猴群对不同山体部位利用的次数占总记录的比例作为该月对不同山体利用的比例, 再求其平均值来代表全年和不同季节的利用比例。该方法同样运用于猴群各行为模式下对山体的利用。采用Kruskal-Wallis来检验多个独立样本间的差异; 采用Mann-Whitney U来检验两个独立样本间的差异; 采用逐步回归分析的方法(Stepwise regression), 以猴群对不同栖息地部位的利用比例为因变量, 不同生态因子及食物部位组成为自变量, 分析影响黑叶猴栖息地利用的关键因子。所有检验的显著性水平设定为0.05。数据的处理和分析均在Microsoft Excel 2010和SPSS 23.0统计软件上完成。
2 结果 2.1 黑叶猴对不同山体部位的利用黑叶猴对不同山体部位的利用频率差异显著(χ2=43.063, df =4, P < 0.001)。其中, 猴群对崖壁的平均利用频率最高, 占总记录的36.67% ± 9.44%;其次是山坡, 占32.30% ± 9.57%;山脚和山顶分别占14.15% ± 5.01%和11.24% ± 8.42%;对平地的利用最少, 仅占5.63% ± 2.92%。
黑叶猴在各行为发生时对不同山体的利用有差异(图 1)(休息:χ2 =44.196, df =4, P < 0.001;移动:χ2 =33.292, df =4, P < 0.001;觅食:χ2=25.383, df =4, P < 0.001)。猴群休息时对崖壁的利用频率最高(43.56% ± 15.17%), 依次是山坡(32.39% ± 10.31%)、山顶(14.06% ± 9.94%)、山脚(8.11% ± 8.11%)和平地(1.88% ± 1.64%)。猴群移动时对山坡的利用频率最高(38.65% ± 17.02%), 依次是崖壁(32.05% ± 12.47%)、山脚(18.03% ± 9.05%)、山顶(8.3% ± 13.53%)和平地(2.97% ± 3.72%)。猴群觅食时对山脚的利用频率最高(30.33% ± 11.29%), 依次是山坡(27.48% ± 10.98%)、平地(19.07% ± 9.74%)、崖壁(18% ± 9.23%)和山顶(5.12% ± 7.92%)。

|
| 图 1 黑叶猴在各行为模式中对不同山体部位的利用 Fig. 1 Limestone hill parts used by François′ langur during various activities |
黑叶猴对不同山体部位的利用无显著季节性变化(表 1)。无论雨季还是旱季, 黑叶猴对崖壁的利用频率均最高, 依次是山坡、山脚、山顶和平地。黑叶猴在各行为模式下对山体部位的利用均无显著季节性变化(表 1)。休息时对崖壁的利用频率最高, 其次是山坡和山顶, 对山脚和平地的利用最少。移动时对山坡的利用频率最高, 其次是崖壁和山脚, 对山顶和平地的利用最少。觅食时, 雨季多利用山坡, 其次是山脚; 旱季多利用山脚, 其次是山坡; 在平地、崖壁和山顶觅食的比例均较少。
| 山顶 Hill-top |
崖壁 Cliff |
山坡 Hill slope |
山脚 Hill bottom |
平地 Flat zone |
||
| 总体Overall | 雨季Rainy season | 11.51 ± 9.13 | 36.97 ± 7.56 | 34.36 ± 10.73 | 11.64 ± 5.67 | 5.51 ± 2.46 |
| 旱季Dry season | 10.96 ± 7.63 | 36.37 ± 11 | 30.24 ± 7.72 | 16.67 ± 2.33 | 5.76 ± 3.3 | |
| Mann-Whitney U | Z=-0.160 | Z=-0.320 | Z=-0.480 | Z=-1.601 | Z=0 | |
| P=0.837 | P=0.749 | P=0.631 | P=0.109 | P=1 | ||
| 休息Resting | 雨季Rainy Season | 13.09 ± 9.76 | 42.64 ± 8.55 | 35.52 ± 10.53 | 7.27 ± 5.15 | 1.48 ± 1.23 |
| 旱季Dry Season | 15.04 ± 10.02 | 44.48 ± 19.63 | 29.25 ± 9.05 | 8.95 ± 4.39 | 2.28 ± 1.88 | |
| Mann-Whitney U | Z=-0.160 | Z=-0.480 | Z=-0.801 | Z=-0.480 | Z=-0.327 | |
| P=0.873 | P=0.631 | P=0.423 | P=0.631 | P=0.744 | ||
| 移动Moving | 雨季Rainy Season | 11.81 ± 17.71 | 29.88 ± 12.99 | 36.44 ± 19.91 | 17.65 ± 8.49 | 4.23 ± 4.63 |
| 旱季Dry Season | 4.79 ± 17.17 | 34.23 ± 11.03 | 40.87 ± 20.13 | 18.4 ± 9.1 | 1.71 ± 1.46 | |
| Mann-Whitney U | Z=-0.166 | Z=-0.801 | Z=-0.161 | Z=-0.489 | Z=-0.769 | |
| P=0.868 | P=0.423 | P=0.872 | P=0.625 | P=0.442 | ||
| 觅食Feeding | 雨季Rainy Season | 5.16 ± 7.33 | 19.19 ± 11.44 | 28.17 ± 12.02 | 24.9 ± 9.02 | 21.82 ± 8.22 |
| 旱季Dry Season | 5.08 ± 9.06 | 16.05 ± 6.17 | 26.79 ± 9.96 | 35.75 ± 11.44 | 16.32 ± 9.77 | |
| Mann-Whitney U | Z=-0.357 | Z=-0.641 | Z=-0.480 | Z=-1.441 | Z=-0.561 | |
| P=0.721 | P=0.522 | P=0.631 | P=0.150 | P=0.575 |
食物可获得性和气候条件对黑叶猴栖息地利用有重要影响。逐步回归分析表明, 花的可获得性对山脚的选择有重要影响, 表现为当花的可获得性降低时, 黑叶猴增加对山脚的利用(r=-0.662, P=0.019)。黑叶猴休息时生态因子对山体利用没有显著影响。移动时, 嫩叶的可获得性对崖壁的选择起重要作用, 表现为当嫩叶可获得性降低时, 黑叶猴增加对崖壁的利用(r=-0.606, P=0.037)。觅食时, 果实的可获得性和平均最低温度是影响平地利用的关键生态因子; 当果实的可获得性提高、月平均气温降低时, 黑叶猴会增加对平地的利用频率(果实可获得性:r=1.103, P=0.003;平均最低温度:r=-0.705, P=0.029)。
2.4 不同山体部位的利用与食物部位组成的关系黑叶猴对不同山体部位的利用与食物组成有密切关系。采用逐步回归分析猴群对不同山体的利用与食物部位组成及食物多样性的关系时发现, 花是影响山顶利用的关键因子, 表现为当猴群利用山顶的频率提高, 采食花的比例也提高(r=0.697, P=0.012);果实+种子是影响山脚利用的关键因子, 表现为当猴群利用山脚的频率提高, 会采食较多的果实和种子等高质量食物(r=0.648, P=0.023)。各行为模式下, 影响不同山体利用的关键因子也不同。休息时, 花和成熟叶是影响山顶利用的关键因子, 表现为当猴群在山顶休息的频率增加, 对花和成熟叶的觅食比例也增加(花:r=0.879, P=0.006;成熟叶:r=0.559, P=0.050);果实+种子是影响山脚利用的关键因子, 当猴群提高在山脚休息的频率时, 会提高采食果实+种子的比例(r=0.608, P=0.036)。移动时对山体部位的利用与食物部位组成没有显著相关关系。觅食时, 果实+种子是影响山坡利用的关键因子, 表现为当猴群在山坡觅食的频率提高, 会降低对果实+种子的觅食比例(r=-0.664, P=0.018);嫩叶是影响山脚利用的关键因子, 表现为当猴群利用山脚的频率提高, 嫩叶的觅食比例降低(r=-0.614, P=0.034)。
3 讨论 3.1 黑叶猴对山体的选择与利用黑叶猴对崖壁和山坡的利用频率较高, 并将崖壁作为主要休息场所, 山坡和山脚为主要移动和觅食场所, 对山顶、平地的利用较少。崖壁地势较高, 视野开阔, 有利于在休息时及时发现潜在危险; 而山坡和山脚植物种类丰富, 植被葱郁, 不但利于觅食还有遮蔽性强的特点。该结果与Zhou等[13]对同一群黑叶猴的山体利用研究结果存在差异(表 2)。Zhou等[13]表明, 黑叶猴对山顶崖壁的利用显著高于山坡山脚, 且很少下到平地觅食, 可能是两个时期的食物资源和气候存在差异。从降雨量来看, 本研究的年降雨量为1372.8 mm, 高于Zhou等[13]研究期间的977 mm。降雨量与嫩叶、果实等食物部位的数量呈明显正相关关系[31, 32]。这说明本研究时期的食物资源总量可能多于Zhou等[13]的研究时期。从食物组成上看, 本研究期间猴群采食的果实较少, 没有显示出对山脚利用频率的增加, 主要原因在于食物种类的分布格局不同。据周岐海[39]描述, 旱季时山坡的树叶都变黄或凋落, 猴群只能冒险到山脚平地大量采食围涎树(Pithecellobium clypearia)的果实和种子。围涎树主要分布在山脚平地, 能提供大量的果实和种子, 是猴群旱季时期的主要替代食物[39]。这种大斑块状分布格局使黑叶猴不需要花费太多时间就能采食到果实和种子, 随后快速回到崖壁休息, 因此尽管猴群对果实的采食量增加, 但是对平地的利用频率却没有显著提高。Zhou等[13]还指出, 黑叶猴倾向选择山顶崖壁可能是以前存在偷猎现象时造成的; 虽然后来偷猎现象几乎绝迹, 但是依赖山顶崖壁的习性保留了下来。而本研究中黑叶猴较于Zhou[13]时期表现出对山体利用的坡位下移, 可能与食物资源的分布有关。与Zhou[13]时期相比, 本研究黑叶猴旱季时对成熟叶的采食量显著增加[40]。石山森林中, 乔木主要分布在山体的中下部[34], 大量采食树叶可能会增加猴群对山体中部的利用频率。生活在扶绥的黑叶猴也表现出偏向利用山顶的现象, 但原因与Zhou等[13]的研究有差异, 主要原因是山顶的喜食食物密度要显著高于山坡[41]。金佛山的黑叶猴则表现出与本研究相同的结果[42], 它们偏向选择山体的中间位置而不喜欢偏上和偏下的位置, 可能原因是资源普遍分布在山体中部。同样的结果还出现在其他灵长类上, 例如生活在云南塔城的滇金丝猴(Rhinopithecus bieti)也偏向选择山体的中部, 对上、下部表现出明显的回避, 主要由于中部既能满足食物需求, 又能保证猴群的安全[5]。邻近同一类型生境中的白头叶猴(Trachypitheucs leucocephalus)也表现出对山体中部利用较多的现象, 对山顶和平地的利用率几乎为零, 同样也是因为山体中部植物类型丰富, 同时具有遮蔽性强、利于觅食的特点[1]。
| 地点 Site |
平均温度 Average temperature/℃ |
年降雨量 Annual rainfall/mm |
山体海拔 Elevation/m |
主要利用山体部位 Hill zone used |
参考文献 References |
| 广西弄岗 | 21.6 | 1372.8 | 300—700 | 坡中上位 | 本研究This study |
| 广西弄岗 | 22 | 977 | 300—700 | 坡上位 | Zhou等[13]; Zhou等[18] |
| 广西扶绥 | 22 | 1022 | 400—600 | 坡上位 | 周岐海等[41] |
| 重庆金佛山 | 12.5 | 1395 | 600—1800 | 坡中位 | 韩宗先等[42] |
| 贵州麻阳河 | 18.3 | 1139 | 280—1441 | 坡中下位 | Zeng等[43] |
黑叶猴对栖息地的利用在很大程度上由觅食利益决定, 这与其他黑叶猴栖息地利用的研究结果相似[13, 41]。黑叶猴以树叶为主食, 喜食嫩叶; 但是在食物缺乏时期, 猴群会采食分布广泛但营养价值较低的成熟叶来替代嫩叶[16]。这时黑叶猴会增加在崖壁休息的时间, 并通过减少移动来保存能量[36]。当食物可获得性升高, 黑叶猴会选择到山脚甚至平地这些植物种类丰富的地方觅食, 尤其是当果实等高质量食物出现时, 黑叶猴会显著提高对平地的利用。对山体利用与食物部位组成的关系进行分析时也发现, 黑叶猴对山脚的利用频率越高, 采食的果实和种子越多。黑叶猴采食的果实和种子多来源于围涎树、铁屎米(Canthium dicoccum)、鸡皮果(C. anisum)等[38, 39], 而这些植物多分布于山脚和平地, 所以极可能是这些植物的果实和种子吸引猴群过来觅食。同样的现象也出现在同域分布的熊猴上, 熊猴的主要食物为芸香竹(Indocalamus calcicolus), 这种植物主要分布在崖壁和山顶, 而研究结果也显示熊猴多在崖壁上觅食[10]。Yamada和Muroyama[44]比较了分别生活在食物充裕和食物贫乏区域的两群日本猕猴(Macaca fuscata)的生境选择, 发现两者具有很大差异:生活在食物贫乏区域的日本猕猴更频繁地到人工林与人类居住区的邻接处觅食, 而生活在食物充裕区域的日本猕猴几乎不会到该邻接处寻找食物。
捕食风险也可能是影响黑叶猴栖息地利用的重要因素之一, 并且存在觅食利益和捕食风险之间的权衡。该结论在其他黑叶猴相关研究中也有体现[13, 30, 41, 45]。当山顶上花的可获得性升高, 黑叶猴会减少对山脚的利用, 且对山顶的利用还随着成熟叶的采食比例的增加而增加, 这表明当猴群在山顶找到替代喜食的食物时, 它们更倾向在低风险的山顶的上觅食。成熟叶虽然口感和营养不佳, 但数量多且分布广, 采食成熟叶是黑叶猴应对食物种类短缺的策略[38]。猴群对山脚的利用频率随着果实和种子的采食比例的增加而增加; 当山顶上的食物减少时, 猴群会增加到风险高的山脚觅食的频率。同样结果也出现在同一地区不同时期的黑叶猴群上。Zhou等[13]的结果表明, 黑叶猴较多的利用山顶崖壁, 尽管不具有觅食的优势, 但是安全性较高; 只有在嫩叶和果实缺乏的旱季, 黑叶猴才会冒险到安全性低的山腰、平地觅食。Zeng等[43]对麻阳河黑叶猴的研究也表明, 猴群在选择栖息地时虽然倾向于靠近水源, 但是不会靠近农田, 除非在食物资源短缺的季节, 当人类或其他威胁不存在时, 才到农田寻找食物。
综上所述, 黑叶猴选择崖壁作为主要的休息场所, 而山坡和山脚是主要的移动和觅食场所。黑叶猴对栖息地的选择受到花、果实等食物可获得性的影响, 并与食物部位组成, 尤其是花、果实和种子、成熟叶有显著相关性。黑叶猴对栖息地的选择是在觅食利益与风险之间进行权衡的结果。
致谢: 黄恒善对野外数据收集提供帮助, 弄岗国家级保护区管理局对研究给予支持, University of Illinois的Paul A. Garber教授帮助写作,特此致谢。| [1] |
黄乘明, 薛跃规, 韦毅, 李友邦. 白头叶猴栖息环境与栖息地选择的研究. 兽类学报, 2000, 20(3): 180-185. DOI:10.3969/j.issn.1000-1050.2000.03.003 |
| [2] |
张洪茂, 胡锦矗. 川西北高原藏原羚夏季生境选择. 四川动物, 2002, 21(1): 12-15. DOI:10.3969/j.issn.1000-7083.2002.01.004 |
| [3] |
Apolloni N, Grüebler M U, Arlettaz R, Gottschalk T K, Naef-Daenzer B. Habitat selection and range use of little owls in relation to habitat patterns at three spatial scales. Animal Conservation, 2018, 21(1): 65-75. |
| [4] |
Li Y M. The effect of forest clear-cutting on habitat use in Sichuan snub-nosed monkey (Rhinopithecus roxellana) in Shennongjia Nature Reserve, China. Primates, 2004, 45(1): 69-72. DOI:10.1007/s10329-003-0059-3 |
| [5] |
黎大勇, 彭正松, 任宝平, Grüter C C, 周歧海, 魏辅文. 塔城滇金丝猴初秋对生境的选择性. 西华师范大学学报:自然科学版, 2006, 27(3): 233-238. DOI:10.3969/j.issn.1673-5072.2006.03.001 |
| [6] |
Zhou Q H, Huang Z H, Wei H, Huang C M. Variations in diet composition of sympatric Trachypithecus francoisi and Macaca assamensis in the limestone habitats of Nonggang, China. Zoological Research, 2018, 39(4): 284-290. DOI:10.24272/j.issn.2095-8137.2018.046 |
| [7] |
Huang Z H, Huang C M, Wei H, Tang H X, Krzton A, Ma G Z, Zhou Q H. Factors influencing positional behavior and habitat use of sympatric macaques in the limestone habitat of Nonggang, China. International Journal of Primatology, 2015, 36(1): 95-112. DOI:10.1007/s10764-015-9813-9 |
| [8] |
Enstam K L, Isbell L A. Microhabitat preference and vertical use of space by Patas monkeys (Erythrocebus patas) in relation to predation risk and habitat structure. Folia Primatologica, 2004, 75(2): 70-84. DOI:10.1159/000076265 |
| [9] |
Cowlishaw G. Trade-offs between foraging and predation risk determine habitat use in a desert baboon population. Animal Behaviour, 1997, 53(4): 667-686. DOI:10.1006/anbe.1996.0298 |
| [10] |
Zhou Q H, Wei H, Huang Z H, Krzton A, Huang C M. Ranging behavior and habitat use of the Assamese macaque (Macaca assamensis) in limestone habitats of Nonggang, China. Mammalia, 2014, 78(2): 171-176. |
| [11] |
谢东明, 路纪琪, 吕九全. 太行山猕猴的冬季生境选择. 兽类学报, 2009, 29(3): 252-258. DOI:10.3969/j.issn.1000-1050.2009.03.004 |
| [12] |
王应祥, 蒋学龙, 冯庆. 中国叶猴类的分类、现状与保护. 动物学研究, 1999, 20(4): 306-315. DOI:10.3321/j.issn:0254-5853.1999.04.012 |
| [13] |
Zhou Q H, Luo B, Wei F W, Huang C M. Habitat use and locomotion of the François' langur (Trachypithecus francoisi) in limestone habitats of Nonggang, China. Integrative Zoology, 2013, 8(4): 346-355. DOI:10.1111/j.1749-4877.2012.00299.x |
| [14] |
郭宝华, 张明海, 李迪强, 王彬. 贵州麻阳河国家级自然保护区黑叶猴夜宿地选择的研究. 野生动物杂志, 2009, 30(2): 64-69. |
| [15] |
广西壮族自治区林业厅. 广西自然保护区. 北京: 中国林业出版社, 1993.
|
| [16] |
Huang C M, Wu H, Zhou Q H, Li Y B, Cai X W. Feeding strategy of François' langur and white-headed langur at Fusui, China. American Journal of Primatology, 2008, 70(4): 320-326. DOI:10.1002/ajp.20490 |
| [17] |
Hu G. Dietary breadth and resource use of François' langur in a seasonal and disturbed habitat. American Journal of Primatology, 2011, 73(11): 1176-1187. DOI:10.1002/ajp.20985 |
| [18] |
Zhou Q H, Wei F W, Li M, Huang C M, Luo B. Diet and food choice of Trachypithecus francoisi in the Nonggang Nature Reserve, China. International Journal of Primatology, 2006, 27(5): 1441-1460. DOI:10.1007/s10764-006-9082-8 |
| [19] |
Li Y B, Ding P, Huang C M, Jiang P P, Wood C. Dietary response of a group of François langur Trachypithecus francoisi in a fragmented habitat in the county of Fusui, China:implications for conservation. Wildlife Biology, 2009, 15(2): 137-146. DOI:10.2981/08-006 |
| [20] |
Zhou Q H, Huang C M, Li Y B, Cai X W. Ranging behavior of the François' langur (Trachypithecus francoisi) in the Fusui Nature Reserve, China. Primates, 2007, 48(4): 320-323. DOI:10.1007/s10329-006-0027-9 |
| [21] |
Zhou Q H, Wei F W, Huang C M, Li M, Ren B P, Luo B. Seasonal variation in the activity patterns and time budgets of Trachypithecus francoisi in the Nonggang Nature Reserve, China. International Journal of Primatology, 2007, 28(3): 657-671. DOI:10.1007/s10764-007-9144-6 |
| [22] |
罗杨, 张明海, 马建章, 吴安康, 汪双喜, 张树森. 干扰生境下黑叶猴日活动时间的分配. 生态学报, 2007, 27(5): 1715-1722. DOI:10.3321/j.issn:1000-0933.2007.05.006 |
| [23] |
周岐海, 黄乘明, 李友邦. 黑叶猴活动时间季节性变化. 动物学杂志, 2007, 42(1): 67-73. DOI:10.3969/j.issn.0250-3263.2007.01.010 |
| [24] |
黄乘明, 周岐海, 李友邦, 蔡湘文, 魏辅文. 广西扶绥黑叶猴活动节律和日活动时间分配. 兽类学报, 2006, 26(4): 380-386. DOI:10.3969/j.issn.1000-1050.2006.04.010 |
| [25] |
陈正仁, 罗阳, 汪双喜. 贵州麻阳河黑叶猴家域及影响因素初探. 贵州林业科技, 2001, 29(2): 34-37. |
| [26] |
黄中豪, 周岐海, 黄乘明, 蒙渊君, 韦华. 广西弄岗黑叶猴的家域和日漫游距离. 兽类学报, 2011, 31(1): 46-54. |
| [27] |
Zhou Q H, Huang C M, Li M, Wei F W. Sleeping site use by Trachypithecus francoisi at Nonggang Nature Reserve, China. International Journal of Primatology, 2009, 30(2): 353-365. DOI:10.1007/s10764-009-9348-z |
| [28] |
Wang S L, Luo Y, Cui G F. Sleeping site selection of Francois's langur (Trachypithecus francoisi) in two habitats in Mayanghe National Nature Reserve, Guizhou, China. Primates, 2011, 52(1): 51-60. DOI:10.1007/s10329-010-0218-2 |
| [29] |
韩家亮, 胡刚. 麻阳河自然保护区黑叶猴夏季的夜宿行为. 兽类学报, 2012, 32(4): 362-367. |
| [30] |
黄中豪, 唐小平, 蒙渊君, 唐华兴, 黄乘明, 周岐海. 黑叶猴在旱季对喀斯特石山生境的利用. 四川动物, 2013, 32(1): 12-18. DOI:10.3969/j.issn.1000-7083.2013.01.003 |
| [31] |
Tsuji Y, Hanya G, Grueter C C. Feeding strategies of primates in temperate and alpine forests:comparison of Asian macaques and colobines. Primates, 2013, 54(3): 201-215. DOI:10.1007/s10329-013-0359-1 |
| [32] |
Ting S, Hartley S, Burns K C. Global patterns in fruiting seasons. Global Ecology and Biogeography, 2008, 17(5): 648-657. DOI:10.1111/j.1466-8238.2008.00408.x |
| [33] |
黄乘明, 李友邦, 周岐海. 白头叶猴对喀斯特石山生境的适应. 生物学通报, 2016, 51(1): 1-4. |
| [34] |
苏宗明, 赵天林, 黄庆昌. 弄岗自然保护区植被调查报告. 广西植物, 1988(S1): 185-214. |
| [35] |
Altmann J. Observational study of behavior:sampling methods. Behaviour, 1974, 49(3/4): 227-267. |
| [36] |
黄中豪, 周岐海, 李友邦, 韦显盛, 韦华, 黄乘明. 弄岗黑叶猴的日活动类型和活动时间分配. 动物学报, 2007, 53(4): 589-599. DOI:10.3969/j.issn.1674-5507.2007.04.002 |
| [37] |
Li Z Y, Rogers E. Habitat quality and activity budgets of white-headed langurs in Fusui, China. International Journal of Primatology, 2004, 25(1): 41-54. DOI:10.1023/B:IJOP.0000014644.36333.94 |
| [38] |
黄中豪, 黄乘明, 周岐海, 韦华, 蒙渊君. 黑叶猴食物组成及其季节性变化. 生态学报, 2010, 30(20): 5501-5508. |
| [39] |
周岐海.弄岗自然保护区黑叶猴(Trachypithecus francoisi)对喀斯特石山生境的行为适应[D].北京: 中国科学院动物研究所, 2005.
|
| [40] |
Zhou Q H, Huang Z H, Wei X S, Wei F W, Huang C M. Factors influencing interannual and intersite variability in the diet of Trachypithecus francoisi. International Journal of Primatology, 2009, 30(4): 583-599. DOI:10.1007/s10764-009-9362-1 |
| [41] |
周岐海, 蔡湘文, 黄乘明. 黑叶猴对广西扶绥破碎生境的选择和利用. 动物学研究, 2010, 31(4): 421-427. |
| [42] |
韩宗先, 王维, 胡锦矗. 重庆金佛山黑叶猴的春季生境选择. 林业科学, 2011, 47(4): 121-128. |
| [43] |
Zeng Y J, Xu J L, Wang Y, Zhou C F. Habitat association and conservation implications of endangered Francois' langur (Trachypithecus francoisi). PLoS One, 2013, 8(10): e75661. DOI:10.1371/journal.pone.0075661 |
| [44] |
Yamada A, Muroyama Y. Effects of vegetation type on habitat use by crop-raiding Japanese macaques during a food-scarce season. Primates, 2010, 51(2): 159-166. DOI:10.1007/s10329-009-0183-9 |
| [45] |
周岐海, 蔡湘文, 黄乘明, 李友邦, 罗亚平. 黑叶猴在喀斯特石山生境的觅食活动. 兽类学报, 2007, 27(3): 243-248. DOI:10.3969/j.issn.1000-1050.2007.03.006 |