 2019, Vol. 39
2019, Vol. 39文章信息
- 陈林, 辛佳宁, 苏莹, 李月飞, 宋乃平, 王磊, 杨新国, 卞莹莹, 田娜
- CHEN Lin, XIN Jianing, SU Ying, LI Yuefei, SONG Naiping, WANG Lei, YANG Xinguo, BIAN Yingying, TIAN Na
- 异质生境对荒漠草原植物群落组成和种群生态位的影响
- Effects of heterogeneous habitats on community composition and niche characteristics of different plant populations in the desert steppe of China
- 生态学报. 2019, 39(17): 6187-6205
- Acta Ecologica Sinica. 2019, 39(17): 6187-6205
- http://dx.doi.org/10.5846/stxb201810182255
-
文章历史
- 收稿日期: 2018-10-18
- 网络出版日期: 2019-05-31





2. 宁夏大学西北土地退化与生态恢复国家重点实验室培育基地, 银川 750021;
3. 宁夏大学农学院, 银川 750021
2. Key Laboratory for Restoration and Reconstruction of Degraded Ecosystem in Northwest China of Ministry of Education, Ningxia University, Yinchuan 750021, China;
3. School of Agriculture, Ningxia University, Yinchuan 750021, China
在自然生态系统中, 植物的生存离不开特定的栖息地, 其生长、发育与周围环境密切相关[1], 环境因素对物种分布、群落结构、生物多样性等有着很大的贡献[2], 也是长期作用的结果[3]。生境(habitat)是指植物或群落生长的具体地段的环境因子的综合[4], 一般是指所占有的资源(如养分、水份等)、环境条件(温度、雨量等)和使这个物种能够存活和繁殖的空间。而生境异质性现象普遍存在于自然生态系统中[5], 往往造成资源可利用性的差异, 会导致物种的生态位分化, 进而形成植被特定的空间分布格局[6]。因此在一定的地域范围内, 生境及其构成要素的丰富与否, 很大程度上影响甚至决定着生物的多样性。有研究表明生境异质性与物种多样性之间存在正相关关系[7], 特别是在干旱半干旱地区, 小尺度生境异质性是导致群落组成不同[8]和物种共存的重要因素[9], 其中, 土壤类型的差异扮演着重要的角色[10-11]。而阐明植物群落组成对生境异质性的响应是群落生态学研究的核心议题之一[6]。
一般认为种群间相互关系可以通过植物利用资源的状况反映[12], 因此资源利用状况是认识群落组成形成机制的主要问题, 而生态位理论可以揭示物种间对可利用资源的量化问题[13]。经典的生态位理论认为, 物种在某种(或多种)环境资源利用上存在差异, 即物种间的生态位分化, 这是物种共存的基本机制之一[14], 也是群落组成变化和群落演替的主要动力[15]。当然, 除生境中的可利用资源外, 其他因素也影响生态位分化, 如干扰[16]和空间[17]。荒漠草原是草原向荒漠过渡的一个最干旱的草原生态系统类型, 植物群落结构简单, 物种组成、植被类型与分布等往往容易发生变化, 对外界扰动非常敏感, 长期受人为和自然干扰, 引起了生境破碎化, 具有生态脆弱性和不稳定性, 承受着巨大的压力[18-19]。围栏封育是保护退化草地植被简便有效和便于推广应用的技术措施之一, 能够促进荒漠化物种对资源利用的能力和生态位功能的发挥, 进而使草地植物群落的组成和功能发生规律性的变化, 草地生态系统在自身弹性下得以恢复和重建[20]。宁夏荒漠草原原生硬质灰钙土斑块散布在广大沙化土地中, 形成类似“土岛”的土被结构[18], 部分裸露的基岩风化物也呈嵌套状分布, 水分、养分等可利用资源数量贫乏且时空不均、人为干扰不断等影响物种共存的因素较多, 植被常呈现出不连续斑块状分布, 用生态位理论解释物种共存也许能得到较好的回答。
目前, 许多学者在荒漠草原开展了大量的研究工作, 涉及的内容主要集中在生物多样性及共存机制[18, 21-23]、放牧影响及管理[24-26]、退化机理及恢复措施[27-30]等方面, 但针对围栏和放牧条件下不同土壤类型生境中植物群落组成和主要种群生态位特征的研究尚少见相关报道。因此, 本研究基于经典的生态位理论, 通过调查荒漠草原植物群落数量特征, 分析以上异质生境中主要种群的生态位宽度和生态位重叠, 以期回答以下问题:(1)荒漠草原植物群落种类组成及其数量特征对异质生境的响应;(2)异质生境群落中各植物种群的生态位是否相同以及优势种的生态位特征发生怎样的变化。希望通过此研究, 能够对揭示植物种群间的相互关系、物种的生态适应性和共存机制、深入研究该区域异质生境下群落演替及植被恢复和保护提供科学依据。
1 研究地区与方法 1.1 研究区概况研究区位于宁夏回族自治区盐池县皖记沟村, 是典型的中温带大陆性气候, 近50年平均气温为8.46℃, 无霜期150 d, 年日照时数为2862.6 h, 年降水量为276.3 mm, 主要集中在7—9月。常见植物以旱生、中旱生类型为主。地貌为缓坡丘陵与薄层沙性黄土覆盖组合。在以风蚀为主的现代侵蚀作用下, 形成了原生黄土斑块与次生沙地和风化基岩镶嵌分布的格局[18]。土壤类型以灰钙土为主, 其次为风沙土和基岩风化沉积土。研究区由于地形地貌的复杂性、土壤类型分布的斑块性、植被恢复的脆弱性以及人类活动的破坏性等, 成为了许多学者的研究热点区域。
1.2 试验设计以荒漠草原不同生境(围栏内外下的不同土壤类型)的植物群落为研究对象, 于2018年7—8月进行样方调查, 考虑到依据样方中个别土样反映整个样方的土壤属性, 其结果存在着很大的不确定性[31], 因此, 本研究在风沙土、灰钙土和基岩风化沉积土生境下, 采用代表性样地法分别选择不同围封年限和不同放牧程度的群落样地。其中, 围封年限1—2 a为短期围封样地, 10—12 a为长期围封样地;放牧程度参照任继周[32]对放牧强度的划分标准, 采用放牧农户调查与野外实地观察相结合的方式, 划分出轻度放牧(0.30—0.45只羊/hm2)、中度放牧(1.00—1.20只羊/hm2)和重度放牧(1.45—1.50只羊/hm2)。每个生境下选择具有代表性、面积为100 m×100 m的样地, 经过统计, 风沙土生境下共布设15个样地(短期围封, 长期围封, 轻度放牧, 中度放牧和重度放牧样地各3个), 灰钙土生境下共布设12个样地(短期围封, 长期围封, 轻度放牧和中度放牧样地各3个), 基岩风化沉积土生境下共布设4个样地(短期围栏和中度放牧样地各2个)。每个样地随机布设5个1 m×1 m的草本样方, 各样方间的距离不低于10 m。调查并记录各样方盖度和植物种类, 以及各植物种的高度、盖度、密度、频度和生物量等。高度为植物的自然高度;盖度测量选择常用的目测估算法;密度为实际测量值;频度为各样地随机设置20个直径为20 cm的圆圈, 统计其出现的植物种类;将每个样方内不同种的植物齐地面刈割后, 分别装入信封带回实验室, 在(105±2)℃烘箱内杀青半小时后将温度调至(65±2)℃烘干至恒重, 用万分之一天平称重, 记为生物量。
1.3 测定项目与方法生态位测定的基本步骤是以资源轴的梯度进行划分。通常有两种不同的做法[33]:一是对资源类型根据实际测量的数据, 分成不同间隔的梯度, 然后将调查数据归入各个资源梯度进行计算;另一种是把群落调查的每个样方作为多种资源的综合状态, 以各个种在不同样方的重要值等指标进行计算, 可以认为这综合反映了不同物种对多种资源的利用, 也反映了不同物种的空间关系。基于此, 本研究以所调查的样方作为资源状态, 样方数作为资源梯度的数目进行生态位相关指标的计算。
1.3.1 重要值重要值可确定各群落的优势种, 能较客观地表达不同植物在群落中的作用与地位。以群落中物种的盖度、密度、高度和频度作为单项指标确定相对盖度(RC)、相对密度(RA)、相对高度(RH)和相对频度(RF), 重要值(IV)计算方法如下:
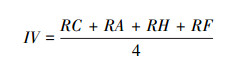
|
式中, 



选用物种丰富度(S)、Shannon-Wiener多样性指数(H)和Pielou均匀度指数(E)进行植物群落物种多样性的测定, 其计算公式为:

|
式中, Pi为样方中物种i在群落中的重要值(IV)。
1.3.3 生态位宽度生态位宽度是描述物种对环境资源多样性利用的一种表示方法, 利用群落调查资料, 采用Levins生态位宽度(Bi)表示, 计算公式如下。

|
式中, nij表示物种i利用资源状态j的数量(本研究以物种i在第j个样方的重要值表示), Ni为物种i的总量(样地中物种i的重要值和), r为样方数。指数Bi值越大, 说明生态位越宽, 该种利用的资源总量越多, 竞争力越强。
1.3.4 生态位总宽度生态位总宽度(TB)计算公式如下:
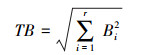
|
式中, TB为物种在第i个样地的生态位总宽度, r为样地的数量。
1.3.5 生态位重叠生态位重叠值(NO)是指一定资源序列上, 2个或者多个物种利用同等级资源而相互重叠的情况, 是一种生态特性, 不仅能反映不同物种在某些生态因子需求上的相似程度, 又能反映不同种群同时利用相同资源的状况[34]。高的生态位重叠意味着种间有一部分环境资源是共同利用的, 可能存在资源利用性竞争。主要有Pianka生态位重叠和Petraitis生态位重叠等, 本文采用方法相对简单、结果对称的Pianka生态位重叠公式计算:

|
式中, nij和nkj分别为物种i和物种k在资源j上的IV, 该方程的值域为[0, 1]。
1.3.6 生态位重叠值的总平均值样地全部种群间生态位重叠值的总平均值(TANO)的计算公式如下:
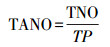
|
式中, TNO为样地内全部种群间生态位重叠值总数, TP为总种对数。
1.3.7 DCA排序/聚类分析物种的分布和数量与生态位宽度和重叠值、物种的聚类和排序均有着密切相关, 也就是与物种利用环境资源的能力有关[35]。DCA排序图可以综合反应各物种空间分布的综合对比关系以及不同物种间的生态特性相似程度, 生态学特性相近的种在排序图上往往比较接近[36]。研究各种群之间资源利用和空间分布的相互关系, 应用DCA排序结合生态位特征分析的方法, 可以得到更为全面的研究结果[37]。本研究以重要值作为指标, 建立物种×样方矩阵, 利用Cannon 5.0软件进行不同生境条件下物种间的生态相似性DCA排序。
1.4 数据处理所有数据在Excel 2016和SPSS 21.0软件中进行统计、计算和制图, 采取单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)进行方差分析和多重比较。数量分类和排序的研究方法多应用于群落的类型划分, 也有不少学者将其应用于物种的归类和排序, 也取得了较好的效果, 研究结果同样可以反映物种的空间分布格局和生态适应性[37-38]。DCA排序不但包括了生态位方面的信息, 更多的综合了物种之间的相互关系[37]。因此, 本研究采用Canoco 5.0软件绘制不同土壤类型生境下各物种生态位重叠指数的种系统聚类排序图。
2 结果与分析 2.1 异质生境下群落特征 2.1.1 种类组成与数量特征实际调查结果显示(图 1), 研究样地内共有42个植物种, 分别属于15科, 36属。总体来看, 植物群落主要以豆科、禾本科、菊科和藜科为主, 4科植物在不同土壤类型生境下样地的数量占比之和分别为59.64%(风沙土)、61.96%(灰钙土)和76.32%(基岩风化沉积土)。从单科来看, 风沙土生境中豆科>菊科>藜科>禾本科>其他单科;灰钙土生境排序为豆科>禾本科>菊科>藜科>其他单科;基岩风化沉积土生境中则为豆科>菊科>禾本科>藜科>其他单科。从围封和放牧来看, 围封样地植物所属科数高于放牧样地。除风沙土生境中重度放牧会显著减少各科植物的种类外, 相同土壤类型生境间轻度和/或中度放牧没有造成样地中植物科数量的显著变化。

|
| 图 1 各样地植物所属科数量 Fig. 1 The number of species in each family in each plot |
研究区以多年生的植物为主, 其次是一年生和一年生至多年生植物(图 2)。其中, 风沙土生境中多年生植物平均为6.55种, 显著低于灰钙土(10.05种)和基岩风化沉积土生境(12.00种)。一年生植物中, 风沙土生境平均为3.59种, 显著高于基岩风化沉积土生境(2.8种)和灰钙土生境(1.70种)。而一年生至多年生植物的种数在风沙土生境(1.63种)、灰钙土生境(1.50种)和基岩风化沉积土生境(1.10种)中差异不大。从围封和放牧来看, 3种土壤类型放牧样地中多年生植物的种类(7.41种)少于围封样地(9.49种), 而增加了一年生植物的种类(3.05种)。

|
| 图 2 各样地植物功能类群 Fig. 2 The plant of annual, annual/perennial, perennials in each plot |
草本占有绝对优势, 物种数占比均高于85%, 其次为半灌木, 调查样方内无灌木和乔木出现(图 3)。在不同土壤类型生境下, 风沙土中草本植物平均有10.81种, 灰钙土中有11.75种, 均低于基岩风化沉积土生境中(14.50种);而风沙土中半灌木平均为0.97种, 低于灰钙土中的1.50种和基岩风化沉积土中的1.40种。从围封和放牧来看, 放牧样地中草本植物平均物种数(10.98种)低于围封样地(12.52种), 半灌木植物的物种数则差异不大。风沙土生境中, 随着放牧程度的加重, 草本植物的平均物种数显著降低, 而对半灌木物种数影响不大。

|
| 图 3 各样地植物生长型 Fig. 3 The plant growth form in each plot |
从表 1中可以看出, 不同生境下调查样地中植物群落数量特征有着显著的差异。总体来看, 在不同土壤类型下, 风沙土生境中的植物群落平均生物量最高(113.60 g/m2), 其次为灰钙土生境(101.52 g/m2), 基岩风化沉积土中最低, 平均值仅为61.70 g/m2。灰钙土中的植物群落平均盖度最高, 为52.63%, 其次是风沙土生境中平均盖度为46.54%, 基岩风化沉积土中最低, 仅为34.20%。风沙土生境中平均密度高达526.67株/m2, 基岩风化沉积土中为515.30株/m2, 灰钙土生境最低, 为388.10株/m2。植物群落中的平均高度在3种土壤类型生境下差异不显著, 大小排序为:基岩风化沉积土(129.43 cm)>灰钙土(118.41 cm)>风沙土(100.89 cm)。
| 土壤类型 Soil types |
围封/放牧 Enclosure/grazing |
生物量 Biomass/(g/m2) |
盖度 Coverage/% |
密度 Density/(株/m2) |
高度 Height/cm |
|
| 风沙土 | 围封 | 短期 | 128.51±46.92 | 60.20±14.66 | 570.90±347.74 | 133.87±36.18 |
| Aeolian sandy soil | 长期 | 141.60±41.09 | 48.60±6.09 | 304.60±137.18 | 126.17±35.77 | |
| 放牧 | 轻度 | 118.52±49.85 | 41.40±4.87 | 450.40±91.33 | 87.45±15.46 | |
| 中度 | 102.11±33.06 | 53.90±17.21 | 1021.93±335.10 | 87.19±25.18 | ||
| 重度 | 77.25±26.99 | 28.60±11.79 | 285.50±249.28 | 69.77±34.01 | ||
| 灰钙土 | 围封 | 短期 | 103.31±14.36 | 59.00±3.02 | 277.10±115.22 | 125.20±8.37 |
| Sierozem soil | 长期 | 125.24±44.27 | 67.90±7.99 | 255.60±66.21 | 173.07±43.38 | |
| 放牧 | 轻度 | 75.59±22.82 | 38.00±11.77 | 337.80±149.21 | 111.33±42.46 | |
| 中度 | 101.93±19.31 | 45.60±8.51 | 681.90±360.06 | 64.03±16.40 | ||
| 基岩风化沉积土 | 围封 | 短期 | 75.96±28.04 | 36.20±17.72 | 636.40±325.97 | 150.39±35.02 |
| Weathering bedrock | 放牧 | 中度 | 47.43±10.38 | 32.20±6.01 | 394.20±131.06 | 108.47±14.62 |
从围封和放牧来看, 3种土壤类型生境下放牧均会减少植物群落的平均生物量, 同时引起群落平均盖度和高度的降低。总体来看, 放牧生境下植株密度(528.62株/m2)高于围封生境(408.92株/m2), 在风沙土和灰钙土生境下, 中度放牧显著增加了群落的密度。但重度放牧则会降低样地内植株密度。在基岩风化沉积土生境中短期围封植株密度高于中度放牧条件, 但差异不显著。
2.1.2 多样性分析物种多样性可以体现生物与生物、生物与环境间的关系, 还可以体现生物资源的丰富性。研究区不同生境样地中平均丰富度的变化范围为8.40—16.20(表 2), 其中, 风沙土生境中丰富度的平均值为11.77, 灰钙土为13.25, 基岩风化沉积土为15.90。从围封和放牧来看, 围封样地平均丰富度(13.46)略高于放牧样地(12.73)。除风沙土生境中重度放牧会显著减少物种丰富度外, 相同土壤类型生境间轻度和/或中度放牧没有造成样地中物种丰富度的显著变化。这和野外调查所观察到的植物生长状况是一致的。
| 土壤类型 Soil types |
围封/放牧 Enclosure/Grazing |
丰富度(S) Richness |
多样性指数(H) Shannon-Wiener diversity index |
均匀度指数(E) Pielou evenness index |
|
| 风沙土 | 围封 | 短期 | 13.07±2.99 | 3.97±1.06 | 1.54±0.28 |
| Aeolian sandy soil | 长期 | 13.20±1.64 | 4.01±0.56 | 1.55±0.14 | |
| 放牧 | 轻度 | 13.80±2.17 | 4.16±0.83 | 1.58±0.24 | |
| 中度 | 10.40±2.59 | 3.08±0.86 | 1.31±0.24 | ||
| 重度 | 8.40±1.84 | 2.45±0.52 | 1.15±0.14 | ||
| 灰钙土 | 围封 | 短期 | 12.00±1.41 | 3.72±0.42 | 1.50±0.12 |
| Sierozem soil | 长期 | 12.80±2.17 | 3.94±0.75 | 1.54±0.20 | |
| 放牧 | 轻度 | 15.40±6.58 | 4.67±2.15 | 1.69±0.53 | |
| 中度 | 12.80±2.17 | 3.93±0.86 | 1.54±0.25 | ||
| 基岩风化沉积土 | 围封 | 短期 | 16.20±1.79 | 5.03±0.64 | 1.80±0.16 |
| Weathering bedrock | 放牧 | 中度 | 15.60±1.14 | 4.80±0.56 | 1.74±0.17 |
Shannon-Wiener多样性指数用来估算群落多样性的高低, 其变化范围为2.45—5.03, 其中, 风沙土生境多样性平均值为3.53, 灰钙土为4.07, 基岩风化沉积土为4.92。围封样地平均值(4.13)略高于放牧样地(3.85), 在风沙土生境中轻度放牧的多样性指数略高于围封样地, 但无显著差异, 而重度放牧显著低于围封样地, 且随着放牧程度的增加, 多样性指数呈降低的趋势。
Pielou均匀度指数用来描述物种中个体的相对丰富度或所占比例, 变化范围为1.15—1.80, 其中, 风沙土生境多样性平均值为1.43, 灰钙土为1.57, 基岩风化沉积土为1.77。从围封和放牧来看, 围封样地均匀度指数平均值(1.59)略高于放牧样地(1.50), 风沙土生境下随着放牧程度的增加, 均匀度指数也呈降低的趋势。而灰钙土生境下, 轻度放牧较之围封可以提高样地内植物群落的均匀度指数, 但无显著差异。
2.2 重要值从重要值总和来看(表 3), 排名前十位的分别是猪毛蒿、虫实、牛枝子、针茅、蒺藜、糙隐子草、苦豆子、阿尔泰狗娃花、砂珍棘豆、狗尾草, 而且猪毛蒿占有绝对优势。从不同土壤类型条件下, 风沙土生境中重要值最大的是猪毛蒿, 其次为虫实和蒺藜, 重要值最小的是球果荠(< 0.001);灰钙土生境中重要值最大的为猪毛蒿, 其次为牛枝子、针茅和赖草, 最小的是地锦;基岩风化沉积土中猪毛蒿最大, 其次为虫实、糙隐子草和草木犀状黄芪, 而重要值最小的是二色补血草和牻牛儿苗。这和野外调查所观察到的植物生长状况是一致的。
| 物种 Plant species |
风沙土 Aeolian sandy soil |
灰钙土 Sierozem soil |
基岩风化沉积土 Weathering bedrock |
总和 Total |
|||||||||||||
| 围封 Enclosure |
放牧 Grazing |
围封 Enclosure |
放牧 Grazing |
围封 Enclosure |
放牧 Grazing |
||||||||||||
| 短期 | 长期 | 轻度 | 中度 | 重度 | 短期 | 长期 | 轻度 | 中度 | 短期 | 中度 | |||||||
| 猪毛蒿 Artemisia scoparia |
0.484 | 0.386 | 0.495 | 0.459 | 0.094 | 0.230 | 0.315 | 0.36 | 0.487 | 0.262 | 0.195 | 3.769 | |||||
| 虫实 Corispermum mongolicum |
0.116 | 0.122 | 0.026 | 0.187 | 0.264 | — | 0.045 | 0.025 | 0.096 | 0.197 | 0.125 | 1.204 | |||||
| 牛枝子 Lespedeza potaninii |
0.101 | 0.152 | 0.138 | 0.058 | 0.007 | 0.136 | 0.268 | 0.134 | 0.100 | 0.045 | 0.019 | 1.158 | |||||
| 针茅 Stipa capillata |
0.020 | 0.017 | 0.013 | 0.027 | — | 0.288 | 0.143 | 0.092 | 0.029 | — | 0.005 | 0.634 | |||||
| 蒺藜 Tribulus terrester |
0.036 | 0.079 | 0.015 | 0.046 | 0.309 | 0.005 | — | 0.036 | 0.033 | — | — | 0.560 | |||||
| 糙隐子草 Cleistogenes squarrosa |
0.068 | — | 0.025 | 0.008 | — | 0.079 | 0.014 | 0.064 | 0.042 | 0.083 | 0.177 | 0.560 | |||||
| 苦豆子 Sophora alopecuroides |
0.063 | 0.105 | 0.047 | 0.118 | 0.016 | 0.038 | 0.033 | 0.032 | 0.033 | 0.029 | 0.03 | 0.546 | |||||
| 阿尔泰狗娃花 Heteropappus altaicus |
0.027 | 0.032 | 0.084 | 0.025 | — | 0.048 | 0.068 | 0.021 | 0.099 | 0.032 | 0.041 | 0.477 | |||||
| 砂珍棘豆 Oxytropis racemosa |
0.028 | 0.023 | 0.045 | 0.044 | 0.007 | 0.041 | 0.018 | 0.04 | 0.048 | 0.033 | 0.090 | 0.417 | |||||
| 狗尾草 Setaria viridis |
0.032 | — | 0.005 | 0.081 | 0.211 | — | 0.027 | 0.019 | 0.022 | 0.012 | — | 0.410 | |||||
| 白草 Pennisetum centrasiaticum |
0.040 | 0.064 | 0.033 | 0.030 | 0.081 | 0.019 | 0.021 | 0.013 | 0.015 | 0.021 | 0.057 | 0.395 | |||||
| 赖草 Leymus secalinus |
0.048 | — | — | 0.004 | — | 0.1 | 0.074 | 0.067 | — | 0.048 | 0.044 | 0.384 | |||||
| 草木犀状黄芪 Astragalus melilotoides |
0.024 | 0.037 | — | 0.030 | — | — | 0.06 | 0.023 | — | 0.109 | 0.072 | 0.357 | |||||
| 老瓜头 Cynanchum komarovii |
0.006 | 0.012 | — | 0.052 | 0.044 | 0.005 | 0.032 | 0.017 | 0.058 | — | 0.022 | 0.248 | |||||
| 猪毛菜 Salsola collina |
0.044 | 0.014 | 0.028 | 0.023 | — | — | 0.025 | 0.030 | 0.026 | 0.034 | 0.022 | 0.244 | |||||
| 披针叶黄华 Thermopsis lanceolata |
0.011 | — | — | 0.031 | — | 0.017 | 0.007 | 0.062 | — | 0.037 | 0.069 | 0.234 | |||||
| 米口袋 Gueldenstaedtia verna |
0.022 | — | 0.025 | 0.021 | — | 0.039 | 0.016 | 0.034 | 0.051 | 0.014 | 0.010 | 0.233 | |||||
| 丝叶山苦荬 Ixeris chinense |
0.010 | 0.029 | 0.02 | 0.014 | — | 0.02 | 0.031 | 0.016 | 0.026 | 0.03 | 0.026 | 0.222 | |||||
| 菟丝子 Cuscuta chinensis |
0.029 | 0.03 | 0.036 | 0.032 | 0.019 | — | — | 0.017 | 0.01 | 0.013 | — | 0.187 | |||||
| 大戟 Euphorbia pekinensis |
0.013 | 0.02 | 0.023 | 0.022 | — | 0.006 | 0.015 | 0.021 | 0.024 | 0.024 | 0.012 | 0.179 | |||||
| 雾冰藜 Bassia dasyphylla |
0.010 | 0.01 | 0.063 | 0.025 | 0.024 | — | 0.021 | — | 0.02 | — | — | 0.173 | |||||
| 远志 Polygala tenuifolia |
0.021 | 0.023 | 0.005 | 0.024 | — | 0.026 | 0.024 | 0.014 | 0.028 | — | — | 0.164 | |||||
| 冬青叶兔唇花 Lagochilus ilicifolius |
0.057 | — | — | — | — | — | 0.061 | — | 0.042 | — | — | 0.160 | |||||
| 角蒿 Incarvillea sinensis |
0.028 | — | 0.023 | — | — | — | 0.038 | — | — | 0.021 | — | 0.110 | |||||
| 二色补血草 Limonium bicolor |
— | 0.051 | — | — | — | — | 0.052 | — | — | 0.004 | — | 0.107 | |||||
| 二裂委陵菜 Potentilla bifurca |
— | — | — | 0.013 | — | — | 0.072 | — | 0.017 | — | — | 0.102 | |||||
| 田旋花 Convolvulus arvensis |
0.011 | 0.005 | — | 0.015 | — | — | 0.015 | 0.047 | — | — | — | 0.093 | |||||
| 叉枝鸦葱 Scorzonera divaricata |
0.005 | 0.042 | — | 0.014 | — | — | — | 0.010 | — | — | 0.015 | 0.087 | |||||
| 灰绿藜 Chenopodium glaucum |
0.014 | — | 0.009 | — | 0.015 | — | 0.045 | — | — | — | — | 0.084 | |||||
| 地锦 Euphorbia humifusa |
0.022 | — | 0.007 | 0.019 | 0.009 | 0.005 | — | 0.005 | — | 0.011 | 0.005 | 0.082 | |||||
| 银灰旋花 Conuoluulus ammannii |
0.006 | — | — | 0.011 | — | 0.013 | — | 0.032 | — | — | — | 0.062 | |||||
| 画眉草 Eragrostis pilosa |
— | — | — | — | 0.041 | — | — | — | — | — | 0.014 | 0.054 | |||||
| 砂蓝刺头 Echinops gmelini |
— | — | — | 0.019 | — | — | — | 0.019 | — | 0.015 | — | 0.053 | |||||
| 乳苣 Mulgedium tataricum |
— | 0.045 | — | — | — | — | — | — | — | — | — | 0.045 | |||||
| 脓疮草 Panzeria alaschanica |
— | — | — | — | — | — | 0.019 | — | 0.021 | — | — | 0.040 | |||||
| 骆驼蓬 Peganum harmala |
0.029 | — | — | — | — | 0.01 | — | — | — | — | — | 0.039 | |||||
| 地梢瓜 Cynanchum thesioides |
0.021 | — | — | — | — | — | — | — | — | 0.017 | — | 0.038 | |||||
| 猫头刺 Oxytropis aciphylla |
— | — | — | — | — | — | 0.014 | — | 0.014 | — | — | 0.029 | |||||
| 沙蓬 Agriophyllum squarrosum |
0.011 | — | — | — | — | — | — | — | — | — | — | 0.011 | |||||
| 火烙草 Echinops przewalskii |
— | — | — | — | — | — | — | — | — | 0.005 | — | 0.005 | |||||
| 牻牛儿苗 Erodium stephanianum |
— | — | — | — | — | — | — | — | — | 0.004 | — | 0.004 | |||||
| 球果荠 Neslia paniculata |
< 0.001 | — | — | — | — | — | — | — | — | — | — | < 0.001 | |||||
| “—”表示此物种在该样地中未出现 | |||||||||||||||||
综合围封和放牧条件来看, 研究区内围封条件下主要优势种集中在猪毛蒿、牛枝子、虫实和针茅等植物之间, 物种的生活型以一年至多年生和多年生草本或半灌木为主。而放牧条件下优势种为猪毛蒿、虫实、蒺藜、牛枝子和狗尾草, 生活型是一年生草本的植物比重有所增加。这和刘小丹等[39]的研究结果一致。
总体来看, 研究区内猪毛蒿是主要优势种, 但在不同土壤类型生境下, 其重要值不同, 风沙土生境下最大, 其次为灰钙土, 基岩风化沉积土生境中最小。但由于该植物不是优良牧草, 其占据优势势必导致草场质量下降。而牛枝子和针茅则表现为灰钙土生境最大, 风沙土和基岩风化沉积土次之。放牧会导致荒漠草原植物群落中多年生草本和半灌木重要值比重的下降, 而增加一年生草本的重要值。
2.3 生态位宽度生态位宽度和生态位重叠值是解释植物对资源环境的利用状况和对资源竞争情况最好的指标[8, 40]。生态位宽度是生物利用资源多样性的一个指标, 在一定程度上表现了物种对环境的适应性和利用资源的能力。物种生态位越宽, 利用资源的能力越强, 分布更为广泛。
不同生境草地种群生态位总宽度排名前10位的分别为:猪毛蒿>牛枝子>砂珍棘豆>虫实>苦豆子>丝叶山苦荬>糙隐子草>白草>阿尔泰狗娃花>猪毛菜, 生态位总宽度接近或大于10.00(表 4)。这些种在研究区广泛分布, 是典型的泛化种。不同土壤类型条件下, 风沙土生境中生态位宽度最大的是猪毛蒿, 其次是猪毛菜和虫实;灰钙土生境中重要值最大的为猪毛蒿, 其次为牛枝子、米口袋和针茅;基岩风化沉积土中猪毛蒿最大, 其次为赖草、阿尔泰狗娃花、丝叶山苦荬、虫实和老瓜头。同一种群不同土壤类型条件下的生态位宽度也不同, 分布较广的猪毛蒿种群生态位宽度值在灰钙土生境下最高(4.74), 风沙土(4.58)和基岩风化沉积土(4.50)次之。多数一年生植物(如虫实、猪毛菜、狗尾草、地锦等)在风沙土生境下生态位宽度最高, 而多年生植物(如针茅、白草、远志、米口袋等)在灰钙土生境下往往生态位宽度值相对较高。
| 物种 Plant species |
风沙土 Aeolian sandy soil |
灰钙土 Sierozem soil |
基岩风化沉积土 Weathering bedrock |
生态位总宽度 Total niche breadth |
|||||||||||||
| 围封 Enclosure |
放牧 Grazing |
围封 Enclosure |
放牧 Grazing |
围封 Enclosure |
放牧 Grazing |
||||||||||||
| 短期 | 长期 | 轻度 | 中度 | 重度 | 短期 | 长期 | 轻度 | 中度 | 短期 | 中度 | |||||||
| 猪毛蒿 Artemisia scoparia |
4.81 | 4.68 | 4.96 | 4.92 | 3.56 | 4.85 | 4.48 | 4.79 | 4.84 | 4.18 | 4.81 | 15.40 | |||||
| 牛枝子 Lespedeza potaninii |
4.07 | 3.53 | 3.11 | 3.09 | 2.00 | 4.96 | 4.44 | 2.88 | 4.53 | 4.40 | 3.37 | 12.49 | |||||
| 砂珍棘豆 Oxytropis racemosa |
2.55 | 3.48 | 4.34 | 2.90 | 1.00 | 4.42 | 1.00 | 4.49 | 4.74 | 3.89 | 2.69 | 11.51 | |||||
| 虫实 Corispermum mongolicum |
3.65 | 2.95 | 1.78 | 4.64 | 4.21 | — | 2.83 | 1.77 | 2.96 | 4.65 | 3.96 | 11.03 | |||||
| 苦豆子 Sophora alopecuroides |
3.68 | 1.30 | 2.91 | 3.78 | 3.14 | 2.96 | 1.98 | 3.57 | 3.44 | 2.61 | 3.89 | 10.34 | |||||
| 丝叶山苦荬 Ixeris chinense |
1.56 | 3.23 | 2.81 | 1.21 | 1.00 | 4.29 | 4.47 | 1.98 | 1.00 | 3.77 | 4.97 | 10.27 | |||||
| 糙隐子草 Cleistogenes squarrosa |
3.43 | — | 4.66 | 1.00 | — | 3.93 | 1.00 | 3.08 | 2.77 | 3.69 | 4.73 | 10.20 | |||||
| 白草 Pennisetum centrasiaticum |
3.41 | 2.72 | 4.84 | 2.74 | 1.58 | 1.76 | 1.90 | 2.26 | 3.06 | 2.75 | 4.51 | 10.08 | |||||
| 阿尔泰狗娃花 Heteropappus altaicus |
2.28 | 3.80 | 3.12 | 2.00 | — | 3.54 | 2.63 | 1.67 | 2.46 | 4.68 | 4.09 | 10.02 | |||||
| 猪毛菜 Salsola collina |
3.41 | 2.87 | 4.57 | 3.34 | — | — | 1.00 | 2.94 | 1.00 | 4.81 | 3.73 | 9.98 | |||||
| 米口袋 Gueldenstaedtia verna |
2.12 | — | 3.74 | 2.39 | — | 4.04 | 2.79 | 4.24 | 3.84 | 1.74 | 1.00 | 9.22 | |||||
| 针茅 Stipa capillata |
2.85 | 3.93 | 2.29 | 1.00 | — | 4.88 | 1.68 | 3.93 | 2.83 | — | 1.00 | 8.99 | |||||
| 赖草 Leymus secalinus |
1.79 | — | — | 1.00 | 1.00 | 2.23 | 1.86 | 4.08 | — | 4.68 | 4.24 | 8.38 | |||||
| 披针叶黄华 Thermopsis lanceolata |
2.92 | — | — | 1.35 | 2.88 | 1.74 | 1.00 | 2.44 | — | 4.40 | 3.92 | 7.96 | |||||
| 菟丝子 Cuscuta chinensis |
3.51 | 4.85 | 1.87 | 3.02 | 1.00 | — | — | 1.83 | 1.28 | 2.87 | — | 7.92 | |||||
| 蒺藜 Tribulus terrester |
2.85 | 1.98 | 3.00 | 2.34 | 4.69 | 2.00 | — | 1.67 | 2.29 | — | — | 7.78 | |||||
| 狗尾草 Setaria viridis |
2.70 | — | 2.00 | 2.54 | 4.53 | — | 2.58 | 2.88 | 1.00 | 1.73 | — | 7.57 | |||||
| 远志 Polygala tenuifolia |
2.19 | 1.85 | 1.00 | 1.00 | 1.00 | 3.43 | 4.36 | 1.97 | 2.95 | — | — | 7.39 | |||||
| 大戟 Euphorbia pekinensis |
1.65 | 2.46 | 1.00 | 1.30 | 3.25 | 1.00 | 1.00 | 1.97 | 1.00 | 3.63 | 3.41 | 7.33 | |||||
| 草木犀状黄芪 Astragalus melilotoides |
1.00 | 3.23 | — | 1.99 | — | — | 1.00 | 1.97 | — | 3.14 | 4.13 | 6.87 | |||||
| 老瓜头 Cynanchum komarovii |
1.00 | 1.00 | — | 1.00 | 2.87 | 1.00 | 2.96 | 1.00 | 1.00 | — | 4.28 | 6.42 | |||||
| 地锦 Euphorbia humifusa |
2.78 | — | 1.83 | 3.12 | 2.94 | 1.00 | — | 1.98 | — | 1.00 | 1.00 | 6.03 | |||||
| 叉枝鸦葱 Scorzonera divaricata |
1.00 | 4.21 | — | 1.41 | — | — | — | 1.00 | — | — | 2.59 | 5.33 | |||||
| 雾冰藜 Bassia dasyphylla |
1.14 | 1.00 | 1.25 | 1.47 | 2.42 | — | 1.99 | — | 1.67 | — | — | 4.32 | |||||
| 角蒿 Incarvillea sinensis |
1.48 | — | 2.97 | — | — | — | 1.00 | — | — | 1.00 | — | 3.61 | |||||
| 田旋花 Convolvulus arvensis |
2.36 | 1.00 | — | 1.00 | — | — | 1.55 | 1.00 | — | — | — | 3.31 | |||||
| 灰绿藜 Chenopodium glaucum |
1.00 | — | 1.55 | — | 1.87 | — | 1.89 | — | — | — | — | 3.24 | |||||
| 猫头刺 Oxytropis aciphylla |
— | — | — | — | — | — | 1.00 | — | 2.91 | — | — | 3.08 | |||||
| 地梢瓜 Cynanchum thesioides |
2.72 | — | — | — | — | — | — | — | — | 1.00 | — | 2.90 | |||||
| 银灰旋花 Conuoluulus ammannii |
1.00 | — | — | 1.00 | — | 1.00 | — | 1.88 | — | — | — | 2.55 | |||||
| 脓疮草 Panzeria alaschanica |
— | — | — | — | — | — | 1.89 | — | 1.71 | — | — | 2.55 | |||||
| 二裂委陵菜 Potentilla bifurca |
— | — | — | 1.00 | — | — | 1.00 | — | 1.96 | — | — | 2.42 | |||||
| 冬青叶兔唇花 Lagochilus ilicifolius |
1.00 | — | — | — | — | — | 1.86 | — | 1.00 | — | — | 2.33 | |||||
| 砂蓝刺头 Echinops gmelini |
— | — | — | 1.79 | — | — | — | 1.00 | — | 1.00 | — | 2.28 | |||||
| 二色补血草 Limonium bicolor |
— | 1.00 | — | — | — | — | 1.00 | — | — | 1.00 | — | 1.73 | |||||
| 骆驼蓬 Peganum harmala |
1.35 | — | — | — | — | 1.00 | — | — | — | — | — | 1.68 | |||||
| 沙蓬 Agriophyllum squarrosum |
1.31 | — | — | — | 1.00 | — | — | — | — | — | — | 1.65 | |||||
| 画眉草 Eragrostis pilosa |
— | — | — | — | 1.00 | — | — | — | — | — | 1.00 | 1.41 | |||||
| 牻牛儿苗 Erodium stephanianum |
— | — | — | — | — | — | — | — | — | 1.00 | — | 1.00 | |||||
| 火烙草 Echinops przewalskii |
— | — | — | — | — | — | — | — | — | 1.00 | — | 1.00 | |||||
| 乳苣 Mulgedium tataricum |
— | 1.00 | — | — | — | — | — | — | — | — | — | 1.00 | |||||
| 球果荠 Neslia paniculata |
< 0.01 | — | — | — | — | — | — | — | — | — | — | < 0.01 | |||||
综合围封和放牧条件来看, 围封条件下生态位宽度排名靠前的分别是猪毛蒿、牛枝子、菟丝子、虫实、丝叶山苦荬和阿尔泰狗娃花等植物, 而放牧条件下生态位宽度较大的为猪毛蒿、苦豆子、砂珍棘豆、糙隐子草、虫实和白草。放牧对猪毛蒿种群生态位宽度的影响较小, 但对其他物种, 特别是一年生草本(如菟丝子、丝叶山苦荬和阿尔泰狗娃花)生态位宽度的影响较大, 寄生草本菟丝子生态位宽度由围封样地的3.74急剧下降到放牧样地的1.80。而长期围封显著降低了灰钙土生境下的多年生草本针茅的生态位宽度, 但在风沙土生境下则相对提高。
总体来看, 在不同生境条件下猪毛蒿的生态位宽度均为最大, 说明这种植物是研究区的主要优势种, 环境适应性强, 占据大部分可利用资源, 中度放牧生境较围封条件下猪毛蒿生态位宽度值大, 但重度放牧会降低其宽度。而牻牛儿苗、火烙草和乳苣的生态位宽度较小, 均为0.100, 球果荠的生态位宽度实际值很小(< 0.01), 说明这些植物生态位宽度较窄, 具有选择性利用资源的特点, 在资源竞争中处于劣势, 也就是说此类植物对研究区环境的要求较高, 资源利用谱低, 对环境的适应能力较弱。这几种植物均为偶见种, 其生态位宽度与实际调查中观察到的现象一致。
从调查样地植物群落中各植物生态位宽度和重要值的相关关系来看, 在3种土壤类型生境下, 两者均呈正相关关系, 但解释变量R2均较低, 也就是说生态位宽度和重要值呈弱相关关系。各物种生态位总宽度和重要值间呈极显著正相关, 解释变量相对较高(图 4)。一般来说, 重要值越大的物种, 往往生态位宽度也越大, 对资源的利用和对环境的适应能力也越强[41], 但也有研究发现重要值大小并非是生态位宽度大小的唯一因素, 即重要值最大的物种其生态位宽度并非最大[8], 说明群落中物种生态位宽度与其重要值不一定呈正相关关系。本研究中虽然呈现出正相关关系, 但解释变量较低, 这可能和调查样地中不同立地的土壤理化性质存在差异叠加上人类活动干扰, 所形成的复杂异质性生境有关。

|
| 图 4 荒漠草原不同生境物种生态位宽度、总宽度与重要值相关关系 Fig. 4 Correlation between niche breadth and IV in desert steppe with heterogeneous habitat and the relationship of total niche breadth and IV |
在分析植物生态位宽度的基础上进一步分析种间生态位重叠, 可以更精确刻画群落的动态、物种组成及物种的优势度等特征[42]。本研究中, 相同植物种间在不同生境下的生态位重叠指数是不同的。猪毛蒿种群在放牧和短期围栏内与其他物种的生态位重叠指数均较高, 原因主要是猪毛蒿植株本身的繁殖特性、对胁迫条件的耐受性以及对环境资源的普适性特征造成其分布范围较广, 进而造成各生境条件下均能存活。而且总体来看, 中度放牧生境较围封条件下猪毛蒿生态位重叠值大。但在长期围封样地中, 特别是灰钙土生境下, 猪毛蒿与其他物种间的重叠指数比重有所降低, 而牛枝子、草木犀状黄芪、针茅等多年生的植物与伴生种的生态位重叠指数比重则明显提高。可以看出不同生境条件下生态位重叠值最大的植物, 其生态需求较为相似, 目前有着较好的共生关系, 但在资源利用上存在竞争关系。而冬青叶兔唇花与大多数共生种的生态重叠指数普遍偏低, 说明冬青叶兔唇花和其他物种对环境资源的需求存在差异。以上结果和野外调查所观察到的植物分布生长状况基本一致。
研究区具有高生态位宽度物种之间具有高的生态位重叠, 但在某些生境条件下, 生态位较窄的物种也会出现较大的生态位重叠, 有部分植物种间(如雾冰藜和老瓜头、草木犀状黄芪和冬青叶兔唇花、老瓜头和二色补血草)的生态位重叠值为1.00, 接近完全重叠。出现上述情况可能是植物种群为了在特殊的生境条件下获得生存, 对有限的环境资源(水分、养分等)展开竞争的结果。事实上, 2个物种之间对资源空间的生态位占有只是无限接近, 故重叠也只是无限接近1.00[39]。这也说明生态位宽度与生态位重叠之间并不存在直接的线性关系, 这是由物种可利用的环境资源空间分布异质性造成的。
2.4.2 DCA排序分析将不同土壤类型生境下各物种生态位重叠指数的种系统聚类排序图(图 5), 与各物种的重要值和生态位宽度值对比可知, 分布于DCA排序图周边的多为重要值较小和生态位宽度较低的种, 如牻牛儿苗(S39)、二色补血草(S37)、角蒿(S29)、地梢瓜(S9)等。而重要值较大和生态位较宽的猪毛蒿(S1)、虫实(S2)、猪毛菜(S3)、牛枝子(S14)和针茅(S35)等则分别位于或靠近各生境排序图的中心。从综合分析排序图(图 5)也可以得出相同的规律。说明越靠近排序图中心的物种, 在群落中往往占有重要的地位。

|
| 图 5 生态位重叠指数的种系统聚类 Fig. 5 Dendrogram of system clustering using niche overlap indices S1:猪毛蒿Artemisia scoparia;S2:虫实Corispermum mongolicum;S3:猪毛菜Salsola collina;S4:蒺藜Tribulus terrester;S5:白草Pennisetum centrasiaticum;S6:田旋花Convolvulus arvensis;S7:菟丝子Cuscuta chinensis;S8:沙蓬Agriophyllum squarrosum;S9:地梢瓜Cynanchum thesioides;S10:狗尾草Setaria viridis;S11:丝叶山苦荬Ixeris chinense;S12:砂珍棘豆Oxytropis racemosa;S13:地锦Euphorbia humifusa;S14:牛枝子Lespedeza potaninii;S15:苦豆子Sophora alopecuroides;S16:雾冰藜Bassia dasyphylla;S17:草木犀状黄芪Astragalus melilotoides;S18:远志Polygala tenuifolia;S19:米口袋Gueldenstaedtia verna;S20:砂蓝刺头Echinops gmelini;S21:二裂委陵菜Potentilla bifurca;S22:披针叶黄华Thermopsis lanceolata;S23:银灰旋花Conuoluulus ammannii;S24:糙隐子草Cleistogenes squarrosa;S25:赖草Leymus secalinus;S26:大戟Euphorbia pekinensis;S27:老瓜头Cynanchum komarovii;S28:骆驼蓬Peganum harmala;S29:角蒿Incarvillea sinensis;S30:灰绿藜Chenopodium glaucum;S31:阿尔泰狗娃花Heteropappus altaicus;S32:脓疮草Panzeria alaschanica;S33:球果荠Neslia paniculata;S34:叉枝鸦葱Scorzonera divaricata;S35:针茅Stipa capillata;S36:冬青叶兔唇花Lagochilus ilicifolius;S37:二色补血草Limonium bicolor;S38:猫头刺Oxytropis aciphylla;S39:牻牛儿苗Erodium stephanianum;S40:火烙草Echinops przewalskii;S41:乳苣Mulgedium tataricum;S42:画眉草Eragrostis pilosa |
有研究发现, DCA排序图可以综合反映各物种空间分布的综合对比关系, 生态学特性相近的种在排序图上往往比较接近[37]。本研究中同样发现有较多的物种对在排序图中比较接近。这与种群生态位除受外部环境因子的影响外, 还受植物自身特性的制约[43]。进一步分析发现这些聚集的物种对在不同土壤类型生境下的生态位重叠值较大、生理生态学特性比较接近, 如牛枝子和骆驼蓬、地梢瓜和角蒿等。
2.4.3 生态位重叠指数总平均值不同生境草地群落植物种群生态位重叠指数的总平均值具有明显差异(图 6)。从不同土壤类型条件来看, 风沙土生境中生态位重叠指数总平均值0.43, 灰钙土生境中生态位重叠指数总平均值为0.48, 基岩风化沉积土生境中为0.58。说明基岩风化沉积土中各种群物种间的重叠程度最大, 而风沙土中物种间的重叠程度最小。

|
| 图 6 荒漠草原异质生境植物种群生态位重叠指数总平均值 Fig. 6 Mean niche overlap index of plant populations in desert steppe with heterogeneous habitat |
从围封和放牧条件来看, 风沙土和灰钙土生境下轻度放牧增加了各生境下物种间的生态位重叠指数, 基岩风化沉积土生境下中度放牧较短期放牧显著增加了生态位重叠指数。但在风沙土生境下, 随着放牧程度的增加, 生态位重叠指数呈减小的趋势。
3 结论与讨论生态位理论能较好地解释种群的环境适应性和资源利用能力[44-45], 已在研究种间关系、群落结构、群落演替、生物多样性、物种共存及进化等方面得到了广泛的应用和较好的解释[42, 46-48], 并得到了不断丰富和发展[49]。
3.1 异质生境下群落组成与生态位特征自2003年以来, 宁夏全面实施围封禁牧, 给草场提供了一个休养生息的机会, 大部分草地生态环境得到了好转。但多数研究表明, 长期围封禁牧会导致草地多样性减少, 而适度的干扰可提高物种丰富度[50]。也有研究发现, 封育措施使得植被数量值明显高于未封育区, 有利于沙化草地植被恢复与群落稳定, 但随着封育年限延长, 枯枝落叶等物质在地表不断增加以及较厚的生物结皮形成将导致植物生长受到抑制, 也就是说长期的竞争排除等生态过程作用使得种群之间演替产生了某种程度的生态位分离, 群落结构特征出现波动[39]。本研究发现, 丰富度、多样性和均匀度指数均呈现出围封样地高于放牧样地, 说明围封措施使荒漠草原植物群落组成多样性更丰富。导致研究结果不同的原因可能与本研究所调查的试验样地与其他研究区域气候条件、植被类型、土壤类型等生境差异较大有关。如有学者对典型草原围封内外样地研究发现, 猪毛蒿等优势种的生态位宽度排位靠前, 说明其在群落中分布广泛, 对资源的利用能力较强[20]。本研究对荒漠草原的研究结果表明, 猪毛蒿种群与其他物种的生态位重叠指数均较高, 但在长期围封样地, 特别是灰钙土生境下, 猪毛蒿与其他物种间的重叠指数比重有所降低, 而其他多年生的植物(如牛枝子、草木犀状黄芪、针茅等)与伴生种间的生态位重叠指数则显著提高, 且不同物种生态位宽度差异有减小的趋势, 说明长期围封后, 以上几种多年生植物在灰钙土生境下能均衡地利用环境资源, 实现协同生长。
同一物种在不同生境中生态位宽度会存在差异, 如牛枝子、砂珍棘豆、虫实、苦豆子等, 在野外调查中, 也观察到这些物种在某一生境下是优势种, 但在另一个生境中只是伴生种, 这主要是生境条件差异导致同一物种表现出不同的生态位。有学者发现土壤因子将会改变荒漠植物的分布格局[51], 本研究也发现不同土壤类型下的物种组成是不一样的, 除了种类和数量不同之外, 其优势物种也是不同的。风沙土具有降水入渗快、渗漏快的特点, 发育了能够适应土壤水分冲击式波动且种群数量或生物量在降雨特别丰富的时期暴发式增长的猪毛蒿、猪毛菜、绵蓬等植物[18], 而灰钙土内部紧实、黏粉粒含量较高导致降水入渗少, 但能保持水分, 针茅、牛枝子多生长于此。说明荒漠草原原始生境的破碎化或者退化会显著改变群落物种共存格局, 降低了物种对资源利用的相似性[18]。这和黄至欢等[52]研究发现喀斯特地区石灰岩土壤和红壤间断分布是影响该地区植物群落内物种组成的重要因素之一的结果相一致。因此, 建议针对不同的立地条件, 必须采取不同的人为措施, 以促进或抑制某一目标植物的生长[39]。当然, 影响生态位的因素较多, 物种的生态习性也是重要原因[34]。针茅对生境条件的要求较高, 资源范围变窄, 多生长于坡顶向阳、硬质灰钙土或裸露基岩生境中, 且往往成为优势种, 而风沙土, 特别是流动性强的生境不适宜针茅生存, 使其生态位宽度骤然减少, 甚至消失。此外, 资源本身的有效性及可利用范围往往限制植物种群的生态位扩展, 导致其生态位宽度处于较窄的范围内[53]。本研究不同土壤类型中, 风化基岩沉积土和灰钙土的硬度较大, 导致根系延伸受到了一定的限制, 使得其上生长的植物生态位宽度有所降低。
3.2 荒漠草原优势种群生态位特征一般种群生态位宽度的大小与它们对环境的适应能力呈正相关。种群生态位越大, 表明占用生态位空间大, 对各种环境资源的利用越充分, 分布范围越广, 往往发展成为群落中的优势群体。反之则说明该种的特化程度较高, 可能分布在局限的区域内。本研究中猪毛蒿、牛枝子、砂珍棘豆和虫实等物种较宽的生态位及资源利用谱决定了它们在该地区的优势地位, 由于其往往通过表型可塑性保护个体, 拓展领域范围, 以提高对环境资源的利用程度和自身的抗逆性[54], 具有较强的资源竞争能力, 因此在荒漠草原地区作为植物群落的泛化种出现。这和吴会峰等[45]在黄土丘陵区研究的结果相一致。而球果荠、乳苣、火烙草、牻牛儿苗等物种生态幅小的特化种, 其在样方中出现频率低, 说明这些植物具有较弱的资源竞争能力, 因此生态位宽度与优势度均处于较低水平。
当然, 生态位宽度与物种本身的生理生态适应性、利用资源数量等因素有关[55]。猪毛蒿为菊科蒿属植物, 抗干旱、抗风沙能力较强, 具有类似CAM植物的叶片形态、根蘖性强、繁殖可塑性强的生物生态复合特征, 且猪毛蒿是典型的r对策物种, 以极强的适应特性能够成功地侵入和定居在不同生境中, 如细小的种子能滞留在土壤中, 不致于随风滚动丢失, 种子萌发后胚根能以极快的速度生长, 使苗体牢牢固着于土壤中抵御风蚀的侵害。其对裸地利用能力也很强, 逢雨迅速生长, 具有较高的扩散能力, 且在沙化生境中占据优势种地位, 加之其具有强烈的化感作用[56], 生态位宽度值较大, 往往成为该地区的优势建群物种[57], 在自然演替过程中成为先锋物种[54], 很大程度抑制了其他植物的生长, 不利于物种共存, 使得种间生态位重叠降低[45], 导致群落生物多样性低。
4 展望生态学的核心是群落生态学, 而群落生态学的核心是物种共存, 但对物种共存的机制目前尚未有统一的结论。中性理论与生态位理论在群落构建中的作用一直存在争议, 近年来, 倾向于将生态位理论与中性理论整合, 认为两者对群落的构建都有作用, 这主要是其相对贡献与研究尺度和生态系统类型都有关系[58]。而真正的生态学突破是将中性理论和生态位理论的关键要素结合起来解释群落多样性模式[59]。如何将当代物种共存理论和生态位理论与基于过程的群落构建框架有机地结合在一起, 从而揭示群落构建和生物多样性维持的内在机制, 是未来研究工作的一个重要发展方向[60]。
由于环境的时空异质性和多维复合作用, 根据野外实地的测定难以把握单一因子的梯度范围和分离诸变量的作用。局域植物群落组成既取决于环境因素的作用(如气候、土壤和地形条件以及干扰), 同时又受制于群落内生物间的相互作用[61]。群落内植物间的相互作用使得过于相似的物种间发生竞争排斥, 从而使各物种间的相似性受限[62], 进而导致了同一群落内物种特征的趋异性[63]。荒漠草原生境条件复杂多变, 物种的竞争能力不只由资源利用能力所决定, 一些研究也证实物种的分布还受生活史、繁殖策略、土壤状况及其他物种干扰等多种因素的影响, 而且在异质生境下不同的物种对间也可能存在他感作用, 在植物分布、数量等方面起了较大作用, 因此今后研究群落内物种间的相互作用关系也很有必要。而荒漠草原优势物种猪毛蒿的化感作用是否对其他伴生种具有抑制或者促进作用, 尚不清楚, 值得今后开展进一步研究。
此外, 荒漠草原影响群落物种共存的另一个关键因子是水分, 在研究异质生境下植物群落组成及其种群生态位特征, 应耦合长期的水分连续监测数据, 这对于揭示影响该区域植物群落构建机制具有重要意义。因此, 荒漠草原异质生境条件下物种间的共存机制还有待于进一步探讨。
| [1] |
霍举颂, 刘卫国, 刘建国, 李宏侠, 徐悦, 玛丽娅·奴尔兰. 影响阜康荒漠-绿洲过渡带荒漠植物数量特征的土壤驱动力分析. 生态学报, 2017, 37(24): 8304-8313. |
| [2] |
Loreau M, Naeem S, Inchausti P, Bengtsson J, Grime J P, Hector A, Hooper D U, Huston M A, Raffaelli D, Schmid B, Tilman D, Wardle D A. Biodiversity and ecosystem functioning:current knowledge and future challenges. Science, 2001, 294(5543): 804-808. DOI:10.1126/science.1064088 |
| [3] |
Nathan R. Long-distance dispersal of plants. Science, 2006, 313(5788): 786-788. DOI:10.1126/science.1124975 |
| [4] |
薛建辉. 森林生态学. 北京: 中国林业出版社, 2006.
|
| [5] |
高杨.土壤营养异质性对鹅绒委陵菜种群分布格局和空间拓展的影响[D].长春: 东北师范大学, 2014. http://cdmd.cnki.com.cn/Article/CDMD-10200-1014372674.htm
|
| [6] |
徐远杰, 林敦梅, 石明, 谢妍洁, 王逸之, 管振华, 向建英. 云南哀牢山常绿阔叶林的空间分异及其影响因素. 生物多样性, 2017, 25(1): 23-33. |
| [7] |
Lundholm J T. Plant species diversity and environmental heterogeneity:spatial scale and competing hypotheses. Journal of Vegetation Science, 2009, 20(3): 377-391. DOI:10.1111/j.1654-1103.2009.05577.x |
| [8] |
字洪标, 阿的鲁骥, 刘敏, 胡雷, 陈焱, 杨有芳, 王长庭. 高寒草甸不同类型草地群落特征及优势种植物生态位差异. 应用与环境生物学报, 2016, 22(4): 546-554. |
| [9] |
Bergholz K, May F, Giladi I, Ristow M, Ziv Y, Jeltsch F. Environmental heterogeneity drives fine-scale species assembly and functional diversity of annual plants in a semi-arid environment. Perspectives in Plant Ecology, Evolution and Systematics, 2017, 24: 138-146. DOI:10.1016/j.ppees.2017.01.001 |
| [10] |
Clark D B, Palmer M W, Clark D A. Edaphic factors and the landscape-scale distributions of tropical rain forest trees. Ecology, 1999, 80(8): 2662-2675. DOI:10.1890/0012-9658(1999)080[2662:EFATLS]2.0.CO;2 |
| [11] |
John R, Dalling J W, Harms K E, Yavitt J B, Stallard R F, Mirabello M, Hubbell S P, Valencia R, Navarrete H, Vallejo M, Foster R B. Soil nutrients influence spatial distributions of tropical tree species. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(3): 864-869. DOI:10.1073/pnas.0604666104 |
| [12] |
赵成章, 张静, 盛亚萍. 高寒山区一年生混播牧草生态位对密度的响应. 生态学报, 2013, 33(17): 5266-5273. |
| [13] |
陈波, 周兴民. 三种嵩草群落中若干植物种的生态位宽度与重叠分析. 植物生态学报, 1995, 19(2): 158-169. DOI:10.3321/j.issn:1005-264X.1995.02.007 |
| [14] |
Chesson P. Mechanisms of maintenance of species diversity. Annual Review of Ecology and Systematics, 2000, 31: 343-366. DOI:10.1146/annurev.ecolsys.31.1.343 |
| [15] |
Tilman D. Causes, consequences and ethics of biodiversity. Nature, 2000, 405(6783): 208-211. DOI:10.1038/35012217 |
| [16] |
Silvertown J. Plant coexistence and the niche. Trends in Ecology & Evolution, 2004, 19(11): 605-611. |
| [17] |
Murrell D J, Law R. Heteromyopia and the spatial coexistence of similar competitors. Ecology Letters, 2003, 6(1): 48-59. |
| [18] |
宋乃平, 王兴, 陈林, 薛毅, 陈娟, 随金明, 王磊, 杨新国. 荒漠草原"土岛"生境群落物种共存机制. 生物多样性, 2018, 26(7): 667-677. |
| [19] |
李国旗, 邵文山, 赵盼盼, 靳长青. 封育对荒漠草原两种植物群落土壤种子库的影响. 草业学报, 2018, 27(6): 52-61. |
| [20] |
龚容, 高琼, 王亚林. 围封对温带半干旱典型草原群落种间关联的影响. 植物生态学报, 2016, 40(6): 554-563. |
| [21] |
高雪峰, 韩国栋, 张国刚. 短花针茅荒漠草原土壤微生物群落组成及结构. 生态学报, 2017, 37(15): 5129-5136. |
| [22] |
Luo W C, Zhao W Z, Liu B. Growth stages affect species richness and vegetation patterns of nebkhas in the desert steppes of China. CATENA, 2016, 137: 126-133. DOI:10.1016/j.catena.2015.09.011 |
| [23] |
Liu B, Zhao W Z, Liu Z L, Yang Y T, Luo W C, Zhou H, Zhang Y Y. Changes in species diversity, aboveground biomass, and vegetation cover along an afforestation successional gradient in a semiarid desert steppe of China. Ecological Engineering, 2015, 81: 301-311. DOI:10.1016/j.ecoleng.2015.04.014 |
| [24] |
闫宝龙, 王忠武, 屈志强, 王静, 韩国栋. 围封对内蒙古典型草原与荒漠草原植被-土壤系统碳密度的影响. 植物生态学报, 2018, 42(3): 327-336. |
| [25] |
刘文亭, 卫智军, 吕世杰, 王天乐, 张爽. 放牧对短花针茅荒漠草原植物多样性的影响. 生态学报, 2017, 37(10): 3394-3402. |
| [26] |
李永强, 李治国, 董智, 王忠武, 屈志强, 韩国栋. 内蒙古荒漠草原放牧强度对风沙通量和沉积物粒径的影响. 植物生态学报, 2016, 40(10): 1003-1014. DOI:10.17521/cjpe.2016.0045 |
| [27] |
阎欣, 安慧. 宁夏荒漠草原沙漠化过程中土壤粒径分形特征. 应用生态学报, 2017, 28(10): 3243-3250. |
| [28] |
沈艳, 刘彩凤, 马红彬, 赵菲, 谢应忠. 荒漠草原土壤种子库对草地管理方式的响应. 生态学报, 2015, 35(14): 4725-4732. |
| [29] |
宋乃平, 杜灵通, 王磊. 盐池县2000-2012年植被变化及其驱动力. 生态学报, 2015, 35(22): 7377-7386. |
| [30] |
Tang Z S, An H, Shangguan Z P. The impact of desertification on carbon and nitrogen storage in the desert steppe ecosystem. Ecological Engineering, 2015, 84: 92-99. DOI:10.1016/j.ecoleng.2015.07.023 |
| [31] |
沈泽昊, 方精云. 基于种群分布地形格局的两种水青冈生态位比较研究. 植物生态学报, 2001, 25(4): 392-398. DOI:10.3321/j.issn:1005-264X.2001.04.002 |
| [32] |
任继周. 草业科学研究方法. 北京: 中国农业出版社, 1998.
|
| [33] |
张继义, 赵哈林, 张铜会, 赵学勇. 科尔沁沙地植物群落恢复演替系列种群生态位动态特征. 生态学报, 2003, 23(12): 2741-2746. DOI:10.3321/j.issn:1000-0933.2003.12.030 |
| [34] |
王鑫, 刘钦, 黄琴, 张华雨, 李宗峰, 张世强, 邓洪平. 崖柏群落优势种生态位及CCA排序分析. 北京林业大学学报, 2017, 39(8): 60-67. |
| [35] |
唐以杰, 余世孝, 柯芝军. 红树林区软体动物生态位的三种分析方法. 生态学杂志, 2006, 25(11): 1442-1448. DOI:10.3321/j.issn:1000-4890.2006.11.028 |
| [36] |
张海艳, 孙国俊, 季敏, 李粉华, 韩敏, 许映莲, 万玉成. 苏南丘陵茶园秋季杂草种间生态关系及群落分类. 生态学杂志, 2013, 32(9): 2289-2297. |
| [37] |
董林水, 张旭东, 周金星, 宋爱云. 晋西黄土区与土石山区交错地带灌木种生态位及排序分析. 南京林业大学学报:自然科学版, 2007, 31(5): 15-20. |
| [38] |
曹杨, 上官铁梁, 张金屯, 闫美芳, 聂二保. 山西五台山蓝花棘豆群落的数量分类和排序. 植物资源与环境学报, 2005, 14(3): 1-6. DOI:10.3969/j.issn.1674-7895.2005.03.001 |
| [39] |
刘小丹, 张克斌, 王黎黎, 杨晓晖. 封育对半干旱区沙化草地群落特征的影响. 北京林业大学学报, 2015, 37(2): 48-54. |
| [40] |
徐治国, 何岩, 闫百兴, 宋长春. 三江平原典型沼泽湿地植物种群的生态位. 应用生态学报, 2007, 18(4): 783-787. DOI:10.3321/j.issn:1001-9332.2007.04.013 |
| [41] |
钱逸凡, 伊力塔, 胡军飞, 张超, 余树全, 沈露, 彭东琴. 普陀山主要植物种生态位特征. 生态学杂志, 2012, 31(3): 561-568. |
| [42] |
原野, 赵中秋, 白中科, 王怀泉, 徐志果, 牛姝烨. 安太堡露天煤矿不同复垦模式下草本植物优势种生态位. 生态学杂志, 2016, 35(12): 3215-3222. |
| [43] |
李斌, 李素清, 张金屯. 云顶山亚高山草甸优势种群生态位研究. 草业学报, 2010, 19(1): 6-13. |
| [44] |
彭文俊, 王晓鸣. 生态位概念和内涵的发展及其在生态学中的定位. 应用生态学报, 2016, 27(1): 327-334. |
| [45] |
吴会峰, 宋丽娟, 杜峰, 郝文芳. 黄土丘陵区草地主要种群生态位及其环境解释. 应用生态学报, 2017, 28(11): 3494-3504. |
| [46] |
杜峰, 山仑, 梁宗锁, 徐学选. 陕北黄土丘陵区撂荒演替生态位研究. 草业学报, 2006, 15(3): 27-35. DOI:10.3321/j.issn:1004-5759.2006.03.004 |
| [47] |
Levine J M, Hillerislambers J. The importance of niches for the maintenance of species diversity. Nature, 2009, 461(7261): 254-257. DOI:10.1038/nature08251 |
| [48] |
王刚. 生态位理论若干问题探讨. 兰州大学学报:自然科学版, 1990, 26(2): 109-113. |
| [49] |
林勇, 艾训儒, 姚兰, 黄伟, 陈斯. 木林子自然保护区不同群落类型主要优势种群的生态位研究. 自然资源学报, 2017, 32(2): 223-234. |
| [50] |
井光花, 程积民, 苏纪帅, 魏琳, 史晓晓, 金晶炜. 黄土区长期封育草地优势物种生态位宽度与生态位重叠对不同干扰的响应特征. 草业学报, 2015, 24(9): 43-52. |
| [51] |
An P, Li X J, Zheng Y R, Eneji A E, Qiman Y, Zheng M Q, Inanaga S. Distribution of plant species and species-soil relationship in the east central Gurbantunggut Desert, China. Journal of Geographical Sciences, 2015, 25(1): 101-112. DOI:10.1007/s11442-015-1156-0 |
| [52] |
黄至欢, 陆奇丰, 陈颖卓. 地锦苗在石灰岩土壤和红壤生境中的繁殖成功的比较. 生物多样性, 2017, 25(9): 972-980. |
| [53] |
王正文, 祝廷成. 松嫩草原主要草本植物的生态位关系及其对水淹干扰的响应. 草业学报, 2004, 13(3): 27-33. DOI:10.3321/j.issn:1004-5759.2004.03.005 |
| [54] |
杜华栋, 焦菊英, 寇萌, 苗芳. 黄土高原先锋种猪毛蒿叶片形态解剖与生理特征对立地的适应性. 生态学报, 2016, 36(10): 2914-2925. |
| [55] |
白晓航, 张金屯. 小五台山森林群落优势种的生态位分析. 应用生态学报, 2017, 28(12): 3815-3826. |
| [56] |
金海洙, 金锺熙. 猪毛蒿对其他植物生长的他感作用. 烟台大学学报:自然科学与工程版, 2001, 14(3): 180-184. |
| [57] |
王勇, 梁宗锁, 龚春梅, 韩蕊莲, 于靖. 干旱胁迫对黄土高原4种蒿属植物叶形态解剖学特征的影响. 生态学报, 2014, 34(16): 4535-4548. |
| [58] |
Legendre P, Mi X C, Ren H B, Ma K P, Yu M J, Sun I F, He F L. Partitioning beta diversity in a subtropical broad-leaved forest of China. Ecology, 2009, 90(3): 663-674. DOI:10.1890/07-1880.1 |
| [59] |
Chase J M. Towards a really unified theory for metacommunities. Functional Ecology, 2005, 19(1): 182-186. DOI:10.1111/j.0269-8463.2005.00937.x |
| [60] |
储诚进, 王酉石, 刘宇, 蒋林, 何芳良. 物种共存理论研究进展. 生物多样性, 2017, 25(4): 345-354. |
| [61] |
牛克昌, 刘怿宁, 沈泽昊, 何芳良, 方精云. 群落构建的中性理论和生态位理论. 生物多样性, 2009, 17(6): 579-593. |
| [62] |
Wilson J B, Gitay H. Limitations to species coexistence:evidence for competition from field observations, using a patch model. Journal of Vegetation Science, 1995, 6(3): 369-376. DOI:10.2307/3236236 |
| [63] |
Grime J P. Trait convergence and trait divergence in herbaceous plant communities:mechanisms and consequences. Journal of Vegetation Science, 2006, 17(2): 255-260. DOI:10.1111/j.1654-1103.2006.tb02444.x |