 2019, Vol. 39
2019, Vol. 39文章信息
- 王小平, 杨雪, 杨楠, 辛晓静, 曲耀冰, 赵念席, 高玉葆
- WANG Xiaoping, YANG Xue, YANG Nan, XIN Xiaojing, QU Yaobing, ZHAO Nianxi, GAO Yubao
- 凋落物多样性及组成对凋落物分解和土壤微生物群落的影响
- Effects of litter diversity and composition on litter decomposition characteristics and soil microbial community
- 生态学报. 2019, 39(17): 6264-6272
- Acta Ecologica Sinica. 2019, 39(17): 6264-6272
- http://dx.doi.org/10.5846/stxb201805251153
-
文章历史
- 收稿日期: 2018-05-25
- 网络出版日期: 2019-05-31

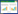




植物凋落物是土壤肥力的主要来源, 而凋落物分解是生态系统碳和营养循环的核心过程, 是生态学领域研究的核心内容之一。近年来, 由于人类活动和全球气候变化, 生物多样性丧失严重, 群落物种组成及结构均发生显著改变[1]。已有研究表明, 陆地生态系统当年生物量的90%都是以凋落物形式存在[2], 群落物种组成及结构的改变势必对凋落物基质质量以及对应的分解者群落、微环境产生显著影响, 从而对生态系统养分循环产生显著影响[3]。因此, 了解不同物种及不同物种组合对凋落物分解特性和土壤微生物群落的影响, 将帮助我们深入了解群落物种及组成变化对生态系统养分循环的影响, 进而为全球变化导致的群落组成和结构变化的生态后果进行准确评估。
微生物作为连接植物与土壤物质循环最重要的因子, 对土壤的形成、发育、结构改良、肥力保持以及植物生长等方面都具有重要作用, 其在凋落物分解中的作用以及与凋落物的关系一直是生态学领域的重点研究内容之一。如土壤微生物在凋落物分解过程中所起作用, 以及凋落物分解对土壤生物群落结构和功能特征影响等方面均积累了大量的研究成果[4-7]。而关于凋落物多样性及组成变化如何影响微生物群落的研究相对较少且主要集中在森林中, 如李姗姗等[8]在研究江西省德兴市森林生态系统中凋落物分解183 d后发现, 凋落物物种多样性与土壤微生物群落多样性呈负相关关系;Sarah等[9]以美国佛罗里达森林的10种单种凋落物为研究对象, 发现相似的凋落物化学组成, 其微生物群落也更相似。
草原凋落物在草原生态系统生产力构成中占有重要地位, 是草原生态系统物质和能量的主要来源, 是土壤与微生物间的物质循环枢纽。作为生物地球化学循环的关键环节, 凋落物在草原土壤质量的维护和生态功能的发挥上具有重要作用。呼伦贝尔草原位于内蒙古东部地区, 是我国北方草原的主要代表。然而, 由于长期过度放牧的影响, 该区域草原出现了不同程度的退化, 生物多样性和稳定性降低, 草原群落的建群种出现了明显替代现象[10-11], 群落的物种多样性及其组成均发生了显著的变化, 进而导致凋落物的种类多样性及其组成的变化。因此, 本研究在该地区选择一原生群落贝加尔针茅(Stipa baicalensis)-羊草(Leymus chinensis)的退化恢复区域, 依据植物多度、盖度、频度和物种的重要值及其在群落中的恢复程度筛选出排序前4的羊草、茵陈蒿(Artemisia capillaris)、麻花头(Serratula centauroides)和二裂委陵菜(Potentilla bifurca)[12]的凋落物为实验材料, 通过设置物种数目以及不同物种组合处理, 在人工控制条件下, 研究不同组合处理的凋落物分解特性与分解过程中的土壤微生物群落结构对不同凋落物组合处理的响应。本实验重点关注以下问题:凋落物分解一段时间后, (1)不同处理的凋落物如何影响凋落物分解特性?如何影响土壤微生物群落?以及三者之间的关系如何?(2)物种数目和物种组成, 哪个因子对凋落物分解及土壤微生物群落影响更大?
1 材料与方法 1.1 采集地概况及实验材料采集地为内蒙古自治区呼伦贝尔市鄂温克族自治旗北辉野外科学观测研究站的围封样地(48.15°-49.00°N, 118.80°-119.75°E), 该区域土壤类型为暗栗钙土,原生植被为贝加尔针茅-羊草草原, 但由于长期过度放牧导致群落严重退化, 因此自2008年开始围封禁牧。到2014年, 经野外调查结果显示共有物种45种, 与稳定群落的物种数目和多样性相似, 显著高于放牧区物种数(总物种数为25种)。在近7年的围封恢复过程中, 恢复速度最快的4种植物为:羊草、茵陈蒿、麻花头和二裂委陵菜。这4种植物的相对盖度、相对多度和相对高度达到总盖度、多度和频度的40%左右, 而贝加尔针茅、大针茅(Stipa grandis)和冰草(Agropyron cristatum)在群落中的多度仍较低。在2014年8月初, 收集羊草、茵陈蒿、麻花头和二裂委陵菜4种植物的凋落物, 风干至恒重;并用土钻收集地表(0-10 cm)新鲜土壤。
1.2 实验设计本实验为单因素嵌套实验,因素为凋落物多样性(Litter Diversity, LD), 包括3个水平(凋落物组成嵌套在凋落物多样性中):(1)单种凋落物(LD1), 即4种凋落物(羊草、茵陈蒿、麻花头和二裂委陵菜)单独分解;(2)两种凋落物混合(LD2), 包括所选4种凋落物分别进行两两等量混合(羊草+茵陈蒿、羊草+麻花头、羊草+二裂委陵菜、茵陈蒿+麻花头、茵陈蒿+二裂委陵菜、麻花头+二裂委陵菜), 共6种凋落物组成;(3)4种凋落物混合(LD4), 即4种凋落物(羊草+茵陈蒿+麻花头+二裂委陵菜)等量混合,即包括3个凋落物多样性水平共11种组合, 并用凋落物组成中植物属名首字母表示这11种组合, 即为L, A, S, P, L+A, L+S, L+P, A+S, A+P, S+P, L+A+S+P。每种凋落物组成3个重复, 共66袋。留取4种植物的部分凋落物, 测定凋落物初始N、C以及木质素含量, 并计算C/N, 每种凋落物测定3次。
将等量组合好的凋落物1.2 g分别装入网孔面积为1 mm2的凋落物袋(10 cm×15 cm)并进行编号, 此种凋落物袋允许土壤微生物以及土壤小型动物进出。2015年2月1日, 将原生境地表土壤放在400 μmol/mol CO2浓度的智能人工气候箱中, 设置其他参数为:温度25 ℃, 湿度50%, 光强60 μmol m-2 s-1, 光照12 h。2015年2月11日, 将适应的土壤混匀后转入直径为12 cm的塑料器皿中, 每皿50 g, 将土壤湿度调为田间持水量, 并将准备好的凋落物袋放置于土壤表面, 对应编号。所有培养皿随机放置, 每隔2天移动位置1次以避免位置效应, 60 d后收获凋落物用于凋落物分解特性(凋落物质量、C、N残余率及C/N)分析, 并将对应的土壤材料用于土壤微生物群落PLFA的测定。
1.3 凋落物化学成分的测定凋落物总C和总N含量的测定使用Vario MICRO cube元素分析仪(ELEMEMTAR, Germany)。
木质素的测定采用凡氏法, 参考Antonietta等[13]。
1.4 凋落物质量损失以及养分变化凋落物质量残余率:

|
凋落物C、N残余率:
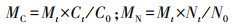
|
式中, Mt表示分解t时间后凋落物残余质量占初始质量的比例(%);MT表示分解t时间后凋落物残余质量(g);M0表示凋落物初始质量(g);MC、MN表示分解t时间后凋落物C、N含量占初始含量的比例(%);C0、N0表示凋落物的初始养分浓度(%);Ct、Nt表示分解时间t后凋落物的C、N浓度(%)。
1.5 土壤微生物测定采用磷脂脂肪酸(Phospholipid Fatty Acid, PLFA)温和甲酯化法[14]和气质联用色谱分析仪(Agilent 7890GC, 5975MSD)进行PLFA生物标记分离及鉴定, 含量计算[15]和表征分类[16-18], 其中, 表征细菌的有:14:0, 15:0, 16:0, 17:0, 18:0, 16:1ω5t, 17:1ω7c, i14:0, i15:0, i16:0, i17:0, a16:0, 16:1ω7c, 18:1ω5c, cy17:0, cy19:0, 17:1ω8, 10Me16:0, 10Me17:0, 10Me18:0;表征真菌的有:18:1ω9c, 18:1ω9t, 18:2ω6c, 18:2ω6t, 18:3ω6c。
1.6 数据分析PLFA生物标记含量计算公式为:PLFA(nmol/g DW)=(PPLFA×S×V)/(POSTD×D×R×W×M)
式中, PPLFA和POSTD分别是样品和标准物质的峰值面积, S为内标标准物质的浓度(ng/μL), D为稀释倍数, R为分取倍数, V为样品的测定体积(μL), W为土壤干重(g DW), M为对应的PLFA的相对分子质量。
采用SPSS 21.0统计学软件中的一般线性模型分析4种凋落物初始化学性质之间的差异显著性;用一般线性模型中的多变量模型(设定平方和为:类型Ⅲ)分析凋落物多样性及组成对凋落物分解相关指标(凋落物质量、C、N残余率及C/N)和各类土壤微生物PLFAs含量影响的显著性, 其中凋落物组成作为主体内因素嵌套在凋落物多样性内。
采用CANOCO(Canoco for Windows 4.5)软件对凋落物分解相关指标(凋落物质量、C、N残余率及C/N)及其土壤微生物(细菌和真菌含量)与凋落物多样性及组成之间进行冗余分析(Redundancy Analysis, RDA)。
建立结构方程模型(Structural Equation Model, SEM)分析不同凋落物组合下, 凋落物初始化学性质(C、N、木质素含量及C/N)、土壤微生物(细菌和真菌含量)与凋落物分解相关指标(凋落物质量、C、N残余率及C/N)的关系, SEM计算和建立采用R语言(版本3.3.1)分析软件, 在程序包“sem”、“dplyr”及“DiagrammeR”下运行。
2 结果与分析 2.1 凋落物初始化学性质羊草凋落物的N和C含量显著高于其他凋落物, 而C/N和木质素含量显著低于其他凋落物(P < 0.05)。麻花头凋落物的C/N含量显著高于其他凋落物, 而N含量显著低于其他凋落物(P < 0.05)。麻花头和二裂委陵菜凋落物的木质素含量显著高于其他凋落物(P < 0.05)(表 1)。
| 物种 Species |
氮/% N |
碳/% C |
C/N | 木质素/(g/g) Lignin |
| 麻花头(S) S. centauroides | 0.75±0.02d | 37.30±0.51b | 50.14±1.22a | 0.31±0.01a |
| 羊草(L) L. chinensis | 1.50±0.01a | 39.63±0.20a | 26.54±0.32d | 0.16±0.00c |
| 二裂委陵菜(P) P. bifurca | 1.01±0.01b | 37.93±0.07b | 37.73±0.48c | 0.32±0.00a |
| 茵陈蒿(A) A. capillaris | 0.95±0.02c | 38.05±0.07b | 40.28±0.81b | 0.19±0.00b |
| 同列中不同小写字母表示差异显著(P < 0.05) | ||||
凋落物多样性仅对凋落物C残余率有显著影响(P < 0.05)(表 2), 表现在两物种混合显著低于单物种(图 1)。而凋落物组成对凋落物质量、C、N残余率及C/N均有显著影响(P < 0.05)(表 2)。
| 分解参数 Decomposition parameters |
凋落物多样性 Litter diversity(df=2) |
凋落物组成 Litter composition(df=8) |
||
| F | P | F | P | |
| 质量残余率Mass remaining rate/% | 0.142 | 0.868 | 2.808 | 0.026 |
| 碳残余率C remaining rate/% | 6.599 | 0.006 | 3.771 | 0.006 |
| 氮残余率N remaining rate/% | 1.705 | 0.205 | 3.251 | 0.013 |
| 碳氮比C/N | 3.225 | 0.059 | 3.825 | 0.006 |
| 同列不同小写字母表示差异显著(P < 0.05) | ||||

|
| 图 1 凋落物多样性对凋落物C残余率的影响 Fig. 1 The effects of litter diversity on litter C remaining rate |
凋落物组合中, 质量残余率方面:4个单物种组合间并没有显著差异(P>0.05);两物种组合中, 麻花头和二裂委陵菜(S+P)组合以及茵陈蒿和二裂委陵菜(A+P)组合最高, 而羊草和茵陈蒿(L+A)组合最低, 前者与后者差异显著(P < 0.05);四物种组合(LD4)与任何组合间均无显著(P>0.05)差异(图 2)。

|
| 图 2 凋落物组成对凋落物质量、C、N残余率及C/N的影响 Fig. 2 The effect of litter composition on litter mass, C, N remaining rate and C/N LD:凋落物多样性Litter diversity;S:麻花头S. centauroides;L:羊草L. chinensis;P:二裂委陵菜P. bifurca;A:茵陈蒿A. capillaris |
C残余率方面:单物种组合中, 麻花头(S)、二裂委陵菜(P)单物种组合的残余率显著(P < 0.05)高于羊草(L)和茵陈蒿(A);两物种组合中, 麻花头和二裂委陵菜(S+P)组合最高, 显著(P < 0.05)高于其他5种组合, 而羊草和茵陈蒿(L+A)组合最低;四物种组合(LD4)显著(P < 0.05)高于羊草和茵陈蒿(L+A)组合(图 2)。
N残余率方面:11种组合中, 单物种麻花头(S)残余率最高, 而两物种组合羊草和茵陈蒿(L+A)最低(图 2)。
凋落物C/N方面:单物种组合中羊草(L)最低, 显著低于其他3种植物, 而其他3种植物间无显著(P>0.05)差异;两物种组合中, 麻花头和二裂委陵菜(S+P)最高, 羊草和二裂委陵菜(L+P)最低, 两者之间差异显著(P < 0.05);四物种组合的C/N显著(P < 0.05)低于单物种茵陈蒿(A), 与其他组合间均无显著(P>0.05)差异。
2.3 凋落物多样性及组成对土壤微生物群落结构的影响仅细菌PLFAs含量受凋落物多样性影响显著(P < 0.05)(表 3), 表现为两物种组合(LD2)显著高于LD4(图 3);仅真菌含量受凋落物组成影响显著(P < 0.05)(表 3), 具体表现为:单物种条件下, 单物种麻花头(S)处理下土壤真菌PLFAs含量显著(P < 0.05)高于其他3种凋落物;而多物种组合间无显著(P>0.05)差异;羊草和二裂委陵菜(L+P)组合土壤真菌PLFAs含量显著(P < 0.05)高于单种羊草(L)的土壤真菌PLFAs含量(图 4)。
| 微生物 Microbe |
凋落物多样性Litter diversity(df=2) | 凋落物组成Litter composition(df=8) | |||
| F | P | F | P | ||
| 细菌(B) Bacteria | 3.787 | 0.034 | 0.491 | 0.850 | |
| 真菌(F) Fungi | 0.400 | 0.675 | 2.779 | 0.027 | |
| F/B | 0.299 | 0.745 | 2.181 | 0.071 | |
| 总微生物T Total microbe | 2.169 | 0.138 | 1.379 | 0.259 | |
| 同列不同小写字母表示差异显著(P < 0.05) | |||||

|
| 图 3 凋落物多样性对土壤细菌PLFAs含量的影响s Fig. 3 The effects of litter diversity on the soil bacterial PLFAs content |

|
| 图 4 凋落物组成对土壤真菌PLFAs含量的影响 Fig. 4 The effects of litter composition on the soil fungi PLFAs content S:麻花头S.centauroides; L:羊草L. chinensis;P:二裂委陵菜P. bifurca; A:茵陈蒿A. capillaris |
RDA排序分析表明:前两个排序轴共解释了53%的变异和90.4%的环境变量与响应变量的关系(图 5)。蒙特卡罗检验结果显示, 所有排序轴对应的环境变量对于响应变量的解释贡献均达到统计学上的显著水平(F=3.15, P=0.022), 其中凋落物多样性及组成的影响大小排序为羊草(L)(26%)>麻花头(S)(18%)>二裂委陵菜(P)(14%)=茵陈蒿(A)(14%)>LD(4%)。通过凋落物分解特性和土壤微生物与凋落物多样性及组成相关性排序可知:细菌含量, 真菌含量, 凋落物质量、C、N残余率及C/N与含羊草(L)凋落物处理呈显著负相关;真菌含量, 凋落物质量、C、N残余率与含麻花头(S)凋落物及含二裂委陵菜(P)凋落物的处理均呈显著正相关;细菌含量, 凋落物C/N与含茵陈蒿(A)凋落物的处理呈显著正相关;细菌含量, 凋落物C/N与凋落物多样性呈显著负相关。

|
| 图 5 土壤微生物和凋落物分解特性与凋落物多样性及组成的冗余分析 Fig. 5 Redundancy analysis of the soil microbe and litter decomposition characteristics and litter diversity and composition CNR:残余碳氮比C/N remaining;CR:碳残余率C remaining rate;NR:氮残余率N remaining rate;MR:质量残余率Mass remaining rate |
由SEM可知(图 6),凋落物初始C含量对凋落物质量、C、N残余率及C/N有显著正的直接影响;凋落物木质素含量对凋落物质量、C、N残余率有显著正的直接影响;凋落物初始N含量对N残余率为显著正的直接影响, 而对C残余率及C/N为显著负的直接影响;凋落物初始C/N对凋落物质量、N残余率为显著正的直接影响, 而对C/N为显著负的直接影响(图 6)。此外, 凋落物初始C、木质素、N含量及C/N均对真菌含量具有显著正影响, 并可通过真菌对凋落物质量残余率产生显著负的间接影响。

|
| 图 6 凋落物初始化学性质、土壤微生物与凋落物分解特性的结构方程模型 Fig. 6 The Structural equation model of initial litter chemical properties, soil microbes and litter decomposition characteristics 仅提供P < 0.05的相关关系,实线和虚线箭头表示正和负路径,箭头宽度与关系强度成正比,*P<0.05,**P<0.01,***P<0.001表示差异显著 |
近年来, 凋落物多样性/组成对凋落物分解的影响[19-21]及对土壤微生物群落的影响[6, 22-23]受到研究者的广泛关注。但将凋落物多样性/组成对凋落物分解、土壤微生物群落的影响及其内在关系共同展开研究的仅有少量报道[9, 24]。如Santonja等[24], 在法国马赛附近的灌木丛林中, 研究了凋落物多样性对凋落物分解和微生物群落的影响, 结果发现:凋落物C、N的释放随凋落物种类的增加而增加;多物种凋落物混合, 细菌多样性较低而真菌多样性较高;微生物丰度及多样性对混合凋落物C、N分解的影响较小。这一研究表明凋落物多样性对凋落物C、N的释放影响更大, 而微生物对凋落物C、N的释放影响较弱。然而, 本研究结果显示:尽管凋落物多样性对凋落物C残余率以及土壤细菌群落均具有显著影响(表 2, 3), 但结合结构方程模型及冗余分析的结果可以看出, 凋落物组成对凋落物分解特性(凋落物质量、C、N残余率及C/N)及土壤微生物(细菌和真菌含量)影响更大, 且真菌群落对凋落物质量分解影响显著。因此, 本研究结果表明在典型草原退化恢复群落中, 凋落物组成而非凋落物多样性, 对凋落物的分解及其地下部土壤微生物群落具有决定性作用。
凋落物组成, 如混合凋落物初始性质, 是影响凋落物分解和土壤微生物群落的主要因素。本研究中羊草初始木质素含量和C/N显著低于其余3种凋落物(表 1), 含羊草的凋落物质量、C、N残余率低(图 2);而麻花头和二裂委陵菜C/N和木质素含量高(表 1), 含二者的凋落物分解质量、C、N残余率高(图 2), 支持凋落物的初始C/N和木质素含量的高低是凋落物分解快慢的重要指标[25-26],即低C/N和低木质素含量的凋落物主要由细菌分解,分解较快; 而高C/N和高木质素含量的凋落物主要由真菌分解,分解较慢[9, 27-29]。如本研究冗余分析结果显示, 初始木质素含量和C/N显著低于其他3种凋落物的羊草与凋落物质量、C、N残余率呈显著负相关(图 5);而初始木质素含量显著高于其他两种凋落物的麻花头和二裂委陵菜与土壤真菌含量, 凋落物质量、C、N残余率呈显著正相关(图 5)。此外, 本研究还发现, 不同混合的凋落物初始性质不仅与凋落物残余率具有直接的正相关关系, 还通过真菌群落间接影响凋落物质量残余率, 即真菌含量与质量残余率呈显著负相关(图 6)。不同凋落物处理的分解者群落是以真菌为主导, 这除了能反映凋落物性质不易分解外[9], 还反映凋落物分解速率缓慢[29], 同时也进一步暗示着生态系统物质循环的缓慢。
作为原生群落的主要物种之一羊草的C/N和木质素含量在分解初期显著低于麻花头、茵陈蒿和二裂委陵菜(表 1), 而原生群落的另一主要物种贝加尔针茅的C/N在30左右[30-31], 也显著低于本实验中麻花头、茵陈蒿以及和二裂委陵菜凋落物的C/N比值, 这些指标均有利于细菌分解、分解较快。基于以上可知:贝加尔针茅-羊草退化群落恢复7年后, 根据植物多度、盖度、频度和物种的重要值及其在群落中的恢复程度排序, 选取4位植物(盖度或多度之和达40%)进行凋落物分解研究, 得出凋落物组成对凋落物分解起决定性作用(图 6), 且此群落在凋落物分解初期就表现为高C/N和高木质素含量(表 1), 并由真菌主导(图 5-6), 此现状或许是草原生态系统恢复缓慢的原因之一。因为凋落物分解及其土壤微生物群落结构是生态系统物质循环的功能指标, 对生态系统的健康具有指示作用。本研究结果表明在退化群落恢复过程中, 地上部植物的种类、盖度和多样性的恢复, 并不能很好地预测该生态系统的功能, 只有结合凋落物分解特征以及微生物群落的结构和功能才能对生态系统功能进行合理评价。
| [1] |
王珍.锡林河流域典型草原生物多样性及其与生产力的关系[D].呼和浩特: 内蒙古大学, 2017.
|
| [2] |
Cebrian J. Patterns in the fate of production in plant communities. American Naturalist, 1999, 154(4): 449-468. DOI:10.1086/303244 |
| [3] |
Swift M J, Heal O W, Anderson J M. Decomposition in terrestrial ecosystems. Studies in Ecology, 1979, 5: 372-372. |
| [4] |
Potthoff M, Dyckmans J, Flessa H, Beese F, Joergensen R G. Decomposition of maize residues after manipulation of colonization and its contribution to the soil microbial biomass. Biology and Fertility of Soils, 2008, 44(6): 891-895. DOI:10.1007/s00374-007-0266-y |
| [5] |
许湘琴, 林植华, 陈慧丽. 凋落物分解对土壤生物的影响. 生态学杂志, 2011, 30(6): 1258-1264. |
| [6] |
Scheibe A, Steffens C, Seven J, Jacob A, Hertel D, Leuschner C, Gleixner G. Effects of tree identity dominate over tree diversity on the soil microbial community structure. Soil Biology and Biochemistry, 2015, 81: 219-227. DOI:10.1016/j.soilbio.2014.11.020 |
| [7] |
Xiao C W, Zhou Y, Su J Q, Yang F. Effects of plant functional group loss on soil microbial community and litter decomposition in a steppe vegetation. Frontiers in Plant Science, 2017, 8: 2040. DOI:10.3389/fpls.2017.02040 |
| [8] |
李姗姗, 王正文, 杨俊杰. 凋落物分解过程中土壤微生物群落的变化. 生物多样性, 2016, 24(2): 195-204. |
| [9] |
Bray S R, Kitajima K, Mack M C. Temporal dynamics of microbial communities on decomposing leaf litter of 10 plant species in relation to decomposition rate. Soil Biology and Biochemistry, 2012, 49: 30-37. DOI:10.1016/j.soilbio.2012.02.009 |
| [10] |
Xie Y Z, Wittig R. The impact of grazing intensity on soil characteristics of Stipa grandis and Stipa bungeana steppe in northern China (autonomous region of Ningxia). Acta Oecologica, 2004, 25(3): 197-204. DOI:10.1016/j.actao.2004.01.004 |
| [11] |
宋明华, 刘丽萍, 陈锦, 张宪洲. 草地生态系统生物和功能多样性及其优化管理. 生态环境学报, 2018, 27(6): 1179-1188. |
| [12] |
辛晓静.物种混合及CO2浓度倍增对呼伦贝尔草原区常见物种凋落物分解的影响[D].天津: 南开大学, 2016.
|
| [13] |
Spadaro A, Ajello A, Morace C, Zirilli A, D'Arrigo G, Luigiano C, Martino F, Bene A, Migliorato D, Turiano S, Ferraù O, Freni M A. Serum chromogranin-A in hepatocellular carcinoma:diagnostic utility and limits. World Journal of Gastroenterology, 2005, 11(13): 1987-1990. DOI:10.3748/wjg.v11.i13.1987 |
| [14] |
Zelles L. Fatty acid patterns of phospholipids and lipopolysaccharides in the characterisation of microbial communities in soil:a review. Biology and Fertility of Soils, 1999, 29(2): 111-129. DOI:10.1007/s003740050533 |
| [15] |
辛晓静, 刘磊, 申俊芳, 赵念席, 高玉葆. 羊草基因型数目与氮添加对土壤微生物群落的交互影响. 生态学报, 2016, 36(13): 3923-3932. |
| [16] |
Zhou Y, Clark M, Su J Q, Xiao C W. Litter decomposition and soil microbial community composition in three Korean pine (Pinus koraiensis) forests along an altitudinal gradient. Plant and Soil, 2015, 386(1/2): 171-183. |
| [17] |
Chang C H, Wu F Z, Yang W Q, Xu Z F, Cao R, He W, Tan B, Justine M F. The microbial community in decaying fallen logs varies with critical period in an alpine forest. PLoS One, 2017, 12(8): e0182576. DOI:10.1371/journal.pone.0182576 |
| [18] |
Thoms C, Gleixner G. Seasonal differences in tree species' influence on soil microbial communities. Soil Biology and Biochemistry, 2013, 66: 239-248. DOI:10.1016/j.soilbio.2013.05.018 |
| [19] |
Li S S, Tong Y W, Wang Z W. Species and genetic diversity affect leaf litter decomposition in subtropical broadleaved forest in southern China. Journal of Plant Ecology, 2017, 10(1): 232-241. DOI:10.1093/jpe/rtw122 |
| [20] |
Zuo X A, Knops J M H. Effects of elevated CO2, increased nitrogen deposition, and plant diversity on aboveground litter and root decomposition. Ecosphere, 2018, 9(2): e02111. DOI:10.1002/ecs2.2111 |
| [21] |
López-Rojo N, Martínez A, Pérez J, Basaguren A, Pozo J, Boyero L. Leaf traits drive plant diversity effects on litter decomposition and FPOM production in streams. PLoS One, 2018, 13(5). DOI:10.1371/journal.pone.0198243 |
| [22] |
Chapman S K, Newman G S. Biodiversity at the plant-soil interface:microbial abundance and community structure respond to litter mixing. Oecologia, 2010, 162(3): 763-769. DOI:10.1007/s00442-009-1498-3 |
| [23] |
Wang H, Liu S R, Wang J X, Shi Z M, Xu J, Hong P Z, Ming A G, Yu H L, Chen L, Lu L H, Cai D X. Differential effects of conifer and broadleaf litter inputs on soil organic carbon chemical composition through altered soil microbial community composition. Scientific Reports, 2016, 6: 27097. DOI:10.1038/srep27097 |
| [24] |
Santonja M, Rancon A, Fromin N, Baldy V, Hättenschwiler S, Fernandez C, Montès N, Mirleau P. Plant litter diversity increases microbial abundance, fungal diversity, and carbon and nitrogen cycling in a Mediterranean shrubland. Soil Biology and Biochemistry, 2017, 111: 124-134. DOI:10.1016/j.soilbio.2017.04.006 |
| [25] |
Palm C A, Sanchez P A. Nitrogen release from the leaves of some tropical legumes as affected by their lignin and polyphenolic contents. Soil Biology and Biochemistry, 1991, 23(1): 83-88. DOI:10.1016/0038-0717(91)90166-H |
| [26] |
Kemp P R, Reynolds J F, Virginia R A, Whitford W G. Decomposition of leaf and root litter of Chihuahuan desert shrubs:effects of three years of summer drought. Journal of Arid Environments, 2003, 53(1): 21-39. DOI:10.1006/jare.2002.1025 |
| [27] |
Potthoff M, Steenwerth K L, Jackson L E, Drenovsky R E, Scow K M, Joergensen R G. Soil microbial community composition as affected by restoration practices in California grassland. Soil Biology and Biochemistry, 2006, 38(7): 1851-1860. DOI:10.1016/j.soilbio.2005.12.009 |
| [28] |
Jacob M, Leuschner C, Thomas F M. Productivity of temperate broad-leaved forest stands differing in tree species diversity. Annals of Forest Science, 2010, 67(5): 503-503. DOI:10.1051/forest/2010005 |
| [29] |
Austin A T, Ballaré C L. Dual role of lignin in plant litter decomposition in terrestrial ecosystems. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(10): 4618-4622. DOI:10.1073/pnas.0909396107 |
| [30] |
于雯超, 宋晓龙, 修伟明, 张贵龙, 赵建宁, 杨殿林. 氮素添加对贝加尔针茅草原凋落物分解的影响. 草业学报, 2014, 23(5): 49-60. |
| [31] |
李文亚, 邱璇, 白龙, 杨殿林. 氮、磷添加对贝加尔针茅草原凋落物分解的影响. 草业学报, 2017, 26(8): 43-53. |