 2019, Vol. 39
2019, Vol. 39文章信息
- 乔雪涛, 何俊, 范秀华, 孙兆军
- QIAO Xuetao, HE Jun, FAN Xiuhua, SUN Zhaojun
- 引黄灌区次生盐碱地速生杨光谱与光合特性对不同秸秆层的响应
- Response of spectral reflectance and photosynthetic characteristics of fast-growing poplar (Populus) to different straw layers in secondary saline-alkali land in the Yellow River irrigation area
- 生态学报. 2019, 39(15): 5573-5583
- Acta Ecologica Sinica. 2019, 39(15): 5573-5583
- http://dx.doi.org/10.5846/stxb201809051893
-
文章历史
- 收稿日期: 2018-09-05
- 网络出版日期: 2019-05-16





2. 宁夏大学新技术应用研究开发中心, 银川 750021
2. Development Center of New Technique Application and Research, Ningxia University, Yinchuan 750021, China
次生盐碱化是指由于不合理的人为措施的干扰与破坏, 所引发土壤发生盐碱化的过程, 易发生于具有潜在盐碱化因素如气候条件干旱、降水稀少、地下水位较高的地区[1]。大量研究表明, 过度开垦使得多年生深根的本土植物被一年生浅根植物(农作物)所替换, 以及长期不合理灌溉, 引起地下潜水上升, 导致深层的盐分随潜水不断上升并累积到地表, 是引发灌区土地次生盐碱化的主要原因[2]。统计表明, 我国西北地区盐碱化土地面积达2216×104 hm2, 约占全国盐碱地总面积的60%, 其中由于灌溉方式不当而引发的次生盐碱地面积约140×104 hm2, 占全国次生盐碱化土地总面积的70%, 已严重影响了当地农业的可持续发展与生态环境的恢复, 因此, 引黄灌区次生盐碱地的防治与植被恢复是当前迫切需要解决的问题[3-4]。
国内外许多研究者针对次生盐碱地区存在的科学难题进行了深入研究, 发现通过恢复和栽种优势深根植物, 能够增加土壤有机质, 改善土壤结构与环境, 降低地下水位, 调节区域小气候, 有助于将水文功能恢复到原始状态, 恢复植被的同时达到控盐改土的目的[2, 5-7]。但次生盐碱地土壤盐分高, 水分有效性差, 直接栽植会由于盐胁迫与干旱胁迫导致林木生长状况差, 造林的成活率低, 植被恢复困难[7]。以往的研究表明在地表覆盖秸秆能减少土表水分蒸发, 改善土壤水热状况, 活化土壤养分, 抑制盐分表聚;在地下铺设秸秆层能够打破长时间浅耕形成的坚固犁底层, 阻断土壤毛细管的连续性, 降低潜水蒸发, 阻隔盐分上行, 防止根层盐化[6-7]。但这些研究大多只注重秸秆层对盐碱土壤的水盐变化、改良利用及其对环境的危害等方面, 而对地上植物的生长、生理和受胁迫等植被恢复状况研究较少。
植物光合作用与植物的生长环境具有密切的相关性, 如适宜的环境会促进植物原有的光合潜能发挥, 通过研究植物光合特性能够有效反映出植物对生长环境的适应机制[8-12]。同时, 植物在不同的环境条件以及不同的生长发育阶段, 其体内色素含量、营养状态以及生理生化状况会发生不同的变化, 从而导致植物反射光谱发生变化[8-9]。利用光谱与光合特性能反映植被体内光合色素、生长状态以及受胁迫等情况[11-14]。因此本研究以西北灌区典型的次生盐碱地为例, 在土壤水盐变化的基础上, 通过分析不同措施下植物光谱及光合特性变化, 研究速生杨反射光谱及光合生理特性对不同秸秆层的响应。探究不同秸秆层的地上速生杨光合速率、色素含量、营养状况以及受胁迫等情况, 对进一步研究利用植物的反射光谱及光合特性等生理指标评估各类盐碱地改良措施的生态效益具有重要意义, 以期为引黄灌区次生盐碱地的土壤改良与植被恢复提供理论指导。
1 材料和方法 1.1 研究地概况试验地选在宁夏北部银川平原的青铜峡灌区(38°01′N, 105°57′E), 是引黄灌区次生盐碱地主要分布区。处于我国西北内陆干旱与半干旱地区, 属中温带大陆性气候。四季分明, 昼夜温差较大, 年平均气温8.3—8.6℃。全年日照时数2955 h, 无霜期176 d。气候条件干燥, 降水极少, 年均降水量仅为193—203 mm。试验地为典型引黄灌区次生盐碱地, 在试验开始前为弃耕的裸荒地, 土地表层盐分表聚明显, 地表有盐霜和结皮, 属重度盐碱地。在裸荒前种植过玉米(Zea mays)、枸杞(Lycium chinense)。试验地0—80 cm土层土壤理化性质如表 1所示。
| 土壤质地 Soil texture |
容重 Bulk density/(g/cm3) |
电导率 Electrical conductivity/(dS/m) |
pH | 钠吸附比 Sodium adsorption ratio/(mmol/L)0.5 |
田间持水量 Field capacity/% |
毛管孔隙度 Capillary porosity/% |
土壤养分Soil nutrient/(g/kg) | |||
| 有机质 Organic matter |
全氮 Total nitrogen |
有效磷 Available phosphorus |
有效钾 Available potassium |
|||||||
| 砂质壤土 Sandy loam |
1.52 | 17.9 | 7.2 | 15.5 | 21.3 | 28.1 | 3.43 | 0.31 | 7.36×10-3 | 5.53×10-2 |
速生杨是西北干旱与半干旱地区农田防护林和防风固沙林的主要树种, 生长迅速, 生物量大, 易繁殖, 根系发达, 生长季吸收养分能力强, 蒸腾作用高, 适应能力强, 在森林生态系统中占主导地位[15-16]。2016年4月, 本试验选取树龄为2年的速生杨带根苗作为试验树种, 截干栽植, 截干高度2.46—2.89 m。试验设无任何处理为对照(空白对照, CK)、填埋秸秆层(B)、覆盖秸秆层(M)和填埋配合覆盖秸秆层(B+M), 4种处理。选取干燥的玉米秸秆, 切碎为10 cm左右作秸秆层材料。根据前人对有关秸秆层的研究, 结合本试验速生杨根系深度、秸秆厚度以及当地地下水深度, 将填埋深度设为距离地表 80 cm处, 覆盖量和填埋量都设为7000 kg/hm2[6-7]。在地表覆盖秸秆后, 其表面铺设5 cm土, 起到固定秸秆的作用。把试验地规划分为4个区组, 每个区组规划分为3个小区, 把4种处理随机分配到4个区组, 每个小区1个处理, 每个处理3次重复。相邻区组之间的间隔为6 m;每个小区规格为9 m×9 m, 相邻小区之间的间隔为3 m。每小区栽植9棵, 株间行距3 m×3 m, 栽植穴的规格1 m×1 m×1 m。采用定额灌溉的方式, 考虑当地的气候因素、灌溉周期与农耕实际情况, 将灌溉额度定为200 mm, 在4月至10月进行灌溉, 每30天灌溉一次[17]。
1.3 试验测定内容及方法 1.3.1 光谱参数本试验利用Unispec-SC光谱仪(PPSYSTEM, USA)测量叶片反射光谱, 其波长测定范围310—1130 nm, 扫描波长间隔3.3 nm, 卤光灯的光强100%, 整合时间4 ms, 重复扫描次数为3次。为确保叶片采集的可靠性和普遍性, 选取有代表性顶部向阳处的成熟叶片, 每个小区随机选3棵速生杨, 每个处理共选9棵, 每棵采集3片叶片, 每片叶测3点不同位置。利用Multispec 5.1软件对光谱仪测量数据进行处理, 得到光谱反射率Rλ, 其中Rλ表示叶片反射与参比反射的比值, λ为波长。本研究选用了已被广泛证实较能反应叶片特征结构以及适合无损光学检测的光谱指数及公式(表 2)。
| 类型 Description |
反射光谱指数 Spectral reflectance indices |
缩写与公式 Acronym and Formulation |
文献来源 References |
| 归一化差值指数 Normalized difference indices |
叶绿素归一化指数 | 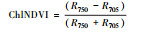
|
[18] |
| 改良红边归一化植被指数 | 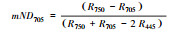
|
[19] | |
| 简单比率指数 Simple ratioindices |
改良归一化差值指数 | 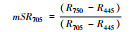
|
[19] |
| 其他 Others |
结构不敏感色素指数 | 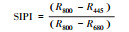
|
[20] |
| 植物衰老反射指数 | 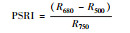
|
[21] | |
| 生理反射指数 | 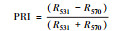
|
[22] | |
| Chl NDVI:叶绿素归一化指数, Chl normalized difference index;mND705:改良红边归一化植被指数, Modified red edge normalized difference vegetation index;mSR705:改良归一化差值指数, Modified red edge ratio;SIPI:结构不敏感色素指数, Structure-insensitive pigment index;PSRI:植物衰老反射指数, Plant senescence reflectance index;PRI:生理反射指数, Physiological reflectance index | |||
利用光谱归一化微分分析, 将反射光谱的数据进行一阶微分处理, 具体计算公式为:
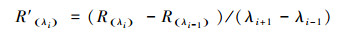
|
式中, λi是波段i的波长值, R(λi)是λi的光谱反射率值, R′(λi)是第i波段的一阶微分数值。并在680—750 nm范围内确定红边位置[14, 23]。
1.3.2 光合参数本试验利用LI-6400光合作用分析仪(Li-cor, USA), 选择天气晴朗的时期测定速生杨的光合特性。测定速生杨叶片的净光合速率(Pn, μmol m-2 s-1)、蒸腾速率(Tr, mmol m-2 s-1)、气孔导度(Gs, mmol m-2 s-1)、胞间CO2浓度(Ci, μmol/mol)和叶片水分利用效率(LWUE, μmol CO2/mmol H2O)等光合指标, 同时记录环境CO2浓度(Ca, μmolCO2/mol)、光合有效辐射(PAR, μmol m-2 s-1)等环境因子。
利用非直角双曲线模型对光响应过程模型拟合分析, 模型的具体表达式为:
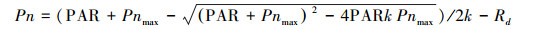
|
式中, Pn为净光合速率, 为表观量子效率(mol/mol);PAR为光合有效辐射(μmol m-2 s-1);Pnmax为最大净光合速率(μmol m-2 s-1);Rd为暗呼吸速率(μmol m-2 s-1)[24-25]。
1.3.3 土壤参数用管式不锈钢土钻采集土壤样品。土壤含水量的测定采用烘干法。土壤含盐量是采集的土壤经过风干后去杂质, 磨碎并过2 mm筛。以土水比1:5提取土壤溶液上清液, 用MP-522电导仪测电导率, 土壤含盐量如下计算公式[26]:
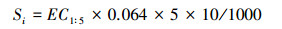
|
式中, Si表示第i土层含盐量(g/kg);EC1:5为土水比1:5的土壤浸提液电导率(dS/m)。
1.4 数据分析试验数据采用单因素方差分析(one-way ANOVA)和最小显著极差法(LSD), 在0.05水平上检验不同处理测定结果的差异显著性, 利用Origin 9.0作图。
2 结果与分析 2.1 光谱特征对秸秆层的响应 2.1.1 反射光谱特征图 1所示, 不同秸秆层的速生杨反射光谱曲线的整体趋势表现出一定的规律。可见光(400—760 nm)范围, 由于叶绿素对于绿色光反射作用极强, 而吸收蓝光与红光进行光合作用, 因此在绿光(550 nm)处有一小的反射峰, 蓝光(450 nm)和红光(670 nm)处有两个吸收谷。在700—800 nm存在反射的陡坡, 在1100 nm附近出现峰值。这是由于叶片内海绵组织对800—1300 nm的近红外波段强烈反射, 导致在光谱曲线上形成了最高反射峰。

|
| 图 1 不同秸秆层的速生杨反射光谱曲线和红光波段(680—750 nm)一阶导数曲线 Fig. 1 The spectral reflectance and first derivative in red area (680—750 nm) of fast-growing poplar leaves under different straw interlayers CK:空白对照, Blank control;B:填埋秸秆层, Bury straw interlayer;M:覆盖秸秆层, Straw mulching;B+M:填埋配合覆盖秸秆层, Straw interlayer plus straw mulching |
光谱噪声会对光谱微分技术造成影响, 采样时的光照、气候情况等都会导致测量的光谱曲线结果产生一定的差异, 通过微分变换, 将原始光谱数据进行一阶微分处理, 能够减少背景噪声的影响进而提高生化参数的监测效果[23]。红边位置是植物反射光谱中最明显的光谱特征之一, 是指在红光范围(680—750 nm)的光谱反射率的一阶微分值达到最大时所对应的波长, 由于红边特征不受叶片结构的影响, 因此适用于更广泛的植物检测, 红边位置与叶绿素的荧光具有很强的相关性, 叶绿素含量越高, 红光区域吸收越多, 红边就会向长波移动[18]。由图 1可知, 有秸秆层的B、M和B+M的在680—750 nm的反射光谱一阶导数最大值所对应的波长位置均在CK的长波方向处, 按各处理峰顶点所对应的波长位置大小顺序是:B+M>B>M>CK。
2.1.2 反射光谱指数不同秸秆层对土壤水盐运移及环境条件影响不同, 引发地上植物体内色素含量、营养状态以及生理生化状况发生相应变化, 导致速生杨叶片反射光谱指数发生变化(图 2)。叶绿素归一化指数(Chl NDVI)以及改良归一化差值指数(mSR705)与叶绿素的含量呈线性关系[18-19]。B、M和B+M的mSR705与Chl NDVI均显著大于CK(P < 0.05), 其中B最大, B+M次之。改良红边归一化植被指数(mND705)与叶绿素含量具有极高的相关性[19]。方差分析表明, B与B+M的mND705均大于其他处理(P < 0.05), 其中B+M最大, B其次。B和B+M两种处理都具有填埋的秸秆层, 表明填埋秸秆层可以显著提高速生杨叶片的叶绿素含量。结构无关色素指数(SIPI)是反映类胡萝卜素与叶绿素a比值的光谱指数[20], B、M和B+M的SIPI均比CK的高(P < 0.05), 其大小关系为B>M>B+M>CK。植物衰老反射指数(PSRI)是与类胡萝卜素和叶绿素比值高度相关的指数[21], 有秸秆的三种处理PSRI均小于CK, 且各处理间差异显著(P < 0.05), 大小关系排序为CK>B+M>M>B, 表明秸秆层可以显著改变地上速生杨叶片的类胡萝卜素含量。光化学反射指数(PRI)与叶黄素循环中的色素含量呈负相关, 当色素含量发生变化, 过多的辐射会被叶黄素循环中的色素吸收, 而无法进行光合作用, 所以当PRI低时, 其光合速率就低[22]。图 2中CK和M的PRI之间无显著差异(P>0.05), 而B与B+M显著高于其他处理(P < 0.05), 根据PRI结果所示, B与B+M比其他处理的地上速生杨光合速率水平要高。

|
| 图 2 不同秸秆层的速生杨叶片光谱指数变化 Fig. 2 Changes of the reflectance spectrum indices of fast-growing poplar leaves under different straw interlayers Chl NDVI:叶绿素归一化指数, Chl normalized difference index;mND705:改良红边归一化植被指数, Modified red edge normalized difference vegetation index;mSR705:改良归一化差值指数, Modified red edge ratio;SIPI:结构不敏感色素指数, Structure-insensitive pigment index;PSRI:植物衰老反射指数, Plant senescence reflectance index;PRI:生理反射指数, Physiological reflectance index |
速生杨的Pn、Tr、Gs、Ci以及LWUE等光合气体交换参数对不同秸秆层的响应是不同的, 并且在不同生长时期也是不同的(表 3)。随着时间的变化, 4种处理的Pn表现出逐渐降低的趋势。与对照组CK相比, B+M提高了不同时期速生杨的Pn, 其前期(5月)、中期(7月)和末期(9月)的Pn分别比CK高0.73%、28.07%和24.56%。B与M在中、后期的Pn与B+M表现出相同的变化趋势, 并且都显著高于CK, 但均低于B+M。秸秆层明显有助于提高速生杨在不同生长期的Pn, 其中, B+M的Pn提高的效果最好。从作用时期看, 各处理在中期对速生杨Pn提高的幅度最大。各处理Gs随着时间变化, 呈现降低的趋势, 总体上B、M与B+M的气孔导度均比CK大(P < 0.05)。各处理的Pn、Gs和Tr呈现相同的变化趋势, 原因是一方面, 由于速生杨的Tr受气孔大小调节, 因此Gs较高, Tr也较大, 因此4种处理速生杨的Tr变化趋势与Gs相同。另一方面, 速生杨会由于Pn的迅速下降, 通过降低Tr来提高LWUE。速生杨LWUE总体呈降低的趋势, 前期差异不明显, 中后期B、M、B+M的LWUE均比CK大(P < 0.05)。而速生杨的Ci随着时间的变化趋势与Pn、Gs和Tr不同, 各处理间在中期差异不显著, 前期和后期B+M均显著大于CK(P < 0.05)。
| 月份 Month |
处理 Treatments |
净光合速率(Pn) Photosynthetic rate/(μmol m-2 s-1) |
气孔导度(Gs) Stomata conductance/(mmol m-2 s-1) |
蒸腾速率(Tr) Transpiration rate/(mmol m-2 s-1) |
胞间CO2浓度(Ci) Intercellular CO2 concentration/(μmol/mol) |
水分利用效率(LWUE) Leaf water use efficiency/(μmol/mmol) |
| 5 | CK | 10.93±0.75a | 69.80±2.72a | 3.58±0.24a | 192.80±4.60c | 3.05±0.05b |
| B | 11.07±4.58a | 66.34±1.80a | 3.60±0.08a | 229.44±7.26a | 3.02±0.16b | |
| M | 10.88±10.36a | 65.61±1.42a | 3.49±0.31b | 182.81±6.27d | 3.20±0.31a | |
| B+M | 11.01±0.45a | 62.28±3.25b | 3.61±0.11a | 207.63±4.40b | 3.05±0.18b | |
| 7 | CK | 5.13±0.74c | 42.52±2.34b | 2.10±0.07c | 226.38±10.56a | 2.44±0.34b |
| B | 6.04±0.07b | 46.34±3.21a | 2.35±0.06b | 227.17±5.78a | 2.57±0.07a | |
| M | 6.22±0.18b | 47.83±1.90a | 2.44±0.10a | 223.03±3.79a | 2.56±0.18a | |
| B+M | 6.57±0.32a | 42.84±3.32b | 2.55±0.03a | 203.22±6.49b | 2.57±0.11a | |
| 9 | CK | 4.27±0.16c | 36.58±2.82c | 2.10±0.09b | 178.06±2.59c | 2.03±0.01b |
| B | 4.54±0.13b | 43.09±7.96b | 2.14±0.02a | 197.77±11.98b | 2.12±0.08b | |
| M | 4.66±0.28b | 41.13±1.57b | 2.16±0.01a | 217.57±3.02a | 2.15±0.13b | |
| B+M | 5.31±0.99a | 50.08±4.49a | 2.24±0.09a | 223.97±7.58a | 2.37±0.34a | |
| 同列具有不用字母表示处理间差异显著(P < 0.05); CK:空白对照, Blank control;B:填埋秸秆层, Bury straw interlayer;M:覆盖秸秆层, Straw mulching;B+M:填埋配合覆盖秸秆层, Straw interlayer plus straw mulching | ||||||
如图 3所示, 不同秸秆层的速生杨光合作用的光响应过程及其特征参数均发生了显著变化。不同处理的速生杨净光合速率(Pn)随着光合有效辐射(PAR)的升高, 整体上呈相似的变化趋势, 均先迅速增加, 然后逐渐变缓。在弱光条件下(PAR≤200 μmol m-2s-1), Pn随着PAR的增加呈线性升高, 表明PAR是光合作用的主导因子, 但4种处理间Pn对PAR的响应差异不明显(P>0.05)。随着PAR继续增加, 当PAR>200 μmol m-2 s-1, Pn上升至最大净光合速率(Pnmax), PAR增至饱和点(LSP), 约250 μmol m-2 s-1, 在此阶段Pn随着PAR增加呈曲线上升, 表明PAR不是该阶段的主导因子, 大气温湿度、Ca、Ci等生理生态因子也对速生杨的光合作用产生重要影响。当PAR超过LSP, 各处理Pn随着PAR增加而无明显变化, 光响应曲线趋于平缓。

|
| 图 3 不同秸秆层的速生杨叶片净光合速率的光响应及其模拟 Fig. 3 Photosynthetic rate-light response curves and simulation of fast-growing poplar under different straw interlayers |
如表 4所示, 空白对照组CK代表了自然状况下速生杨光响应参数值。方差分析表明, 与CK相比, 有秸秆层的三种处理的光响应参数呈显著差异。B、M和B+M的Pnmax与LSP都显著提高(P < 0.05), 其中B+M最高, B和M次之, CK最低。LCP和Rd差异水平结果相同, 处理M大于CK(P < 0.05), B与B+M均小于CK, 其中B最小。
| 处理 Treatments |
表观量子效率(Φ) Apparent quantum yield/(mol/mol) |
饱和光强(LSP) Light saturation point/(μmol m-2 s-1) |
最大净光合速率(Pnmax) Maximum photosynthetic rate/(μmol m-2 s-1) |
光补偿点(LCP) Light compensation point/(μmol m-2 s-1) |
暗呼吸速率(Rd) Respiration rate/(μmol m-2 s-1) |
| CK | 0.056±0.0029a | 243.83±15.66c | 12.80±1.27c | 9.81±1.14b | 0.557±0.049b |
| B | 0.056±0.0012a | 250.65±10.37bc | 13.68±1.69b | 5.53±0.49d | 0.314±0.034d |
| M | 0.055±0.0017a | 266.67±8.59b | 13.75±2.34b | 14.05±0.84a | 0.768±0.142a |
| B+M | 0.050±0.0011b | 306.75±7.31a | 14.88±2.29a | 8.96±0.64c | 0.447±0.083c |
图 4表示5月、7月和9月灌溉后30天内的土壤含水量变化。随着时间推进, 4种处理的土壤含水量均逐渐下降, 但下降程度具有差异。CK为空白处理, 其含水量下降程度高, 一个灌溉周期后的值最低;与CK对比发现, B、M和B+M均一定程度改变了土壤含水量的下降速度, 并提高灌溉后的土壤含水量, 其中处理B+M蓄水效果较好, 表明秸秆层均能够提高土壤含水量, 其中填埋秸秆层的蓄水效果最好。

|
| 图 4 不同秸秆层在不同时期土壤含水量变化 Fig. 4 Soil water content in the different stages under different straw interlayers |
由图 5可以看出, 随着时间推移, 不同处理条件, 土壤盐分含量总体呈降低趋势, 但降低程度具有明显差异。与空白对照组CK相比, B与B+M在不同时期的土壤盐分含量均低于其他处理(P < 0.05), 但处理B与B+M间无显著差异(P>0.05)。处理M也能够降低不同时期的盐分含量, 但盐分均高于处理B与B+M(P < 0.05)。表明秸秆层能够显著降低不同时期土壤盐分含量, 并且填埋秸秆层的抑盐效果要高于覆盖秸秆层。从不同的月份来看, 5月份各处理盐分含量相对较高, 原因是可能是5月份仅进行了一次灌溉, 未完全发挥出秸秆的淋洗作用, 随着时间的推进, 灌溉次数的增加, 各处理的盐分均有所下降, 其中有填埋秸秆层的处理B与B+M土壤盐分含量降低程度最大。

|
| 图 5 不同秸秆层在不同时期土壤盐分含量 Fig. 5 Soil salt content in the different stages under different straw interlayers |
光合作用是所有绿色植物中极为复杂和最基本的生理过程, 其机制涉及各种组分, 包括光合色素和光系统, 电子传递系统和二氧化碳减少途径, 因此任何由胁迫引起的损害都可能影响绿色植物的整体光合能力[10, 27]。同时, 光合作用是植物生产力高低的决定因素和对环境胁迫程度的反应指标[28]。本试验所在的西北干旱与半干旱地区, 降水少, 蒸发量大, 土壤水分大量无效蒸发以及土壤盐分积聚, 所导致的干旱胁迫和盐胁迫是限制本地区植被成活的重要原因[3-4]。Chaves等人研究表明盐胁迫会抑制或破坏植物光系统的部分功能, 电子传递系统以及光合作用的能量, 使得植物叶片中的叶绿素a、叶绿素b、类胡萝卜素以及酶活性等明显降低, 导致光合作用下降;水分胁迫通过气孔限制和叶肉细胞光合活性下降的非气孔限制两种机制引发植物光合效率降低[12]。在本试验中, 5月处于改良前期, 土壤盐分较高且差异不明显, 各处理的速生杨净光合速率、蒸腾速率和气孔导度等光合气体交换参数之间差异不明显, 与其他时期相比光合效率水平较低, 其原因主要是盐胁迫引起的。经过数月的灌溉和降水的淋洗作用, 在7月与9月, 与空白对照组相比, 地下填埋秸秆层的B与B+M土壤盐分下降到较低水平, B和B+M明显提升不同时期地上速生杨Pn、Tr、Gs, 表明填埋秸秆层对速生杨的光合作用效能及其光能利用效率产生了明显的提升作用。另外, 叶片水分利用效率(LWUE)的大小可以反映出植物对逆境的适应能力[29-30]。在前期, 各处理土壤水盐环境大致相同, 各处理的LWUE差异不明显;中后期, B+M的LWUE高于CK和其他处理, 表明填埋搭配覆盖秸秆层处理后的水盐环境更适合速生杨生长。这与王琳琳等对刺槐的研究结果相符[28]。最大净光合速率(Pnmax)体现了植物对强光的利用能力, 饱和光强(LSP)和光补偿点(LCP)表示对光能(强光和弱光)的利用能力和程度, 表观量子效率(Φ)表征了植物对弱光的利用能力, 暗呼吸速率(Rd)表示植物的线粒体呼吸[29]。B、M和B+M的最大净光合速率与饱和光强均显著提高, 表示具有秸秆层的处理提升了速生杨的有机物的合成能力以及强光的利用能力和程度。B和B+M其中光补偿点和暗呼吸速率明显降低, M则相反, 表示填埋秸秆层的处理提升了速生杨对弱光的利用能力和程度, 并降低植物的呼吸消耗。
植物的光谱特性是指植物受电磁波作用时, 会在某些特定的波段, 由于电子跃迁、分子振动和转动等复杂作用, 形成能够反映物质结构和组成的光谱吸收与反射特征, 这种对不同波长位置光谱的响应特性叫植物的光谱特性[14]。植物光谱特性与植物的色素含量、水分含量、以及营养状况密切相关, 通过光谱可以检测和分析植物的生长状况、光合能力及受胁迫程度等生理状况[12, 27, 31-32]。本试验中, 处理B+M的速生杨叶片光合色素含量最高, 光合特性以及营养状况最好, 红边位置以及其他光谱参数的结果与CK相比都有显著提升。而处理B与M也可以显著提升速生杨光合色素量和光合特性, 表明秸秆层对缓解灌区次生盐碱地速生杨生长所受胁迫具有积极的作用, 其中处理B+M的效果最好。光化学反射指数(PRI)与植物叶片光能利用效率呈正相关, 能够反映不同植物在不同营养条件和不同生长阶段的光能利用效率, 因为PRI与叶黄素循环中的色素含量呈负相关, 当色素含量发生变化, 过多的辐射会被叶黄素循环中的色素吸收, 而无法进行光合作用, 所以当PRI低时, 其光合速率就低[33-34]。本试验中, PRI所与光合特性所反应的结果较为一致, 进一步表明了秸秆层对提升灌区次生盐碱地植物光合效率具有积极的作用。另外, 本试验两种与叶片光合色素含量相关的光谱指数SIPI和PSRI所得出来的结果稍有差异, 这是由于反应叶片色素的特征光谱指数会因植被类型、生长阶段以及取样方案的不同而有所差异, 因此在选择利用光谱分析技术更准确的研究植被时, 要根据具体的研究对象和条件, 通过更进一步的研究筛选出最适宜该试验植物光谱特征的指数。
引黄灌区农业生产中会产生的大量秸秆废弃物, 探究利用秸秆层改良灌区次生盐碱地是一种重要、有效的秸秆利用方式, 也是一种合理、因地制宜的土壤改良方式。不仅能够避免造成环境污染和资源浪费, 还能增加土壤有机质和养分, 改善土壤水盐胁迫环境。本研究中, 土壤水盐变化的结果表明, 与对照组CK对比, B、M和B+M土壤含水量较高, B和B+M的盐分含量低, 由此可知, 在地表覆盖秸秆层的处理M虽然能够提高了土壤含水量, 但降低土壤盐分的作用较弱, 而栽植穴底部具有秸秆层的处理B和B+M有助于提高土壤水分, 有效降低土壤盐分, 起到控盐保水的效果, 这与赵永敢等人的研究结果相一致[35]。结合土壤的水盐的变化来看, 不同秸秆层对土壤水盐调控程度不同, 造成各处理的土壤环境条件不同, 影响地上速生杨生理状况发生相应变化, 填埋秸秆层为速生杨根层创造了较适宜的水盐土壤环境, 从而减轻了水盐胁迫对速生杨光谱与光合特性的抑制作用。
在以往对次生盐碱地土壤改良与植被恢复的研究当中, 一般较为注重土壤改良状况, 而在植被恢复方面, 对于农作物一般看其产量或生物量的多少, 而对于具有生态效益的非经济植物, 利用传统的化学分析法检测植物生理状况, 操作步骤繁琐, 花费时间较长, 还会对植物造成一定的损伤, 更不利于对同一植物的生长状况、光合效率和受胁迫等情况进行长期动态监测。而通过检测植物光谱与光合特性可以揭示出地上植物生长及生理状况的变化, 不仅迅速、准确、无损伤, 并且通过一次测定就可以分析出光合色素、生长状态以及受胁迫等生理状况, 基于此, 可以通过更进一步研究, 探索利用植物的光谱及光合特性评估各类盐碱地改良措施的植被恢复效益, 对各类改良措施的植被恢复状况进行更为高效且准确的评估。
4 结论速生杨的光谱及光合特性对不同秸秆层的响应是不同的, 秸秆层有助于提升灌区次生盐碱地中速生杨的光谱及光合等生理状况。B、M和B+M均能显著提升速生杨叶片的最大净光合速率与饱和光强, 其中, B和B+M还明显降低速生杨叶片的光补偿点和暗呼吸速率, M则不行。B和B+M均显著提高速生杨叶片的净光合速率、蒸腾速率、气孔导度等光合参数。另外, B+M的速生杨叶片光合色素含量最高, 营养状况最好, 并且其他光谱参数结果与CK相比都有显著提高。处理B和M也可以显著提高速生杨叶片光合色素含量和营养等生理状况, 但效果均不如B+M。
不同秸秆层对土壤的水盐调控程度不同, 秸秆层能够改善灌区次生盐碱地的高盐、干旱的土壤环境, 减轻盐胁迫与干旱胁迫对速生杨生理状况的抑制作用, 从而引起速生杨光谱与光合状况发生相应变化。B与B+M均能降低不同时期的土壤盐分含量, 且两处理间无显著差异, 然而, B+M在不同时期提高土壤含水量的效果均高于B。M也能降低土壤盐分和提升土壤含水量, 但不同时期的效果均不如处理B+M。
综合试验结果, 在引黄灌区次生盐碱地, 不同秸秆层均能不同程度的调节土壤水盐变化, 缓解速生杨所受水盐胁迫, 提升速生杨光谱与光合特性, 其中B+M秸秆层模式效果最佳。
| [1] |
Hatton T J, Nulsen R A. Towards achieving functional ecosystem mimicry with respect to water cycling in southern Australian agriculture. Agroforestry Systems, 1999, 45(1/3): 203-214. DOI:10.1023/A:1006215620243 |
| [2] |
Barrett-Lennard E G. Restoration of saline land through revegetation. Agricultural Water Management, 2002, 53(1/3): 213-226. |
| [3] |
马传明, 靳孟贵. 西北地区盐渍化土地开发中存在问题及防治对策. 水文, 2007, 27(1): 78-81. DOI:10.3969/j.issn.1000-0852.2007.01.021 |
| [4] |
杨劲松. 中国盐渍土研究的发展历程与展望. 土壤学报, 2008, 45(5): 837-845. DOI:10.3321/j.issn:0564-3929.2008.05.010 |
| [5] |
Jiang S C, He N P, Wu L, Zhou D W. Vegetation restoration of secondary bare saline-alkali patches in the Songnen plain, China. Applied Vegetation Science, 2010, 13(1): 47-55. DOI:10.1111/j.1654-109X.2009.01048.x |
| [6] |
李芙荣, 杨劲松, 吴亚坤, 姚荣江, 余世鹏, 张建兵. 不同秸秆埋深对苏北滩涂盐渍土水盐动态变化的影响. 土壤, 2013, 45(6): 1101-1107. |
| [7] |
张建锋. 盐碱地的生态修复研究. 水土保持研究, 2008, 15(4): 74-78. |
| [8] |
Van Gaalen K E, Flanagan L B, Peddle D R. Photosynthesis, chlorophyll fluorescence and spectral reflectance in Sphagnum moss at varying water contents. Oecologia, 2007, 153(1): 19-28. DOI:10.1007/s00442-007-0718-y |
| [9] |
Gamon J A, Surfus J S. Assessing leaf pigment content and activity with a reflectometer. New Phytologist, 1999, 143(1): 105-117. DOI:10.1046/j.1469-8137.1999.00424.x |
| [10] |
Ashraf M, Harris P J C. Photosynthesis under stressful environments:An overview. Photosynthetica, 2013, 51(2): 163-190. DOI:10.1007/s11099-013-0021-6 |
| [11] |
陈媛媛, 江波, 王效科, 李丽. 北京典型绿化树种幼苗光合特性对硬化地表的响应. 生态学报, 2017, 37(11): 3673-3682. |
| [12] |
Chaves M M, Flexas J, Pinheiro C. Photosynthesis under drought and salt stress:regulation mechanisms from whole plant to cell. Annals of Botany, 2009, 103(4): 551-560. DOI:10.1093/aob/mcn125 |
| [13] |
Naidu R A, Perry E M, Pierce F J, Mekuria T. The potential of spectral reflectance technique for the detection of Grapevine leafroll-associated virus-3 in two red-berried wine grape cultivars. Computers and Electronics in Agriculture, 2009, 66(1): 38-45. DOI:10.1016/j.compag.2008.11.007 |
| [14] |
薛忠财, 高辉远, 彭涛, 姚广. 光谱分析在植物生理生态研究中的应用. 植物生理学报, 2011, 47(4): 313-320. |
| [15] |
胡砚霞, 黄进良, 杜耘, 韩鹏鹏, 王久玲, 黄维. 洞庭湖洲滩速生杨树林变化信息提取方法. 生态学报, 2014, 34(24): 7243-7250. |
| [16] |
Jain S K, Singh P. Economic analysis of industrial agroforestry:poplar (Populus deltoides) in Uttar Pradesh (India). Agroforestry Systems, 2000, 49(3): 255-273. DOI:10.1023/A:1006388128069 |
| [17] |
孙兆军.银川平原盐碱荒地改良模式研究[D].北京: 北京林业大学, 2011. http://cdmd.cnki.com.cn/Article/CDMD-10022-1011132781.htm
|
| [18] |
Gitelson A, Merzlyak M N. Spectral reflectance changes associated with autumn senescence of Aesculus hippocastanum L. and Acer platanoides L. leaves. Spectral features and relation to chlorophyll estimation. Journal of Plant Physiology, 1994, 143(3): 286-292. DOI:10.1016/S0176-1617(11)81633-0 |
| [19] |
Sims D A, Gamon J A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sensing of Environment, 2002, 81(2/3): 337-354. |
| [20] |
Penuelas J, Baret F, Filella I. Semi-empirical indices to assess carotenoids/chlorophyll a ratio from leaf spectral reflectance. Photosynthetica, 1995, 31(2): 221-230. |
| [21] |
Merzlyak M N, Gitelson A A, Chivkunova O B, Rakitin V Y. Non-destructive optical detection of pigment changes during leaf senescence and fruit ripening. Physiologia Plantarum, 1999, 106(1): 135-141. |
| [22] |
Gamon J A, Peñuelas J, Field C B. A narrow-waveband spectral index that tracks diurnal changes in photosynthetic efficiency. Remote Sensing of Environment, 1992, 41(1): 35-44. DOI:10.1016/0034-4257(92)90059-S |
| [23] |
吴寿国, 余学军, 李凯, 蒋玉俭, 张汝民. 海盐对绿竹叶片反射光谱及叶绿素荧光参数的影响. 生态学报, 2014, 34(17): 4920-4930. |
| [24] |
孙宝玉, 韩广轩, 陈亮, 王安东, 吴立新, 赵敏. 短期模拟增温对黄河三角洲滨海湿地芦苇光响应特征的影响. 生态学报, 2018, 38(1): 167-176. |
| [25] |
叶子飘, 于强. 光合作用光响应模型的比较. 植物生态学报, 2008, 32(6): 1356-1361. DOI:10.3773/j.issn.1005-264x.2008.06.016 |
| [26] |
Pang H C, Li Y Y, Yang J S, Liang Y S, Yang J S. Effect of brackish water irrigation and straw mulching on soil salinity and crop yields under monsoonal climatic conditions. Agricultural Water Management, 2010, 97(12): 1971-1977. DOI:10.1016/j.agwat.2009.08.020 |
| [27] |
张金林, 李惠茹, 郭姝媛, 王锁民, 施华中, 韩庆庆, 包爱科, 马清. 高等植物适应盐逆境研究进展. 草业学报, 2015, 24(12): 220-236. DOI:10.11686/cyxb2015233 |
| [28] |
王琳琳, 李素艳, 孙向阳, 张涛, 付颖, 张红蕾. 不同隔盐措施对滨海盐碱地土壤水盐运移及刺槐光合特性的影响. 生态学报, 2015, 35(5): 1388-1398. |
| [29] |
谢东锋, 张光灿, 夏宣宣, 郎莹. 不同浓度酚酸对欧美杨I-107苗木生长和光合特性的影响. 生态学报, 2018, 38(5): 1789-1798. |
| [30] |
贾中民, 王力, 魏虹, 冯汉茹, 李昌晓. 垂柳和旱柳对镉的积累及生长光合响应比较分析. 林业科学, 2013, 49(11): 51-59. |
| [31] |
彭涛, 李鹏民, 贾裕娇, 高辉远. 介绍两种无损伤测定植物活体叶片色素含量的方法. 植物生理学通讯, 2006, 42(1): 83-86. |
| [32] |
刘伟国, 范秀华. 长白山红松阔叶林主要树种光谱反射特征对光环境的响应. 生态学报, 2009, 29(9): 4720-4726. DOI:10.3321/j.issn:1000-0933.2009.09.015 |
| [33] |
Demmig-Adams B, Adams Ⅲ W W. The role of xanthophyll cycle carotenoids in the protection of photosynthesis. Trends in Plant Science, 1996, 1(1): 21-26. |
| [34] |
Chaves M M, Maroco J P, Pereira J S. Understanding plant responses to drought-from genes to the whole plant. Functional Plant Biology, 2003, 30(3): 239-264. DOI:10.1071/FP02076 |
| [35] |
赵永敢, 李玉义, 胡小龙, 王婧, 逄焕成. 地膜覆盖结合秸秆深埋对土壤水盐动态影响的微区试验. 土壤学报, 2013, 50(6): 1129-1137. |