 2019, Vol. 39
2019, Vol. 39文章信息
- 邓健, 张丹, 张伟, 任成杰, 郝雯晖, 刘冲, 韩新辉, 杨改河
- DENG Jian, ZHANG Dan, ZHANG Wei, REN Chengjie, HAO Wenhui, LIU Chong, HAN Xinhui, YANG Gaihe
- 黄土丘陵区刺槐叶片-土壤-微生物碳氮磷化学计量学及其稳态性特征
- Carbon, nitrogen, and phosphorus stoichiometry and homeostasis characteristics of leaves, soil, and microbial biomass of Robinia pseudoacacia forests in the Loess Hilly Region of China
- 生态学报. 2019, 39(15): 5527-5535
- Acta Ecologica Sinica. 2019, 39(15): 5527-5535
- http://dx.doi.org/10.5846/stxb201808281845
-
文章历史
- 收稿日期: 2018-08-28
- 网络出版日期: 2019-05-16





2. 西北农林科技大学农学院, 杨凌 712100
2. College of agronomy, Northwest Agriculture and Forestry University, Yangling 712100, China
C、N、P是植物生长的必须元素, 其循环和耦合关系对维持植物个体良好生长和生态系统可持续性具有重要意义[1]。生态化学计量学为研究从个体到生态系统不同层次的能量平衡和多重化学元素平衡提供了新思路, 并成为生物稳态性和养分元素限制研究的新方法[2-4]。植物、微生物等生物体为了应对土壤养分限制等外界条件变化对自身生长的影响, 通过对体内的化学元素浓度及不同化学元素间的计量比例进行调节, 从而维持内部化学组成相对稳定, 即内稳态调节[5]。生物的内稳态性是生态化学计量学理论的基础和核心, 能够综合反映物种对生存环境变化的适应性[2]。阐明生态系统土壤-植物-微生物生态化学计量学特征及其内稳态性对于了解生态系统结构、功能和稳定性具有重要意义[6]。
目前, 国内外学者对陆地生态系统植物、土壤和微生物C、N、P生态化学计量学特征进行了大量研究, 尤其是生态化学计量学在限制性养分元素判别方面的应用形成了“限制元素稳定性假说”[7]、植物N:P养分限制理论等一系列成果[8];土壤微生物生物量C:N:P计量比也被广泛用于指示植被恢复过程中土壤质量和养分平衡特征[9]。然而, 由于生物生存环境、养分供应状况、生长发育阶段等外界变化均会对植物和微生物养分元素计量比产生影响[10-12], 更多学者开始关注陆地生态系统生物体的内稳态调节。相关研究主要集中在草原植物内稳态特征、氮磷添加和盐胁迫等处理对特定植物内稳性的影响、土壤微生物内稳态性与土壤中C、P元素的关系等方面[13-16]。这些研究对丰富生态化学计量学和内稳态调节理论有重要意义, 但大多数研究主要关注某一组分(土壤、植物或者微生物)的生态化学计量学特征, 将“植物-土壤-微生物”连续体作为系统研究内部C、N、P计量学和内稳态特征及其相互关系的研究还比较少。土壤为植物和微生物生长提供了基本环境和营养来源, 而植物生长和微生物活动为土壤提供了养分来源和转换驱动力, 而且随植被生长年限的变化, 植物、微生物与土壤之间的养分循环关系也会发生改变[17]。因此, 运用生态化学计量学方法, 将植物-土壤-微生物作为连续体, 弄清生态系统植物和微生物元素计量比在植被生长过程中随土壤C、N、P含量及其化学计量比变化的稳态性特征, 对于阐明森林生态系统元素循环和制约变化规律, 丰富陆地生态系统生态化学计量学理论具有重要意义[2]。
黄土高原地区是我国生态环境最为脆弱的地区之一, 开展植被恢复是该区域减少水土流失、改善生态环境的重要手段[18]。植被恢复在增加植被覆盖的同时也影响了该区域生态系统养分循环过程及其平衡特征, 如随着恢复年限增加土壤C库不断积累, 但也可能导致植物受N或P元素限制等问题[19]。由于不同的植被恢复方式和恢复阶段的生态系统养分平衡存在较大差异[4, 20], 导致目前黄土高原典型人工林植物-土壤-微生物系统养分循环耦合关系及其稳定机制尚不明确。刺槐(Robinia pseudoacacia)是黄土高原地区主要的人工造林树种, 不同阶段栽植的刺槐林发挥着巨大的水土保持、土壤改良等生态效益。因此, 本研究选取黄土丘陵沟壑区栽植5年、10年、20年、30年和45年的人工刺槐林为对象, 在分析刺槐林叶片-土壤-微生物化学计量学特征的基础上, 重点分析不同恢复年限叶片和微生物随土壤养分变化的内稳态特征, 研究有助于更深入认识脆弱生境地区恢复植被系统养分循环规律和系统稳定性机制, 为黄土高原植被恢复和经营管理提供科学依据。
1 材料和方法 1.1 研究区概况研究区位于陕西省延安市安塞区五里湾流域内(109°19′—109°22′ E, 36°51′—36°52′ N), 海拔1010—1400 m。属于典型的黄土丘陵沟壑区, 土壤为黄土母质发育形成的黄绵土, 抗侵蚀能力差, 水土流失严重;气候类型为暖温带半干旱气候, 年平均降水量535 mm, 降水主要集中在6—9月, 年平均气温8.8℃, 无霜期159 d。该流域自20世纪70年代开始进行人工造林, 形成了不同恢复年限的人工植被群落。主要的造林树种有刺槐、柠条(Caragana korshinskii)、山杏(Armeniaca sibirica)等, 主要草本植物有铁杆蒿(Artemisia gmelinii)、草木犀(Melilotus officinalis)、长芒草(Stipa bungeana)、阿尔泰狗娃花(Heteropappus altaicus (Willd) Novopokr)、狗尾草(Setaria viridis)等。
1.2 试验设计和采样试验于2016年8月中旬开展, 试验选取立地条件相似、干扰程度较低、分别种植5年、10年、20年、30年和45年的刺槐林(编号为RP5、RP10、RP20、RP30和RP45, 林龄通过当地林业部门造林记录和走访农户确定)进行研究(表 1), 各年限刺槐林在造林前均为耕地, 主要种植作物为谷子(Setaria italica)、糜子(Panicum miliaceum)等。每个年限刺槐林选择3块立地条件相近的重复样地, 每个样地内设置大小为20 m×20 m的标准样方, 每个样方内随机选择5个1 m×1 m的小样方调查草本植被。每个20 m×20 m的样方内用梅花采样法选取5个采样点, 去除表面枯落物和植物残体后, 用土钻(直径为4 cm)采集距离树干80 cm以外的0—10 cm土壤。将同一样方内5个样点的土样混匀后分成两份, 一份过2 mm土筛后在4℃冷藏保存用于测定微生物生物量养分;一份在阴凉处风干保存用于测定土壤养分。每个20 m×20 m的样方内随机选择10株刺槐作为采样树, 分别在4个方向的树冠不同部位采集叶片, 将同一样方内的叶片混合后用四分法取部分装入牛皮纸袋, 带回实验室用水洗去灰尘, 表面风干后置60℃烘干, 用粉碎机粉碎后测定叶片养分。
| 样地Sample land | RP05 | RP10 | RP20 | RP30 | RP45 |
| 坐标Location | 36°52′05″N 109°20′41″E |
36°52′00″N 109°21′38″E |
36°52′08″N 109°21′06″E |
36°51′36″N 109°21′06″E |
36°52′16″N 109°20′54″E |
| 海拔Elevation/m | 1239.12 | 1332.31 | 1245.2 | 1283.6 | 1121.34 |
| 坡度Slope/(°) | 35 | 37 | 30 | 35 | 28 |
| 坡向Aspect/(°) | EN30 | ES38 | ES20 | EN16 | ES10 |
| 坡位Slope position | 上坡位 | 上坡位 | 上坡位 | 上坡位 | 上坡位 |
| 容重Bulk density/(g/cm3) | 1.04 | 1.09 | 1.29 | 1.05 | 0.98 |
| 刺槐郁闭度Canopy density/% | 30 | 48 | 55 | 64 | 60 |
| 刺槐密度Stand density/(株/hm2) | 1660 | 1540 | 1535 | 1350 | 1240 |
| 林下草本盖度Herb cover degree/% | 56.07 | 46.83 | 59.13 | 83.87 | 73.40 |
| 物种丰富度Species richness index | 18 | 13 | 24 | 21 | 23 |
| 林下主要植物 Mainly vegetation type |
狗尾草 Setaria viridis、 长芒草 Stipa bungeana |
草地老鹳草 Geranium pratense、 翻白委陵草 Potentilla discolor |
铁杆蒿 Artemisia gmelinii、 达乌里胡枝子 Lespedeza davurica |
铁杆蒿 Artemisia gmelinii |
铁杆蒿 Artemisia gmelinii、 悬钩子 Rubus corchorifolius |
| RP, 刺槐, Robinia pseudoacacia; 数字表示林龄Figure indicates the stand age; EN, 东偏北, East to north; ES, 东偏南, East to south | |||||
土壤和植物叶片有机碳含量均采用重铬酸钾-外加热容量法测定;土壤和植物叶片全氮分别采用浓硫酸-高氯酸和浓硫酸-H2O2消煮后用全自动凯氏定氮仪(KDY-9830)测定, 全磷经上述消煮后用钼酸铵比色法测定;采样时用环刀法测定土壤容重[21]。土壤微生物生物量C采用氯仿熏蒸-K2SO4溶液浸提后用总有机碳分析仪(TOC, 日本岛津)测定, 微生物量N采用氯仿熏蒸-K2SO4溶液浸提后用全自动凯氏定氮仪测定;微生物生物量P采用氯仿熏蒸-NaHCO3溶液浸提后用钼锑抗显色法测定[22-23]。
1.4 数据处理和统计分析内稳态指数(Homeostasis index, H)采用如下公式进行计算[5]:
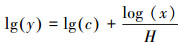
|
式中, H为内稳态指数, 因变量y为叶片或微生物生物量C、N、P含量或计量比, 自变量x为对应的土壤C、N、P含量或计量比, c为积分常数。按照Persson等的分类, 当方程拟合显著时, 将H>4、2<H<4、1.33<H<2、H<1.33分别划为稳态、弱稳态、弱敏感态和敏感态;但当方程拟合不显著时认为是绝对稳态[5]。
物种丰富度指数(R)=S, S为样方内物种数量[24]。
生态化学计量比用摩尔比表示;采用单因素方差分析(One-way ANOVA)检验各指标不同处理间的差异, 用Duncan法进行多重比较, 显著性水平为P<0.05。采用Pearson相关性分析土壤养分指标与叶片、微生物生物量C、N、P化学计量学指标间的相关性。数据整理和计算采用Microsoft Excel 2010, 使用SAS 9.0进行统计分析, 使用Origin 2016作图。
2 结果 2.1 不同年限刺槐林叶片-土壤-微生物C、N、P含量变化特征植物叶片C、N、P含量总体表现为随恢复年限的增加先增加后减小的趋势, 且均在恢复30年时达到最高(P<0.05)(表 2), 其中C、N、P含量分别比恢复5年高28.99%、96.59%和7.66%;但30年到45年间C、N、P含量分别下降了9.80%、10.33%和20.95%(P<0.05)。土壤C、N含量均表现出随恢复年限显著增加的趋势(P<0.05, 表 2), 从恢复5年到45年C、N分别增加了260.57%和302.41%;土壤P含量变化不显著(P>0.05)。土壤微生物生物量C、N和P含量均随恢复年限的增加显著增加(P<0.05, 表 2), 其中微生物C从恢复5年到30年增加了355.68%, 但从30年到45年下降了6.00%;微生物N和P从恢复5年到45年分别增加了175.24%和249.40%。
| 样品Samples | 样地Sample land | 碳含量C content | 氮含量N content | 磷含量P content |
| RP05 | 362.28±7.46c | 16.29±1.00e | 2.02±0.14a | |
| RP10 | 380.81±12.89c | 22.38±0.94d | 2.07±0.06a | |
| 叶片Leaf/(g/kg) | RP20 | 439.85±13.32b | 25.05±0.54c | 2.15±0.04a |
| RP30 | 467.31±8.51a | 32.02±0.75a | 2.17±0.11a | |
| RP45 | 421.53±12.06b | 28.71±0.8b | 1.72±0.06b | |
| RP05 | 4.00±0.05c | 0.30±0.09d | 0.52±0.04a | |
| RP10 | 4.65±0.27c | 0.50±0.03c | 0.55±0.07a | |
| 土壤Soil/(g/kg) | RP20 | 4.97±0.09c | 0.61±0.05c | 0.49±0.04a |
| RP30 | 8.26±0.71b | 0.87±0.07b | 0.60±0.04a | |
| RP45 | 14.41±0.91a | 1.20±0.13a | 0.57±0.03a | |
| RP05 | 98.54±2.63d | 19.75±1.67d | 4.28±0.09d | |
| RP10 | 102.36±9.42d | 21.48±2.16d | 6.70±0.57cd | |
| 微生物 | RP20 | 216.97±10.35c | 29.09±1.24c | 8.41±0.87bc |
| Microbes/(mg/kg) | RP30 | 449.02±9.41b | 48.78±2.83b | 10.56±0.89b |
| RP45 | 422.06±13.67a | 54.36±1.61a | 14.95±2.26a | |
| 同列数据后具有不同字母表示不同年限数据间差异显著(P<0.05);数据格式为平均值±标准误 | ||||
不同恢复年限叶片、土壤、微生物生物量C:N分别为17.03—26.03、9.55—16.94、5.57—10.76, 总体表现为叶片C:N>土壤C:N>微生物生物量C:N(图 1)。其中叶片C:N随恢复年限的增加先降低后稳定, 恢复30年最低, 比恢复5年下降了34.58%。土壤C:N随恢复年限增加先降低后增加, 从恢复5年到20年降低了43.61%, 从恢复20年到45年增加了47.81%。微生物生物量C:N从恢复5年到30年之间显著增加了84.01%(P<0.05), 在30年到45年之间下降了15.84%(P<0.05)。不同年限C:N变异系数表现为微生物生物量(24.90%)>土壤(21.36%)>叶片(16.29%)。

|
| 图 1 不同恢复阶段叶片、土壤、微生物C:N:P化学计量比 Fig. 1 C:N:P stoichiometry of leaf, soil and microbe in different restoration years 不同的小写字母表示不同恢复年限之间差异显著(P<0.05) |
不同恢复年限叶片、土壤、微生物生物量C:P分别为465.04—634.48、19.89—65.81和39.64—110.53, 各恢复年限均表现为叶片C:P>微生物生物量C:P>土壤C:P(图 1)。叶片和土壤中C:P均随恢复年限增加而增加, 从恢复5年到45年分别增加了36.43%和230.84%(P<0.05)。微生物生物量C:P随恢复年限的增加呈现波动变化, 从恢复10年到30年逐渐增加(P<0.05), 但随后出现下降。不同年限C:P变异系数表现为土壤(49.30%)>微生物生物量(33.01%)>叶片(11.51%)。
不同恢复年限叶片、土壤、微生物生物量N:P分别为17.89—37.03、1.24—4.68和7.15—10.26, 总体表现为叶片N:P>微生物生物量N:P >土壤N:P(图 1)。叶片中N:P和土壤N:P均随恢复年限增加而增加, 从恢复5年到45年分别增加了106.92%和275.98%(P<0.05)。微生物生物量N:P随着恢复年限的增加呈现波动变化, 从恢复10年到30年显著增加了43.43%(P<0.05), 但随后出现下降。不同年限N:P变异系数表现为土壤(41.27%)>叶片(24.41%)>微生物生物量(14.61%)。
2.3 土壤养分与叶片、微生物生物量C:N:P化学计量学的关系土壤C、N、P含量和计量学特征与叶片、微生物生物量C:N:P化学计量学特征的关系分析结果(表 3)显示, 土壤C、N含量与除微生物N:P外的其他计量学指标均具有显著的相关关系(P<0.05), 除与叶片C:N为负相关关系外, 与其他指标均为正相关关系;土壤P与叶片和微生物C:N:P计量学特征相关性不显著。土壤C:N仅与叶片C:N正相关, 与其他指标关系不显著;土壤C:P、N:P与除微生物C:P和N:P外的其他指标均显著相关(P<0.05)。
| 指标 Indicator |
叶片Leaf | 微生物Microbe | |||||
| C:Nleaf | C:Pleaf | N:Pleaf | C:Nmicrobe | C:Pmiccrobe | N:Pmiccrobe | ||
| Csoil | -0.672** | 0.902** | 0.899** | 0.559* | 0.404 | 0.008 | |
| Nsoil | -0.832** | 0.895** | 0.960** | 0.692** | 0.526* | 0.017 | |
| Psoil | -0.472 | 0.255 | 0.425 | 0.245 | 0.353 | 0.299 | |
| C:Nsoil | 0.556* | 0.004 | -0.220 | -0.264 | -0.127 | 0.225 | |
| C:Psoil | -0.637* | 0.904** | 0.877** | 0.545* | 0.353 | -0.062 | |
| N:Psoil | -0.813** | 0.902** | 0.942** | 0.693** | 0.474 | -0.081 | |
| * P<0.05; ** P<0.01 | |||||||
对恢复过程中叶片和微生物生物量C、N、P养分和化学计量学比随土壤变化的内稳态特征分析结果表明(表 4), 叶片P、C:N和微生物生物量P、C:N、N:P利用稳态性模型模拟结果不显著, 属于绝对稳态型;其他指标中, 叶片C内稳态指数为9.001(>4), 属于稳态型指标;叶片C:P和N内稳态指数较低, 属于弱稳态型指标, 而N:P内稳态指数较最低(1.949), 属于弱敏感型指标。微生物生物量C(0.835)和N(1.317)内稳态指数都小于1.33, 属于敏感型指标;微生物生物量C:P稳态性指数为2.585, 属于弱稳态类型。综合来看, 叶片的养分和计量学指标的内稳态性高于微生物。
| 类别Type | 变量Variable | H | R2 | P | 等级Grade | |
| x | y | |||||
| Csoil | Cleaf | 9.001 | 0.295 | <0.100 | 稳态 | |
| Nsoil | Nleaf | 2.329 | 0.849 | <0.100 | 弱稳态 | |
| 叶片Leaf | Psoil | Pleaf | 8.123 | 0.019 | 0.622 | 绝对稳态 |
| C:Nsoil | C:Nleaf | 3.542 | 0.180 | 0.115 | 绝对稳态 | |
| C:Psoil | C:Pleaf | 3.920 | 0.839 | <0.100 | 弱稳态 | |
| N:Psoil | N:Pleaf | 1.949 | 0.880 | <0.100 | 弱敏感 | |
| Csoil | Cmicrobe | 0.835 | 0.742 | <0.100 | 敏感 | |
| Nsoil | Nmicrobe | 1.317 | 0.838 | <0.100 | 敏感 | |
| 微生物生物量 | Psoil | Pmicrobe | 0.796 | 0.100 | 0.252 | 绝对稳态 |
| Microbe biomass | C:Nsoil | C:Nmicrobe | 3.975 | 0.051 | 0.417 | 绝对稳态 |
| C:Psoil | C:Pmicrobe | 2.585 | 0.234 | <0.100 | 弱稳态 | |
| N:Psoil | N:Pmicrobe | 17.241 | 0.020 | 0.618 | 绝对稳态 | |
| H, 内稳态指数, Homeostasis index | ||||||
植物-土壤-微生物系统中C、N、P的积累和转化是陆地生态系统元素循环的基础和核心环节[25]。植被恢复会通过增加有机物归还、改变土壤理化环境等途径影响土壤养分[26-27]。本研究中土壤C、N含量和微生物生物量C、N、P含量均随着刺槐恢复年限的增加而增加, 这与现有研究结果一致[28], 说明植被恢复促进了土壤养分积累和微生物养分转化;但土壤P含量没有发生显著变化。随着恢复年限和土壤条件的变化, 植物叶片中的C、N、P含量及其平衡状态也会发生改变, 结果显示不同年限刺槐叶片C、N、P含量平均值分别为414.36、24.89 g/kg和2.03 g/kg, 其中叶片C含量低于全球陆生植物叶片碳平均含量464 g/kg[29];变化趋势随恢复年限增加先增加后降低, 这与马任甜等发现的黄土高原刺槐林不同林龄间C含量表现为中龄林>成熟林>幼龄林的趋势基本一致[30], 主要可能是生长中后期叶片富含C的结构性物质积累导致C含量上升[30-31]。各恢复阶段叶片N含量均明显高于我国植物叶片平均N含量(18.60 g/kg)[32]和黄土高原刺槐叶片平均N含量(21.61 g/kg)[33], 而叶片P含量接近甚至低于全球叶片P含量(2.00 g/kg)[29]和黄土高原刺槐叶片平均P含量(2.09 g/kg)[33], 总体表现为氮含量丰富, 而磷相对缺乏, 尤其是生长30年以后更加明显, 这与此前的相关研究结果一致[34-35]。
植物、土壤、微生物中的C:N:P计量学特征能够更好地反应恢复生态系统的养分限制和平衡状态[36]。土壤养分库是植物和微生物吸收养分的主要来源, 本研究中土壤C:N随恢复年限增加先下降后上升, 主要可能是恢复前期刺槐根系固氮促使土壤氮素含量快速增加, 而后期凋落物增加引起有机碳大量输入[37];C:P和N:P均随恢复年限不断增加, 主要是由于土壤C、N的持续增加而P没有显著变化导致[30]。土壤微生物生物量C:N、C:P和N:P均随着恢复年限的增加呈波动变化, 主要是由于植被恢复过程中微生物群落可通过自身结构和代谢进行C:N:P计量学特征的调节[38]。叶片C:N:P计量学特征变化能够反映植物体从土壤中获取养分和利用养分的能力, 本研究中刺槐叶片C:N平均为20.11, 明显高于黄土高原刺槐叶片C:N平均水平(16.27)[30], 但低于全球叶片C:N平均值(22.5)[29]。C:N随恢复年限增加而不断降低, 说明叶片中N的吸收和积累速率要快于C。叶片C:P和N:P均随着恢复年限的增加而增加, 表明刺槐林地生长过程对P元素的吸收利用效率不断增加;恢复45年的刺槐叶片C:P和N:P分别为634.48和37.03, 明显高于全球叶片C:P(595)[31]和全国植物叶片平均N:P(32)[32];而现有研究表明植物叶片N:P>35(转换质量比为16)可能导致植物生长受到P限制[8]。植物和微生物吸收的P元素主要来自于土壤, 受土壤中P元素含量和有效性限制较大[19, 33];而土壤中可供植物和微生物吸收的有效P元素主要来自磷酸盐分解, 植物根系分泌物(如有机酸)和微生物胞外酶(如碱性磷酸酶)会促进土壤P元素向活性态无机磷转化以利于吸收利用[39]。随植被恢复年限的增加, 植物生物量增加储存了大量从土壤吸收的P元素抵消了外界向土壤输入的P元素(如磷沉降), 从而导致土壤P含量的变化不显著。同时, 随生长年限的增加植物很容易从大气和土壤吸收固定C、N, 但土壤可供植物吸收的有效P元素相对不足, 从而造成了植物叶片C:P和N:P的升高, 这在此前多项黄土高原地区的研究中均被提出[30, 35], 结合土壤养分的不协调变化, 说明该区域植被恢复可能面临着生态系统养分失衡的问题, P元素可能成为黄土丘陵区刺槐生长后期的限制养分。
3.2 植物和微生物生物量对土壤养分变化的响应及其稳态性土壤是植物生长和微生物活动的重要基础条件, 植物和微生物吸收的养分及其生态化学计量特征受到土壤养分条件的强烈影响[36]。通过分析土壤养分指标与刺槐叶片和土壤微生物生物量C:N:P化学计量学特征的关系发现, 土壤C、N含量与除微生物N:P外的其他计量学指标显著相关(P<0.05), 而土壤C:P、N:P也与除微生物C:P和N:P外的其他指标显著相关(P<0.05), 说明在植被恢复过程中土壤养分与植物和微生物养分计量学特征存在较强的耦合关系, 其中又以植物与土壤的关系更加紧密。
在面对土壤养分改变等外界环境因子变化时, 植物和微生物通过内稳态调节来保持体内化学构成的稳定[2, 16], 生物对体内元素的内稳态调节是生态化学计量学中的重要参数。本研究中刺槐叶片的N:P内稳态指数仅为1.949, 属于弱敏感型指标, 远低于羊草及其他草原维管束植物地上部分的内稳态指数(分别为4.61—5.61和3.08—14.09)[14, 16], 表明刺槐叶片N:P容易受到土壤养分条件的影响而发生变化, 进一步支持了植物叶片N:P可以作为判定植物养分平衡状态的重要指标[8], 说明植物N和P的综合管理对生态系统过程具有重要意义[6]。刺槐叶片C、N和C:P相对于N:P的稳态性指数高, 属于稳态或弱稳态指标, 其中C主要通过光和作用合成调节, 而N主要通过根瘤菌固氮和根系吸收N素进行调节, 在面对土壤养分变化时植物能够调节这些指标在体内保持相对稳定状态, 说明刺槐在研究区域具有较好的适应能力。土壤微生物在应对代谢底物养分条件变化时会通过自身化学计量学比、胞外酶、群落结构和代谢过程等多种途径进行调节, 因此能够维持微生物生物量C:N:P计量学特征的基本稳定[38], 本研究结果也证实了微生物生物量C:N、C:P和N:P均具有相对较高的内稳态性。随植被恢复年限的增加, 土壤微生物生物量和多样性均会伴随着代谢底物中C和N积累而显著增加[40], 而C和N作为微生物体的基本结构性元素, 也会随微生物群落的变化而变化, 由此导致了微生物生物量C、N稳态性指数均较低(0.835和1.317), 属于敏感型指标。
本研究中叶片P、C:N和微生物生物量P、C:N、N:P利用内稳态模型模拟结果不显著, 拟合优度都比较低(R2∈[0.019, 0.180]), 且这些指标的内稳态指数差异大(H∈[0.796, 17.241]), 这在相关研究中也曾遇到[5], 庾强等认为由于其检验不显著说明这些指标伴随着土壤养分变化的趋势不显著, 说明这些指标具有绝对稳定性[6, 16]。如由于土壤P相对C、N处于缺乏状态, 植物和微生物通过分泌磷酸酶等途径获取自身所需的P元素[38], 由此导致随着恢复年限增加, 土壤P变化不显著但植物和微生物P不断积累, 因而二者的关系不能被内稳态模型很好地解释;而植物C和N可以从大气固定, 因而对土壤养分的依赖程度也较低, 造成C:N随土壤C:N变化不显著。然而需要注意的是, 理想情况下分析误差是模型中指标变异的唯一来源, 可以将回归模拟不显著(P>0.05)的指标归为绝对稳定[41], 但这种归类忽略了微生物或植物某一指标本身在环境中是高度可变的情形, 或者其与土壤指标之间不具有事实上的必然联系, 这种情况下就无法期望通过内稳态模型验证指标的内稳态性。因此, 也有研究认为拟合优度较低的指标内稳态指数不具有统计学意义, 需要结合长期观测和控制实验才能进一步揭示其稳态性特征[5, 14]。
4 结论(1) 除土壤P含量外, 叶片、土壤和微生物生物量C、N、P含量及其计量学特征其他指标均随着恢复年限增加而发生显著变化, 总体表现为增加趋势;人工刺槐林生长后期可能面临P限制。
(2) 土壤C、N、C:P、N:P与大部分叶片和微生物生物量计量学指标关系紧密, 说明在植被恢复过程中其对植物和微生物生长具有重要作用。
(3) 叶片和微生物生物量C、N、P及其计量学特征中大部分指标与土壤的关系能够被内稳态模型很好地模拟(P<0.01);其中叶片N:P、微生物C、N对土壤养分变化较为敏感;其他指标比较稳定, 说明植物和微生物均会通过内稳态调节来改变自身元素构成以适应土壤养分变化, 刺槐在黄土丘陵区具有较好的适应性;微生物的内稳态指数低于刺槐, 对土壤养分的变化更加敏感。
致谢: 中国科学院水利部水土保持研究所安塞水土保持综合试验站为本研究野外采样提供帮助, 特此致谢。| [1] |
Hessen D O, Elser J J. Elements of ecology and evolution. Oikos, 2005, 109(1): 3-5. DOI:10.1111/j.0030-1299.2005.14055.x |
| [2] |
Sterner R W, Elser J J. Ecological Stoichiometry:The Biology of Elements from Molecules to the Biosphere. Princeton, New Jersey: Princeton University Press, 2002.
|
| [3] |
Zechmeister-Boltenstern S, Keiblinger K M, Mooshammer M, Peñuelas J, Richter A, Sardans J, Wanek W. The application of ecological stoichiometry to plant-microbial-soil organic matter transformations. Ecological Monographs, 2015, 85(2): 133-155. DOI:10.1890/14-0777.1 |
| [4] |
李鑫, 曾全超, 安韶山, 董扬红, 李娅芸. 黄土高原纸坊沟流域不同植物叶片及枯落物的生态化学计量学特征研究. 环境科学, 2015, 36(3): 1084-1091. |
| [5] |
Persson J, Fink P, Goto A, Hood J M, Jonas J, Kato S. To be or not to be what you eat:regulation of stoichiometric homeostasis among autotrophs and heterotrophs. Oikos, 2010, 119(5): 741-751. DOI:10.1111/j.1600-0706.2009.18545.x |
| [6] |
Yu Q, Chen Q S, Elser J J, He N P, Wu H H, Zhang G M, Wu J G, Bai Y F, Han X G. Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability. Ecology Letters, 2010, 13(11): 1390-1399. DOI:10.1111/j.1461-0248.2010.01532.x |
| [7] |
Han W X, Fang J Y, Reich P B, Woodward F I, Wang Z H. Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in China. Ecology Letters, 2011, 14(8): 788-796. DOI:10.1111/j.1461-0248.2011.01641.x |
| [8] |
Tessier J T, Raynal D J. Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation. Journal of Applied Ecology, 2003, 40(3): 523-534. DOI:10.1046/j.1365-2664.2003.00820.x |
| [9] |
吴建平, 韩新辉, 许亚东, 任成杰, 杨改河, 任广鑫. 黄土丘陵区不同植被类型下土壤与微生物C, N, P化学计量特征研究. 草地学报, 2016, 24(4): 783-792. |
| [10] |
Spohn M. Element cycling as driven by stoichiometric homeostasis of soil microorganisms. Basic and Applied Ecology, 2016, 17(6): 471-478. DOI:10.1016/j.baae.2016.05.003 |
| [11] |
Fanin N, Fromin N, Buatois B, Hättenschwiler S. An experimental test of the hypothesis of non-homeostatic consumer stoichiometry in a plant litter-microbe system. Ecology Letters, 2013, 16(6): 764-772. DOI:10.1111/ele.12108 |
| [12] |
许淼平, 任成杰, 张伟, 陈正兴, 付淑月, 刘伟超, 杨改河, 韩新辉. 土壤微生物生物量碳氮磷与土壤酶化学计量对气候变化的响应机制. 应用生态学报, 2018, 29(7): 2445-2454. |
| [13] |
Wang H, Li H Y, Zhang Z J, Muehlbauer J D, He Q, Xu X H, Yue C L, Jiang D Q. Linking stoichiometric homeostasis of microorganisms with soil phosphorus dynamics in wetlands subjected to microcosm warming. PLoS One, 2014, 9(1): e85575. DOI:10.1371/journal.pone.0085575 |
| [14] |
李晴宇.盐碱胁迫条件下羊草的生态化学计量内稳性研究[D].长春: 吉林大学, 2017. http://cdmd.cnki.com.cn/Article/CDMD-10183-1017156353.htm
|
| [15] |
陶韦, 武嘉文, 刘长发, 方蕾, 刘远, 苑静涵, 李晋. 翅碱蓬生态化学计量内稳性对模拟氮磷沉降的响应. 水生态学杂志, 2017, 38(4): 18-26. |
| [16] |
Yu Q, Elser J J, He N P, Wu H H, Chen Q S, Zhang G M, Han X G. Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland. Oecologia, 2011, 166(1): 1-10. DOI:10.1007/s00442-010-1902-z |
| [17] |
汪宗飞, 郑粉莉. 黄土高原子午岭地区人工油松林碳氮磷生态化学计量特征. 生态学报, 2018, 38(19): 6870-6880. |
| [18] |
刘国彬, 上官周平, 姚文艺, 杨勤科, 赵敏娟, 党小虎, 郭明航, 王国梁, 王兵. 黄土高原生态工程的生态成效. 中国科学院院刊, 2017, 32(1): 11-19. |
| [19] |
Deng J, Sun P S, Zhao F Z, Han X H, Yang G H, Feng Y Z, Ren G X. Soil C, N, P and its stratification ratio affected by artificial vegetation in subsoil, loess Plateau China. PLoS One, 2016, 11(3): e0151446. DOI:10.1371/journal.pone.0151446 |
| [20] |
葛晓改, 曾立雄, 肖文发, 黄志霖, 周本智. 模拟N沉降下不同林龄马尾松林凋落叶分解-土壤C、N化学计量特征. 生态学报, 2017, 37(4): 1147-1158. |
| [21] |
鲍士旦. 土壤农化分析 (第三版). 北京: 中国农业出版社, 2000.
|
| [22] |
Wu J, Joergensen R G, Pommerening B, Chaussod R, Brookes P C. Measurement of soil microbial biomass C by fumigation——extraction——an automated procedure. Soil Biology and Biochemistry, 1990, 22(8): 1167-1169. DOI:10.1016/0038-0717(90)90046-3 |
| [23] |
冉宜凡, 许明祥, 李彬彬, 马露洋, 张圣民. 黄土丘陵区不同土壤-微生物-植物系统生态化学计量特征对肥力梯度的响应. 西北农林科技大学学报:自然科学版, 2017, 45(10): 77-84, 93-93. |
| [24] |
孙岩, 何明珠, 王立. 降水控制对荒漠植物群落物种多样性和生物量的影响. 生态学报, 2018, 38(7): 2425-2433. |
| [25] |
贺纪正, 陆雅海, 傅伯杰. 土壤生物学前沿. 北京: 科学出版社, 2015.
|
| [26] |
刘玉林, 朱广宇, 邓蕾, 陈磊, 上官周平. 黄土高原植被自然恢复和人工造林对土壤碳氮储量的影响. 应用生态学报, 2018, 29(7): 2163-2172. |
| [27] |
邓强, 李婷, 袁志友, 焦峰. 黄土高原4种植被类型的细根生物量和年生产量. 应用生态学报, 2014, 25(11): 3091-3098. |
| [28] |
蒋跃利, 赵彤, 闫浩, 黄懿梅, 安韶山. 黄土丘陵区不同土地利用方式对土壤微生物量碳氮磷的影响. 水土保持通报, 2013, 33(6): 62-68. |
| [29] |
Elser J J, Sterner R W, Gorokhova E, Fagan W F, Markow T A, Cotner J B, Harrison J F, Hobbie S E, Odell G M, Weider L W. Biological stoichiometry from genes to ecosystems. Ecology Letters, 2000, 3(6): 540-550. DOI:10.1046/j.1461-0248.2000.00185.x |
| [30] |
马任甜, 安韶山, 黄懿梅. 黄土高原不同林龄刺槐林碳、氮、磷化学计量特征. 应用生态学报, 2017, 28(9): 2787-2793. |
| [31] |
Ågren G I. Stoichiometry and nutrition of plant growth in natural communities. Annual Review of Ecology, Evolution, and Systematics, 2008, 39: 153-170. DOI:10.1146/annurev.ecolsys.39.110707.173515 |
| [32] |
Han W X, Fang J Y, Guo D L, Zhang Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytologist, 2005, 168(2): 377-385. DOI:10.1111/j.1469-8137.2005.01530.x |
| [33] |
马露莎, 陈亚南, 张向茹, 杨佳佳, 安韶山. 黄土高原刺槐叶片生态化学计量学特征. 水土保持研究, 2014, 21(3): 57-61, 67-67. |
| [34] |
朱秋莲, 邢肖毅, 张宏, 安韶山. 黄土丘陵沟壑区不同植被区土壤生态化学计量特征. 生态学报, 2013, 33(15): 4674-4682. |
| [35] |
曾全超, 李鑫, 董扬红, 安韶山. 黄土高原延河流域不同植被类型下土壤生态化学计量学特征. 自然资源学报, 2016, 31(11): 1881-1891. DOI:10.11849/zrzyxb.20160038 |
| [36] |
胡培雷, 王克林, 曾昭霞, 张浩, 李莎莎, 宋希娟. 喀斯特石漠化地区不同退耕年限下桂牧1号杂交象草植物-土壤-微生物生态化学计量特征. 生态学报, 2017, 37(3): 896-905. |
| [37] |
Kou M, Garcia-Fayos P, Hu S, Jiao J Y. The effect of Robinia pseudoacacia afforestation on soil and vegetation properties in the Loess Plateau (China):a chronosequence approach. Forest Ecology and Management, 2016, 375: 146-158. DOI:10.1016/j.foreco.2016.05.025 |
| [38] |
周正虎, 王传宽. 微生物对分解底物碳氮磷化学计量的响应和调节机制. 植物生态学报, 2016, 40(6): 620-630. |
| [39] |
田中民. 根系分泌物在植物磷营养中的作用. 咸阳师范学院学报, 2001, 16(6): 60-63, 69-69. DOI:10.3969/j.issn.1672-2914.2001.06.020 |
| [40] |
Zhou Z H, Wang C K, Jiang L F, Luo Y Q. Trends in soil microbial communities during secondary succession. Soil Biology and Biochemistry, 2017, 115: 92-99. DOI:10.1016/j.soilbio.2017.08.014 |
| [41] |
Yu Q, Wilcox K, La Pierre K, Knapp A K, Han X G, Smith M D. Stoichiometric homeostasis predicts plant species dominance, temporal stability, and responses to global change. Ecology, 2015, 96(9): 2328-2335. DOI:10.1890/14-1897.1 |