 2019, Vol. 39
2019, Vol. 39文章信息
- 石丹, 倪九派, 倪呈圣, 刘锦春
- SHI Dan, NI Jiupai, NI Chengsheng, LIU Jinchun
- 巫山高山移民迁出区不同弃耕年限对植物物种多样性的影响
- The effects of different restoration years on plant diversity in Wushan's alpine emigrant region
- 生态学报. 2019, 39(15): 5584-5593
- Acta Ecologica Sinica. 2019, 39(15): 5584-5593
- http://dx.doi.org/10.5846/stxb201802260392
-
文章历史
- 收稿日期: 2018-02-26
- 网络出版日期: 2019-05-16





2. 西南大学生命科学学院, 重庆 400715
2. School of life sciences, Southwest University, Chongqing 400715, China
群落演替是植被生态学的重要研究内容, 对区域植物群落演替规律的认识, 是植物群落管理、利用改造和生态修复的基础依据。物种多样性变化更是群落演替进程中的一个重要指标, 它反应了群落组成和结构的变化规律[1-3]。国内外学者对其进行了大量的研究, 包括群落演替特征和驱动机制[4-8]、群落演替过程中物种多样性变化[9-14]、不同生境下的群落演替研究[15-25]、生境改变对植物多样性的影响[26-28]。渝东南、渝东北地区处于武陵山脉、秦巴山脉地带, 具有典型的喀斯特地貌, 土壤石漠化现象突出。随着高山生态移民搬迁进行, 迁出地人为干扰减少, 有效的维护了当地生态平衡和生物多样性, 推动了高山地区生态环境的恢复。移民后, 形成大量的弃耕地, 弃耕地的植物群落演替过程属于次生裸地上的进展研究。本研究以重庆市巫山县高山移民迁出区不同年限的弃耕地为对象, 对弃耕地植被自然恢复过程和规律进行了初步探索, 以期能对高山移民迁出区的植物管理、生态效益评估、撂荒地群落演替阶段研究提供参考, 加快修复脆弱的高山生态环境。
1 研究区域概况研究区位于重庆巫山五里坡自然保护区(31°21′—31°22′N, 110°5′—110°6′E)境内, 是典型的高山移民搬迁区迁出点, 属于中深切割中山地形, 平均海拔1415 m。年平均气温18.4℃, 平均降雨量1049.3 mm, 降水集中在7、8月份。土壤以山地黄壤、紫色土为主, 主要种植粮食作物和经济作物。自然植被有柏木(Cupressus funebris)、胡桃(Juglans regia)、油杉(Keteleeria fortunei)、臭椿(Ailanthus altissima), 梧桐(Firmiana platanifolia)、白花泡桐(Paulownia fortunei)、落叶松(Larix gmelinii)、瓜子黄杨(Buxus sinica)、红豆杉(Taxus chinensis)、崖栢(Thuja sutchuenensis)等。未搬迁之前人口为2580人。2009年对搬迁后剩余人口进行调查, 人口数为127人。
2 研究方法 2.1 样地设置与调查在2015年8—9月, 采用空间序列替代时间序列的方法进行采样。选取了土壤类型、地型相似但是弃耕年限不同的撂荒坡耕地作为采样地点, 撂荒地恢复的年限分别为1a、5a、8a。取当地居民耕地作为对照。不同类型样地分别设置3个标准样地。在每个标准样地范围内按照坡上、坡中、坡下的顺序取3个样方。草本群落样地取2 m×2 m的样方, 灌木以及乔木林地取5 m×5 m的样方。调查内容包括乔木种类以及其高度、冠幅、胸径, 灌木种类以及其高度、冠幅、胸径, 草本种类以及其数目、植株高、盖度。调查样地特征及基本情况如表 1。
| 指标 Index |
不同弃耕年限Different abandoned years | ||||
| A1 | A2 | A3 | A4 | R1 | |
| 坡向Slope aspect | N | N | N | N | N |
| 坡位Slope position | 坡中 | 坡中 | 坡中 | 坡中 | 坡中 |
| 坡度Slope degree /° | 32.5 | 32 | 30 | 31 | 32.5 |
| 海拔Altitude/m | 1133 | 1134 | 1137 | 1130 | 1135 |
| 群落类型Vegetational type | 农作物 | 草本 | 草本 | 草本、灌木、乔木 | 草本、灌木、乔木 |
| 草本盖度Herbage coverage/% | 50 | 90 | 75 | 20 | 30 |
| 灌木郁闭度Shrub canopy density/% | — | — | — | 23 | 30 |
| 乔木郁闭度Forest canopy density/% | — | — | — | 60 | 5 |
| N:北, North;A1:耕地, Cultivated land;A2:弃耕1a耕地, Cultivated land abandoned for 1 year;A3:弃耕5a耕地, Cultivated land abandoned for 5 years;A4:弃耕8a耕地, Cultivated land abandoned for 8 years;R1:弃耕5a生长着大量白花银背藤, Cultivated land abandoned 5 years grew Argyreia seguinii | |||||
物种多样性测度指标选取α多样性测度指标, 包括:Margalef丰富度指数(R)、Simpson优势度指数(D)、Shannon多样性指数(H′)、Pielou均匀度指数(Jw)。以及用相似性指数Jaccard(Cj)和Sorenson(Cs)和多样性阈值(Dv)。研究不同弃耕年限撂荒地、耕地、原始林地的植物多样性, 其计算公式如下:
(1) Margalef丰富度指数:
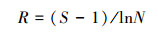
|
(1) |
式中, S为物种数目;N为所有物种个数之和。
(2) Simpson优势度指数:
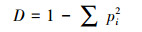
|
(2) |
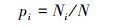
|
(3) |
式中, Ni为属于第i种物种的有Ni个;N为所有物种个数之和。
(3) Shannon多样性指数:
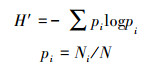
|
(4) |
式中, Ni为属于第i种物种的有Ni个;N为所有物种个数之和。
(4) Pielou均匀度指数:
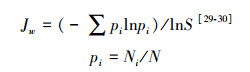
|
(5) |
式中, Ni为属于第i种物种的有Ni个;N为所有物种个数之和;S为物种数目。
(5) Jaccard相似性指数

|
(6) |
式中, j为2个群落或样地共有种数, a和b分别为样地A和样地B的物种数。
(6) Sorenson相似性指数

|
(7) |
式中, j为2个群落或样地共有种数, a和b分别为样地A和样地B的物种数。
(7) 多样性阈值
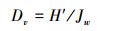
|
(8) |
对不同样地不同生长型的多样性指数进行加权, 权重的计算公式如下所示
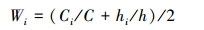
|
(9) |
式中, C为群落总盖度:C=∑Ci(i=1, 乔木层;i=2, 灌木层;i=3, 草本层);h为群落各生长型的平均高度总和:h=∑hi (i=1, 乔木层;i=2, 灌木层;i=3, 草本层);Wi为群落第i个生长型多样性指数加权参数。Ci为第i个生长型的盖度;hi为第i个生长型的平均高度。乔木层的林冠厚度按照乔木层平均高度的1/3计算, 灌木层按照灌木层平均高度的1/2计算, 草本层按照草本层平均高度的100%计算[31]。
群落总体多样性指数按照以下公式计算:

|
(10) |
式中, W1、W2、W3分别为乔木层、灌木层、草本层的加权参数;D1、D2、D3分别为乔木层、灌木层、草本层的多样性指数。

|
(11) |
以上公式中, 根据实际调查内容中, 对草本层使用相对高度代替相对密度, 乔木层的相对密度是单位面积的乔木株数。乔木层中的相对盖度主要是相对基盖度[32]。
数据统计与分析使用SPSS18.0软件进行, 不同数据组间的差异水平的比较采用的是单因素方差分析法(LSD)。
根据植物物种多样性阈值分级评价标准对不同弃耕年限样地植被群落多样性进行评定, 植物物种多样性阈值分级评价标准见表 2。
调查可知, 在不同群落中, 总共拥有的植物种类有28科40属46种。其中蕨类植物有3科3属3种。藤本植物有1科1属1种, 乔木3科3属3种, 灌木6科6属7种。种类数目较多的科分别为:菊科(Compositae)7种, 蔷薇科(Rosaceae)5种, 禾本科(Gramineae)3种, 伞形科(Umbelliferae)3种。
群落中存在大量偶见种, 在A2样地中, 个体数只有一株的有:小蓬草(Conyza canadensis)、长柄山蚂蝗(Podocarpium podocarpum)、蛇莓(Duchesnea indica)。
在A3样地中, 个体数只有一株的植物有:牛膝(Achyranthes bidentata)、马唐(Digitaria sanguinalis)。
在A4样地中, 个体数只有一株的有:油桐(Vernicia fordii)、长叶女贞(Ligustrum compactum)、樱桃(Cerasus pseudocerasus)、乌蕨(Stenoloma chusanum)、蕨(Pteridium aquilinum)、马唐(Digitaria sanguinalis)、灯笼草(Clinopodium polycephalum)。
在R1样地中, 个体数只有一株的有:小蓬草(Conyza Canadensis)、磨芋(Amorphophallus rivieri)、光滑高粱泡(Rubus lambertianus)、山莓(Rubus corchorifolius)。
由表 3可知A2、A3、A4都由10科以上的植物组成, 其中种类最多的是A3, 由17科植物组成。种类最少的是A1, 只有1科植物组成。A1植被类型为农作物, A2、A3、R1植被类型为草本+灌木, A4植被类型为草本+灌木+乔木。
| 样地类型 Types |
科 Family |
属 Genus |
种 Species |
物种数Numbers of species | |||
| 乔木Tree | 灌木Shrub | 草本Herb | 藤本Vine | ||||
| A1 | 1 | 1 | 1 | 0 | 0 | 1 | 0 |
| A2 | 12 | 19 | 19 | 0 | 2 | 17 | 1 |
| A3 | 17 | 23 | 24 | 0 | 1 | 23 | 0 |
| A4 | 13 | 16 | 17 | 3 | 4 | 10 | 0 |
| R1 | 6 | 8 | 9 | 0 | 1 | 8 | 0 |
不同群落物种组成和结构特征如下表 3所示。
重要值在一定程度上评价了物种在群落结构中的优势地位, 是一种植物相当于另外一种植物在资源占据以及利用方面的优势度。
由表 4可知, A2草本层前三的优势物种依次为艾(Artemisia argyi)、磨芋(Amorphophallus rivieri)、野胡萝卜(Daucus carota), 其重要值分别为0.185、0.096、0.083。A2灌木层有两种植物:寒莓(Rubus buergeri)和猫儿刺(Ilex pernyi), 其重要值为0.590和0.410。A3草本层前三的优势物种依次为小蓬草(Conyza canadensis)、一年蓬(Erigeron annuus)、大麻(Cannabis sativa), 其重要值分别为0.197、0.092、0.074。A3灌木层只有一种植物密蒙花(Buddleja officinalis)。A4草本层前三的优势物种依次为顶芽狗脊(Woodwardia unigemmata)、白茅(Imperata cylindrica)、千里光(Senecio scandens), 其重要值分别为0.229、0.181、0.119。A4灌木层前三的优势物种依次为山莓(Rubus corchorifolius)、胡枝子(Lespedeza bicolor)、长叶女贞(Ligustrum compactum), 其重要值分别为0.325、0.192、0.164。A4乔木层有三种植物, 依次为胡桃(Juglans regia)、油桐(Vernicia fordii)、樱桃(Cerasus pseudocerasus), 其重要值分别为0.581、0.215、0.205。R1草本层前三的优势物种依次为白花银背藤(Argyreia seguinii)、菟丝子(Cuscuta chinensis)、艾(Artemisia argyi), 其重要值分别为0.277、0.170、0.138。R1灌木层只有一种植物:山莓(Rubus corchorifolius)。
| 样地类型 Types |
层次 Layers |
种名 Species |
重要值 Important value |
| A1 | 农作物 | 白菜Brassica pekinensis | 1.000 |
| A2 | 草本 | 艾Artemisia argyi | 0.185 |
| 磨芋Amorphophallus rivieri | 0.096 | ||
| 野胡萝卜Daucus carota | 0.083 | ||
| 灌木 | 寒莓Rubus buergeri | 0.590 | |
| 猫儿刺Ilex pernyi | 0.410 | ||
| A3 | 草本 | 小蓬草Conyza canadensis | 0.197 |
| 一年蓬Erigeron annuus | 0.092 | ||
| 大麻Cannabis sativa | 0.074 | ||
| 灌木 | 密蒙花Buddleja officinalis | 1.000 | |
| 菟丝子Cuscuta chinensis | 0.170 | ||
| 艾Artemisia argyi | 0.138 | ||
| 灌木 | 山莓Rubus corchorifolius | 1.000 | |
| A4 | 草本 | 顶芽狗脊Woodwardia unigemmata | 0.229 |
| 白茅Imperata cylindrica | 0.181 | ||
| 千里光Senecio scandens | 0.119 | ||
| 灌木 | 山莓Rubus corchorifolius | 0.325 | |
| 胡枝子Lespedeza bicolor | 0.192 | ||
| 长叶女贞Ligustrum compactum | 0.164 | ||
| 乔木 | 胡桃Juglans regia | 0.581 | |
| 油桐Vernicia fordii | 0.215 | ||
| 樱桃Cerasus pseudocerasus | 0.205 | ||
| R1 | 草本 | 白花银背藤Argyreia seguinii | 0.277 |
| 菟丝子Cuscuta chinensis | 0.170 | ||
| 艾Artemisia argyi | 0.138 | ||
| 灌木 | 山莓Rubus corchorifolius | 1.000 |
弃耕5a, 群落有两种发展趋势, 一种是小蓬草(Conyza canadensis)为优势植物, 其重要值为0.197, 另一种是以白花银背藤(Argyreia seguinii)为优势作物, 其重要值为0.277。
3.2 不同群落植物物种多样性比较物种多样性是用来判断群落以及生态系统的稳定性指标, 包括物种的丰富度以及均匀度。物种的多样性越大, 表示群落的稳定性越强, 则此生境越有利于植被生存[34]。
由表 5可知, A1、A2、A3、A4、R1草本层、灌木层植物丰富度指数(R)、均匀度指数(Jw)、优势度指(D)数、多样性指数(H′)差异显著情况。
| 层次 Layer |
样地 Plots |
Margalef丰富度指数(R) Margelef richness index |
Shannon多样性指数(H′) Shannon diversity index |
Pielou均匀度指数(Jw) Pielou evenness index |
Simpson优势度指数(D) Simpson dominance index |
| 草本层Herb layer | A1 | 0.000e | 0.000d | 0.000d | 0.000d |
| A2 | 3.006b | 0.938a | 0.762b | 0.817a | |
| A3 | 3.831a | 0.956a | 0.702b | 0.800b | |
| A4 | 2.427c | 0.826b | 0.866a | 0.818a | |
| R1 | 1.710d | 0.516c | 0.571c | 0.529c | |
| 灌木层Shrub layer | A1 | — | — | — | — |
| A2 | 0.621b | 0.292b | 0.954b | 0.480b | |
| A3 | 0.000c | 0.000c | 0.000c | 0.000c | |
| A4 | 1.559a | 0.642a | 0.919a | 0.746a | |
| R1 | 0.000c | 0.000c | 0.000c | 0.000c | |
| 乔木层Tree layer | A1 | — | — | — | — |
| A2 | — | — | — | — | |
| A3 | — | — | — | — | |
| A4 | 1.443 | 0.452 | 0.946 | 0.625 | |
| R1 | — | — | — | — | |
| 总体多样性指数 | A1 | 0.000e | 0.000d | 0.000e | 0.000e |
| Total diversity index | A2 | 2.609b | 0.830a | 0.794b | 0.761a |
| A3 | 2.735a | 0.683b | 0.501c | 0.571c | |
| A4 | 1.837c | 0.638b | 0.910a | 0.726b | |
| R1 | 1.392d | 0.420c | 0.465d | 0.430d | |
| 同列不同小写字母表示不同类型样地间差异显著(P < 0.05) | |||||
A3和R1灌木层只有一种灌木, 因此灌木层各多样性指数为0。只有A4存在草灌乔群落。不同群落灌木层多样性指数变化情况如下, 丰富度指数R、多样性指数H′、优势度指数D数值均为:A4>A2。均匀度指数Jw数值为:A2>A4。R1各个层次的多样性指数以及总体多样性指数均小于除了对照A1的其他群落。
由图 1可知, 随弃耕年限的增长, 不同群落草本层多样性指数变化情况如下:丰富度指数R的大小依次为:A3>A2>A4>A1, 多样性指数H′数值的大小依次为:A3>A2>A4>A1, 均匀度指数Jw数值大小依次为:A4>A2>A3>A1, 优势度指数D数值大小依次为:A4>A2>A3>A1。随弃耕年限的增长, 丰富度指数R与多样性指数H′的变化趋势一致, 均先增加后减少, 在A3达到最高。均匀度指数Jw和优势度指数D的变化趋势一致, 均先减小后增加, 在A4达最高。

|
| 图 1 不同群落草本层植物物种多样性指数 Fig. 1 The herbaceous diversity index in different communities A1:耕地, Cultivated land;A2:弃耕1a耕地, Cultivated land abandoned for 1 year;A3:弃耕5a耕地, Cultivated land abandoned for 5 years;A4:弃耕8a耕地, Cultivated land abandoned for 8 years;R:Margalef丰富度指数, Margalef richness index;H′:Shannon多样性指数, Shannon diversity index;Jw:Pielou均匀度指数, Pielou evenness index;D:Simpson优势度指数, Simpson dominance index |
由图 2可知, 随着弃耕年限的增长, 不同群落总体多样性指数变化情况如下:总体丰富度指数Rt的大小依次为:A3>A2>A4>A1, 总体多样性指数Ht′数值的大小依次为:A2>A3>A4>A1, 总体均匀度指数Jtw数值大小依次为:A4>A2>A3>A1, 总体优势度指数Dt数值大小依次为:A2>A4>A3>A1。随弃耕年限的增长, 总体丰富度指数Rt先增加后减少, 在A3处达到最大, 总体多样性指数Ht′一直减小, 在A2处最大, 总体均匀度指数Jtw先减小后增加, 在A4处达到最大, 总体优势度指数Dt先增加后减少, 在A4达最高。

|
| 图 2 不同群落植物物种总体多样性指数 Fig. 2 The total plant diversity index in different communities Rt:总体丰富度指数, Total richness index;Ht′:总体多样性指数, Total diversity index;Jtw:总体均匀度指数, Total evenness index;Dt:总体优势度指数, Total dominance index |
A1为农耕地, 植被为单一农作物, 因此不将A1与其他不同弃耕年限样地进行相似性指数计算与比较。
从图 3可以看出, A2和A3的相似性指数最高, 其Jaccard相似性指数值为:0.344, Sorenson相似性指数值为:0.512。然后是A2和R1, Jaccard相似性指数值为:0.217, Sorenson相似性指数值为:0.357;再是A2和A4, Jaccard相似性指数值为:0.200, Sorenson相似性指数值为:0.333。再是A3和R1, 其Jaccard相似性指数值为:0.179, Sorenson相似性指数值为:0.303。再是A4和R1, 其Jaccard相似性指数值为:0.083, Sorenson相似性指数值为:0.154。相似性指数最小的是A3和A4, 其Jaccard相似性指数值为:0.051, Sorenson相似性指数值为:0.098。弃耕初期A2和A3、R1的相似性指数都很高, 可见弃耕地与相邻年限的样地之间相似性最高, 而A3与A4之间相似性指数低的原因是, A3是草本层为主, 灌木层只有一种植被, 而A4具有草本、灌木、乔木三种类型, 它们之间群落结构差异巨大。

|
| 图 3 不同群落组成间的相似性 Fig. 3 Similarity of species composition between different communities Cj:Jaccard相似性指数, Jaccard similarity index;Cs:Sorenson相似性指数, Sorenson similarity index |
不同群落的植物物种多样性阈值见表 6。
| 模式Mode | 层次Layer | A1 | A2 | A3 | A4 | A5 |
| 物种多样性阈值(Dv) | 草本 | 0.000 | 1.230 | 1.362 | 0.954 | 0.903 |
| Threshold of plant | 灌木 | — | 0.306 | 0.000 | 0.699 | 0.000 |
| species diversity | 乔木 | — | — | — | 0.477 | — |
由表 6可知, 不同弃耕年限样地草本层的群落多样性阈值最大的是A3, 多样性阈值为1.362, 其次是A2、A4和R1, 多样性阈值分别为1.230、0.954、0.903, A1的多样性阈值最小为0。由表 2植物物种多样性阈值的分级评价标准可知, A2、A3、A4、R1属于Ⅳ级, 说明耕地在弃耕1a、5a、8a后, 其草本植物物种多样性一般。Y1属于Ⅴ级, 说明农耕地的草本植物物种多样性差。不同弃耕年限样地灌木层的群落多样性阈值最大的是A4, 多样性阈值为0.699, 其次是A2, 多样性阈值为0.306, 最后是A3和R1, 多样性阈值为0。根据植物物种多样性阈值的分级评价标准可知, A4属于Ⅳ级, 说明其耕地在弃耕8a后, 其灌木植物物种多样性一般。A2、A3、R1属于Ⅴ级, 说明弃耕地在弃耕1a、5a后, 其灌木植物物种多样性差。本研究只有弃耕8a后的耕地出现了乔木层, 其多样性阈值为0.477, 按照植物物种多样性阈值的分级标准可知, 其耕地弃耕8a后, 其乔木层植物物种多样性差。
4 讨论 4.1 弃耕年限对群落物种组成及结构的影响弃耕地植物群落恢复演替属于次生演替, 包括植物繁殖体的传播、定居、群聚、竞争、反应、稳定等阶段[35]。随着演替时间的推移, 弃耕地植物的群落结构由简单到复杂, 物种组成也发生变化[36]。在自然演替的情况下, 弃耕地植被演替阶段分为:草本植物群落阶段→灌丛植物群落阶段→乔木群落阶段, 到达乔木群落阶段, 群落相当稳定, 总历时为40—50a。在陕西省丹汉江流域弃耕区形成灌草群落需7—10a, 形成草灌乔群落则需要30a以上[37]。本研究中, 随着弃耕年限的增加, 群落演替阶段为:灌丛植物群落阶段→乔木群落阶段, 历时为8a。这是因为研究区丰富的水热条件, 以及弃耕地中原本施有丰富的氮、磷、钾等营养元素, 加快植被恢复进程。退耕还草演替初期, 一年生植物较多, 它们生活周期短、种子数量多、传播能力强, 在短时间内成为优势种, 一年生植物的生长改善了土壤环境, 植物残体加快了土壤有养分累积。在演替中期, 出现多年生植物, 它们根系强大、竞争扩张能力强、保水力强, 进入群落并逐渐成为优势种。在群落的后期, 一般以耐阴植物为主[38]。这与本研究的结果一致。本研究中, 弃耕1a, 群落的优势物种是艾(Artemisia argyi), 弃耕5a, 群落的优势物种是小蓬草(Conyza canadensis), 弃耕8a, 群落的优势作物是顶芽狗脊(Woodwardia unigemmata)。艾(Artemisia argyi)是喜阳喜湿润的多年生草本, 小蓬草(Conyza canadensis)是喜阳喜湿润的一年生草本, 顶芽狗脊(Woodwardia unigemmata)和白茅(Imperata cylindrica)是耐阴喜湿的多年生草本, 因此可以看出, 随着弃耕年限的增长, 撂荒地优势物种由喜阳的一年生或多年生草本向着耐阴的多年生草本转变。由于灌木和乔木的生长, 林隙下光线减少, 草本盖度种类减小, 耐阴的多年生草本开始入侵成为优势种。在亚热带山地常绿阔叶林中阴性植物的个体数占总地表植物的大部分[39]。植物群落的分布、数量、个体大小除了受相互间的影响外, 还受到生境、干扰程度、以及遗传特性的影响, 是环境中各种因素综合作用的结果[40-41]。弃耕8a, 灌层优势作物为山莓(Rubus corchorifolius), 山莓(Rubus corchorifolius)是荒地植被演替中的先锋物种。
4.2 弃耕年限对群落植物物种多样性的影响本研究中, 弃耕5a, 群落出现了两种发展趋势, 一种是以白花银背藤(Argyreia seguinii)为优势物种的群落, 一种是以小蓬草(Conyza canadensis)为优势物种的群落。弃耕5a生长着大量白花银背藤(Argyreia seguinii)的群落的各层次的多样性指数以及总体多样性指数均小于除了对照的其他群落, 白花银背藤(Argyreia seguinii)的大量生长阻碍了群落演替进程。白花银背藤(Argyreia seguinii)曾被划分为湖北省中度危害的林业有害植物[42-43], 它繁殖生长能力强, 生长在植被表面, 与其他植物竞争光能和养分, 从而限制其他作物生长。在群落恢复过程中提高对白花银背藤(Argyreia seguinii)生长状态的监控, 消除对植物物种多样性的影响。
随着弃耕年限的增长, 群落草本层多样性指数H′和丰富度指数R先增加后减少, 这与郝红敏在典型草原撂荒地的研究结果一致。原因可能是演替初期, 一年生草本植物较多, 导致多样性和丰富度指数较高, 随着演替的进行, 多年生草本出现, 成为优势种, 一年生草本植物在竞争中被淘汰, 从而多样性和丰富度指数降低, 演替后期, 多年生草本个体变大, 下方空间被耐阴的植株占据, 因此多样性和丰富度指数再次升高[38]。有研究表明, 植物物种多样性随演替时间的延长而增长, 在演替中后期达到最高值, 随后又下降。多样性指数H′最高为0.956, 没有超过4.5[44], 弃耕多年草本层植物物种多样性指数不高, 表明群落植被尚处于恢复期。
4.3 群落植物的多样性评估按照植物物种多样性阈值的分级标准可知, 群落草本层、灌木层植物多样性最高达到Ⅳ级, 表示其植物物种多样性“一般”。群落乔木层植物多样性最高到达Ⅴ级。表示其植物物种多样性“差”。经过8a的恢复, 研究区植物物种多样性未达到“较好”状态, 需要长期的恢复才能达到植被丰富的状态。
庙堂村处于重庆巫山五里坡自然保护区, 实施移民搬迁对于该自然保护区的植被生态恢复非常有利。实施高山移民搬迁以来, 耕地在自然恢复过程中, 逐渐形成稳定的植物群落, 有助于当地生态建设和保护。在恢复过程中, 可以利用撂荒地植被演替的规律, 引入相应演替阶段的植物, 加速撂荒地植被的恢复过程, 适当引进本土植被, 比如小蓬草(Conyza canadensis)、巫山杜鹃(Rhododendron roxieoides)、野胡萝卜(Daucus carota)等, 提高群落丰富度。同时加大对白花银背藤(Argyreia seguinii)的监测和管理, 防止其大量生长破坏生态结构。
5 结论随着弃耕年限的增长, 植被群落结构发生变化:草+灌→草+灌+乔。群落草本层优势物种发生改变, 由一年生喜阳草本或者田间杂草变为多年生耐阴草本植物。草本层和灌木层植物物种多样性随着演替年限的延长而增长, 在演替的中后期达到最大值。演替的过程中出现白花银背藤(Argyreia seguinii), 影响了其他草本的生长, 阻碍演替进程, 需对其加强监督与治理。对不同弃耕年限样地植被群落多样性进行评定, 都未达到Ⅲ级别, 说明目前研究年限植物群落的恢复状况未达到较好状态, 需要长期的恢复才能达到植被丰富的状态。
| [1] |
Park H C, Lee G G, Lee J H. Analysis of vegetation recovery trend in abandoned paddy wetland-Focused on flora and vegetation changes of clear zone surrounding Pyeongtaek K-55 Air Force Base-. KSCE Journal of Civil Engineering, 2015, 19(4): 864-872. DOI:10.1007/s12205-014-1634-4 |
| [2] |
Kämpf I, Mathar W, Kuzmin I, Hölzel N, Kiehl K. Post-Soviet recovery of grassland vegetation on abandoned fields in the forest steppe zone of Western Siberia. Biodiversity and Conservation, 2016, 25(12): 2563-2580. DOI:10.1007/s10531-016-1078-x |
| [3] |
马长明, 袁玉欣. 国内外退耕地植被恢复研究现状. 世界林业研究, 2004, 17(4): 24-27. DOI:10.3969/j.issn.1001-4241.2004.04.005 |
| [4] |
Clements F E. Nature and structure of the climax. Journal of Ecology, 1936, 24(1): 252-284. DOI:10.2307/2256278 |
| [5] |
Willis K J, Whittaker R J. Ecology——Species diversity——scale matters. Science, 2002, 295(5558): 1245-1248. DOI:10.1126/science.1067335 |
| [6] |
Gleason H A. The structure and development of the plant association. Bulletin of the Torrey Botanical Club, 1917, 44(10): 463-481. DOI:10.2307/2479596 |
| [7] |
范玮熠, 王孝安, 郭华. 黄土高原子午岭植物群落演替系列分析. 生态学报, 2006, 26(3): 706-714. DOI:10.3321/j.issn:1000-0933.2006.03.010 |
| [8] |
文丽, 宋同清, 杜虎, 王克林, 彭晚霞, 曾馥平, 曾昭霞, 何铁光. 中国西南喀斯特植物群落演替特征及驱动机制. 生态学报, 2015, 35(17): 5822-5833. |
| [9] |
刘硕, 贺康宁, 王晓江. 鄂尔多斯沙地不同退牧年限植物群落多样性及变异性研究. 西北植物学报, 2010, 30(3): 586-592. |
| [10] |
李昌龙, 徐先英, 金红喜, 王多泽, 李菁菁. 玛曲高寒草甸沙化过程中群落结构与植物多样性. 生态学报, 2014, 34(14): 3953-3961. |
| [11] |
王世雄, 王孝安, 李国庆, 郭华, 朱志红. 陕西子午岭植物群落演替过程中物种多样性变化与环境解释. 生态学报, 2010, 30(6): 1638-1647. |
| [12] |
刘振国, 李镇清. 退化草原冷蒿群落13年不同放牧强度后的植物多样性. 生态学报, 2006, 26(2): 475-482. DOI:10.3321/j.issn:1000-0933.2006.02.021 |
| [13] |
高凯, 朱铁霞, 韩国栋. 围封年限对内蒙古羊草-针茅典型草原植物功能群及其多样性的影响. 草业学报, 2013, 22(6): 39-45. |
| [14] |
李裕元, 邵明安. 子午岭植被自然恢复过程中植物多样性的变化. 生态学报, 2004, 24(2): 252-260. |
| [15] |
肖德荣, 田昆, 张利权. 滇西北高原纳帕海湿地植物多样性与土壤肥力的关系. 生态学报, 2008, 28(7): 3116-3124. |
| [16] |
罗宁, 刘尊驰, 于航, 刘彤. 古尔班通古特沙漠南部植物多样性的区域差异. 生态学报, 2016, 36(12): 3572-3581. |
| [17] |
盛茂银, 熊康宁, 崔高仰, 刘洋. 贵州喀斯特石漠化地区植物多样性与土壤理化性质. 生态学报, 2015, 35(2): 434-448. |
| [18] |
张雪妮, 吕光辉, 王庭权, 马玉, 阿布里孜·阿不都热合曼, 赵晓英, 郭振洁, 朱修逸. 荒漠区垂直河岸带植物多样性格局及其成因. 生态学报, 2015, 35(18): 5966-5974. |
| [19] |
卢训令, 汤茜, 梁国付, 丁圣彦. 黄河下游平原不同非农生境中植物多样性. 生态学报, 2015, 35(5): 1527-1536. |
| [20] |
张喜, 王莉莉, 刘延惠, 文弢, 崔迎春, 姜霞, 张佐玉, 霍达, 李丹. 喀斯特天然林植物多样性指数和土壤理化指标的相关性. 生态学报, 2016, 36(12): 3609-3620. |
| [21] |
贺金生, 陈伟烈. 陆地植物群落物种多样性的梯度变化特征. 生态学报, 1997, 17(1): 91-99. DOI:10.3321/j.issn:1000-0933.1997.01.014 |
| [22] |
卢群, 曾小康, 石俊慧, 陈里娥, 周凯, 雷安平, 昝启杰. 深圳湾福田红树林群落演替. 生态学报, 2014, 34(16): 4662-4671. |
| [23] |
Abella S R, Crouse J E, Covington W W, Springer J D. Diverse responses across soil parent materials during ecological restoration. Restoration Ecology, 2015, 23(2): 113-121. DOI:10.1111/rec.12160 |
| [24] |
Emery S M, Gross K L. Dominant species identity, not community evenness, regulates invasion in experimental grassland plant communities. Ecology, 2007, 88(4): 954-964. DOI:10.1890/06-0568 |
| [25] |
谢晋阳, 陈灵芝. 暖温带落叶阔叶林的物种多样性特征. 生态学报, 1994, 14(4): 337-344. DOI:10.3321/j.issn:1000-0933.1994.04.012 |
| [26] |
李文娇, 刘红梅, 赵建宁, 修伟明, 张贵龙, 皇甫超河, 杨殿林. 氮素和水分添加对贝加尔针茅草原植物多样性及生物量的影响. 生态学报, 2015, 35(19): 6460-6469. |
| [27] |
刘文亭, 卫智军, 吕世杰, 王天乐, 张爽. 放牧对短花针茅荒漠草原植物多样性的影响. 生态学报, 2017, 37(10): 3394-3402. |
| [28] |
温璐, 董世魁, 朱磊, 施建军, 刘德梅, 王彦龙, 马玉寿. 环境因子和干扰强度对高寒草甸植物多样性空间分异的影响. 生态学报, 2011, 31(7): 1844-1854. |
| [29] |
马克平. 生物群落多样性的测度方法Ⅰα多样性的测度方法(上). 生物多样性, 1994, 2(3): 162-168. DOI:10.3321/j.issn:1005-0094.1994.03.007 |
| [30] |
马克平, 刘玉明. 生物群落多样性的测度方法Ⅰα多样性的测度方法(下). 生物多样性, 1994, 2(4): 231-239. DOI:10.3321/j.issn:1005-0094.1994.04.009 |
| [31] |
徐远杰, 陈亚宁, 李卫红, 付爱红, 马晓东, 桂东伟, 陈亚鹏. 伊犁河谷山地植物群落物种多样性分布格局及环境解释. 植物生态学报, 2010, 34(10): 1142-1154. DOI:10.3773/j.issn.1005-264x.2010.10.003 |
| [32] |
刘锦春, 何丙辉, 徐小军, 胡恒, 熊明彪, 罗亚. 汶川草坡乡地震次生灾害迹地植物群落的恢复研究. 西南大学学报:自然科学版, 2013, 35(4): 51-56. |
| [33] |
蔡燕红.杭州湾浮游植物生物多样性研究[D].青岛: 中国海洋大学, 2006: 61-61. http://cdmd.cnki.com.cn/Article/CDMD-10423-2006181965.htm
|
| [34] |
陈志辉, 王克林, 陈洪松, 何寻阳. 喀斯特环境移民迁出区植物多样性研究. 中国生态农业学报, 2008, 16(3): 723-727. |
| [35] |
张海峰, 彭鸿, 陈存根, 张小林, 廖纯艳. 南水北调中线水源区弃耕地草本植被演替初步研究. 西北植物学报, 2005, 25(5): 973-978. DOI:10.3321/j.issn:1000-4025.2005.05.023 |
| [36] |
万雪琴.坡耕地退耕还林后的植被恢复及其改土效应[D].雅安: 四川农业大学, 2003: 59-59. http://cdmd.cnki.com.cn/Article/CDMD-10626-2003094355.htm
|
| [37] |
王星, 李占斌, 李鹏. 陕西省丹汉江流域退耕地恢复过程中的植被演替. 应用生态学报, 2012, 23(2): 347-356. |
| [38] |
郝红敏, 刘玉, 王冬, 杨政, 武高林. 典型草原开垦弃耕后不同年限群落植物多样性和空间结构特征. 草地学报, 2016, 24(4): 754-759. |
| [39] |
区余端, 苏志尧, 李镇魁, 林义辉. 地形因子对粤北山地森林不同生长型地表植物分布格局的影响. 应用生态学报, 2011, 22(5): 1107-1113. |
| [40] |
黄德青, 于兰, 张耀生, 赵新全. 祁连山北坡天然草地地下生物量及其与环境因子的关系. 草业学报, 2011, 20(5): 1-10. |
| [41] |
Mitchell J E, West N E, Miller R W. Soil physical properties in relation to plant community patterns in the shadscale zone of northwestern Utah. Ecology, 1966, 47(4): 627-630. DOI:10.2307/1933941 |
| [42] |
苏长江. 国家级自然保护区内有害生物葛藤的防治对策. 南方农业, 2016, 10(9): 126-126, 128-128. DOI:10.3969/j.issn.1673-890X.2016.09.072 |
| [43] |
戴丽, 陶惠萍, 曾祥福, 雷永松. 林业有害植物葛藤风险分析. 中国森林病虫, 2011, 30(1): 29-32. DOI:10.3969/j.issn.1671-0886.2011.01.009 |
| [44] |
刘琪璟, 胡理乐, 李轩然. 小流域治理20年后的千烟洲植物多样性. 植物生态学报, 2005, 29(5): 766-774. DOI:10.3321/j.issn:1005-264X.2005.05.009 |