 2019, Vol. 39
2019, Vol. 39文章信息
- 刘爽, 王雅, 刘兵兵, 刘海龙, 刘勇
- LIU Shuang, WANG Ya, LIU Bingbing, LIU Hailong, LIU Yong
- 晋西北不同土地管理方式对土壤碳氮、酶活性及微生物的影响
- Effects of different land management practices on soil carbon and nitrogen, enzyme activities, and microbial diversities northwest of Shanxi
- 生态学报. 2019, 39(12): 4376-4389
- Acta Ecologica Sinica. 2019, 39(12): 4376-4389
- http://dx.doi.org/10.5846/stxb201804030748
-
文章历史
- 收稿日期: 2018-04-03
- 修订日期: 2018-12-29





2. 山西省农业科学院农业环境与资源研究所, 山西省土壤环境与养分资源重点实验室, 太原 030031;
3. 中国农业科学院农业信息研究所/农业部农业信息服务技术重点实验室, 北京 100081
2. Institute of Agricultural Environment and Resources, Shanxi Academy of Agricultural Sciences, Key Laboratory of Soil Environment and Nutrient Resources of Shanxi Province, Taiyuan 030031;
3. Agricultural Information Institute, Chinese Academy of Agricultural Sciences, Key Laboratory of Agri-Information Service Technology, Ministry of Agriculture, Beijing 100081
土地利用和管理是人类利用土地各种活动的综合反映, 是影响土壤肥力变化的主要因素[1]。不同的土地利用和管理方式, 因植被覆被(草地、荒地、农地)、人为活动(耕作和免耕)等不同, 对土壤理化性状产生影响, 进而影响土壤酶活性在土壤垂直方向上的分布[2], 并对土壤微生物群落结构和多样性产生影响[3-4]。土壤酶在土壤生态系统的物质循环和能量流动方面扮演着重要角色, 它参与土壤的发生发育以及土壤肥力形成和演化的全过程, 具有高度催化作用[5]。土壤酶活性可反映土壤中生物化学过程的强度和方向, 可作为评价土壤肥力状况的指标, 也可作指示土壤生态系统的健康和可持续性以及土地管理引起为土壤质量变化的生物活性指标[6-8]。有研究表明, 肥料、作物残体和耕作管理方式可影响土壤酶的分布和活性[9-10]
土壤微生物群落是土壤重要的活体成分, 作为土壤养分转化和循环的催化剂, 是土壤碳氮元素矿化的来源, 其数量和活性是评价土壤质量的重要指标[11]。土壤微生物对环境变化具有敏感性, 不同的管理方式、土壤、植被条件下, 微生物各群落对环境因素的响应不同, 对其数量的消长和种群结构可产生较大的影响[12]。土壤微生物与土壤酶紧密联系, 在土壤生态系统服务传递过程中扮演重要的角色[13], 两者参与有机物质分解和合成, 以及无机物的氧化与还原的过程, 因此是土壤生态系统代谢的重要动力[14]。
为了更好地了解晋西北地区不同土地管理方式对土壤质量的影响, 本试验在山西省北部忻州市五寨县设置四种土地管理方式, 苜蓿样地、免耕样地、翻耕样地和荒地(对照), 研究不同管理方式对土壤肥力、土壤酶(蔗糖酶、磷酸酶、过氧化氢酶、脲酶)活性的影响, 分析不同管理方式土壤微生物群落结构和多样性, 以及微生物与土壤环境因子的关系, 评估不同土地管理方式对土壤质量的影响, 以期为晋西北地区土地管理和生态建设提供参考。
1 材料与方法 1.1 区域概况研究区位于山西省忻州市五寨县胡会乡石咀头村, 具体地理位置为111°28′—113°E和38°44′—39°17′N, 涵盖华北黄土高原的半干旱区和沙区。该区属于温带大陆性气候, 冬季受蒙古西伯利亚高压控制, 长而严寒, 且雨雪偏少;春季干旱多风, 气候干燥;夏季气温适中且雨量高度集中;该地区昼夜温差大, 年平均气温4.9℃左右, 1月最冷(-13.3℃), 7月最热(20.1℃)。无霜期120 d左右, 有效积温2452℃, 该区年平均降雨量在450—500 mm之间, 降水多集中于7月和8月, 约占年降水量的44%。根据中国土壤分类系统, 研究区土壤为沙黄土, 与土壤分类学中认为典型旱成土相似, 土壤质地松散, 孔隙度高, 渗透性好, 通气性强, 肥力低, 土壤有机质含量低[15]。
1.2 研究方法 1.2.1 样地选择及设置2015年8月下旬于山西省忻州市五寨县石咀头村, 选择四种在当地具有代表性的土地利用和管理方式进行试验研究:研究中选取的100 m(长)× 80 m(宽)的地块, 在实验当地进行调查, 该地块自1965年开始未受到人为干扰。该地块在实验前, 为撂荒地, 自2008年开始, 对该地块进行开垦。当地主要以传统翻耕种植玉米为主, 且多为多年连作的方式。本研究中的4块实验样地, 每一块实验样地为100 m(长)× 20 m(宽), 4块实验样地具体设置如下:一块样地种植紫花苜蓿, 从2008—2015年未进行耕作(免耕苜蓿样地MX), 苜蓿自2008年种植后, 每年秋季不进行收割, 不进行任何耕作措施;另一个样地于2008年春季, 进行常规的翻耕起垄, 于常规耕作下2008—2015年种植玉米(Zea mays L.)(翻耕样地FG), 于每年春季播种前翻耕一次, 翻耕深度约为50 cm, 秋季籽实收获后, 地上部分秸秆于第二年春季移除地块;第三块样地于2008年春季, 采用免耕播种机进行播种, 于2008—2015年免耕种植玉米(免耕样地MG), 秋季籽实收获后, 秸秆全部粉碎覆盖于地表;最后一个地块不进行任何耕种措施(荒地HD), 地表主要以草本为主, 且植被覆盖较少。
1.2.2 样品采集及预处理本实验在每个样地各设置10 m×10 m样方3个, 在每个样方内设置5个取样点, 采用5点混合取样法采集土壤样品, 即利用对角线法选取5个点, 首先去除地表植被和覆盖物, 再用土钻钻取0—50 cm深的土壤样品, 从上至下10 cm等间距取样, 每一个样方内采集土壤样品数为:5个采样点×5深度=25个, 进行5点混合, 即将每个深度上的5个采样点的土壤样品进行等量均匀混合, 后最终每个样方内土壤样品数为5个, 每个样地3个样方, 每个样地的土壤样品数为15个。每个样品取样重量200—300 g, 将所取的新鲜土样分为两份, 用无菌自封袋密封, 置于冰盒尽快带回实验室, 并于-20℃下保存。在24 h之内, 将每个样地的同一土层深度的5个土壤样品混合均匀并过直径为2 mm的网筛, 除去根系、石块等杂物。其中一部分风干后用于土壤化学性质的测定;另一部分放入-80℃冰箱保存, 用于土壤酶活性测定和和高通量测序。
1.2.3 土壤化学性质和酶活性测定土壤pH值(1:2.5)采用电位法测定[16]。土壤有机碳(OC)采用重铬酸钾法进行测定;土壤硝态氮和铵态氮采用全自动间断化学分析仪进行测定(CleverChem 380, Germany)。土壤过氧化氢酶的测定采用KMnO4滴定法;其活性以20 min内每g土壤分解的过氧化氢毫克数表示;土壤蔗糖酶的测定采用3, 5-二硝基水杨酸法, 其活性以37℃下培养24 h后1 g土壤中生成的葡萄糖毫克数表示;土壤脲酶的测定采用苯酚钠-次氯酸钠比色法, 其活性以37℃下培养24 h后1 g土壤中生成NH4+-N的毫克数表示;土壤磷酸酶的测定采用磷酸苯二钠比色法, 其活性以37℃下培养24 h后1 g土壤中生成酚的毫克数表示[17]。
1.2.4 样品DNA提取和高通量测序将四个样地的所采取的0—10 cm深度的3个重复土壤样品和10—20 cm深度的3个重复土壤样品均匀混合为一个样品后, 分别称取0.5 g, 用于DNA提取:利用土壤试剂盒(PowerSoil® DNA Isolation试剂盒)按照试剂盒操作步骤提取样本的DNA.将提取的基因组DNA, 利用引物序列520F(AYTGGGYDTAAAGNG)和802R(TACNVGGGTATCTAATCC)对细菌16S rDNA高变区V4区进行扩增;ITS1F(TCCGTAGGTGAACCTGCGG)和ITS2R(GCTGCGTTCTTCATCGATGC)对真菌ITS1区2域进行扩增。利用引物序列PCR反应条件为:98℃预变性30 s;98℃变性30 s;50℃退火30 s;72℃延伸30 s;27个循环;72℃保温5 min, 在4℃下保存。扩增结果进行2%琼脂糖凝胶电泳, 最后将扩增产物进行Illumina MiSeq高通量测序与分析。本研究的测序和生物信息服务在上海派森诺生物科技有限公司完成。
1.3 数据处理与分析微生物群落OTU分析:首先运用QIIME软件识别疑问序列:软件剔除长度小于50 bp的序列以及序列尾部质量值在20以下的碱基, 再对低复杂度的序列进行过滤, 去除预处理后序列中的非扩增区域序列, 并利用USEARCH检查并去除序列中的嵌合体。再利用Uclust软件对符合要求的序列在97%的相似水平上进行OUT聚类, 并采用Greengene数据库对所得序列进行比对分析, 最后以97%的相似性作为阈值划分操作分类单元(OTU)。
微生物群落多样性分析:利用QIIME软件对土壤样品中细菌和真菌的α多样性值进行计算, 包括群落丰富度指数Chao1和ACE, 群落多样性的指数Shanon(H)[18]和Simpson(D)[19]。
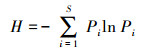
|
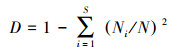
|
式中, S是微生物群落中物种的总数目。Pi为物种i的相对丰度, 即Pi= Ni/N, Ni是调查样方内物种i的植株数量, N是调查样方内所有物种的数量。
统计分析:应用SPSS 22.0软件对不同样地同一土层深度的土壤酶和各化学性质间的显著性差异进行单因素方差分析(one—way analysis)和多重比较分析(LSD), 显著性水平设为P<0.05。应用Canoco 4.5对土壤化学性质、土壤酶活性和微生物群落之间的关系进行冗余分析(RDA)。应用Origin 9.0软件进行图形绘制。
2 结果与分析 2.1 土壤化学性质表 1所示, 4个样地土壤pH在0—50 cm深度都呈现碱性(8.56—9.63), 在0—40 cm深度, 翻耕样地(FG)和荒地(HD)土壤pH无显著差异, 且总体上显著高于免耕(MG)和苜蓿(MX)样地。土壤有机碳含量总体表现为免耕样地最高, 荒地最低, 两个样地土壤有机碳差异具有显著性。且在0—40 cm深度, 苜蓿样地土壤有机碳含量也显著高于荒地。4个样地土壤硝态氮(NO3--N)含量在0—50 cm深度都呈现出免耕和苜蓿样地(6.64—13.34 mg/kg)显著高于翻耕样地和荒地(0.54—7.49 mg/kg)。苜蓿样地表层土壤(0—20 cm)氨态氮(NH4+-N)含量显著高于其他样地, 翻耕氨态氮含量较低。而在20—40 cm深度, 免耕样地氨态氮含量最高, 荒地最低。
| 深度 Depth/cm |
样地 Sample |
pH | 有机碳 Organic carbon/(mg/g) |
硝态氮 Nitrate nitrogen/(mg/kg) |
铵态氮 Ammonium nitrogen/(mg/kg) |
| 0—10 | MG | 9.05±0.03b | 5.85±0.54a | 12.70±0.02a | 2.95±1.29bc |
| FG | 9.44±0.07a | 4.66±0.90b | 0.93±0.20c | 0.82±0.08c | |
| MX | 9.13±0.08b | 4.53±0.07b | 13.34±0.46a | 6.29±0.01a | |
| HD | 9.42±0.06a | 3.83±0.03c | 7.49±0.13b | 4.93±0.04b | |
| 10—20 | MG | 9.07±0.04a | 4.83±0.20a | 8.01±1.14b | 1.64±0.09b |
| FG | 9.42±0.09a | 3.63±0.52c | 0.76±0.10c | 1.45±0.10bc | |
| MX | 8.82±0.05b | 4.32±0.09b | 11.16±0.95a | 2.53±0.31a | |
| HD | 9.60±0.10a | 3.38±0.04c | 0.84±0.37c | 1.38±0.06c | |
| 20—30 | MG | 9.09±0.01b | 3.54±0.15a | 8.97±1.08b | 3.11±0.06a |
| FG | 9.56±0.05a | 3.56±0.45a | 1.08±0.04c | 2.15±0.05b | |
| MX | 8.56±0.44b | 3.58±0.31a | 11.61±0.32a | 1.98±0.21b | |
| HD | 9.57±0.01a | 3.00±0.08b | 1.45±0.58c | 1.31±0.17c | |
| 30—40 | MG | 9.01±0.01 b | 3.91±0.31ab | 8.37±1.21b | 2.74±0.74a |
| FG | 9.38±0.07a | 3.72±0.01b | 0.67±0.23c | 2.50±1.21ab | |
| MX | 8.89±0.13b | 3.30±0.04ac | 9.07±0.56a | 1.86±0.06ab | |
| HD | 9.48±0.12a | 2.56±0.13d | 0.70±0.17c | 1.66±0.02b | |
| 40—50 | MG | 9.18±0.04b | 4.41±0.13a | 8.61±1.23a | 1.02±0.12c |
| FG | 9.36±0.10b | 3.73±0.24b | 0.54±0.05b | 2.11±0.21a | |
| MX | 9.08±0.06c | 2.52±0.10c | 6.64±0.94a | 1.41±0.24b | |
| HD | 9.63±0.03a | 2.47±0.00c | 0.70±0.18b | 0.84±0.22c | |
| MG:免耕No tillage land;FG:翻耕Conventional tillage land;MX:苜蓿Alfalfa land;HD:荒地Uncultivated land。不同字母表示不同样地同一土层之间P < 0.05水平上差异显著 | |||||
土壤酶主要来源于动植物及微生物细胞的分泌及其残体的腐解。在0—50 cm土层深度, 4个样地的土壤过氧化氢酶随深度的增加变化趋势基本一致, 即都随着土层深度的增加而降低。苜蓿样地0—50 cm深度土壤过氧化氢酶活性变化范围为2.36—3.95 mg g-1h-1, 显著高于其他样地, 而荒地土壤过氧化氢酶活性变化范围为0.62—0.97 mg g-1h-1, 显著低于其他样地。在0—30 cm土层, 不同土地恢复模式对土壤蔗糖酶活性影响一致, 免耕样地土壤蔗糖酶活性显著高于其他样地, 表层0—10 cm酶活性值最高(860 mg kg-1h-1), ,而荒地蔗糖酶活性最低;而30—40 cm, 蔗糖酶活性在四个样地之间呈现显著性差异, 表现为:翻耕>苜蓿>免耕>荒地。4种土地恢复模式下, 土壤脲酶基本表现为0—10 cm深度活性最高, 苜蓿样地的脲酶活性在0—10 cm和20—30 cm显著高于其他样地, 而在10—20 cm和30—50 cm, 翻耕样地脲酶活性显著高于其他样地。总体上, 苜蓿和翻耕样地脲酶活性显著高于其他两样地。不同土地管理方式对土壤磷酸酶活性的影响, 表现为0—10 cm土层, 翻耕样地显著低于其他样地;10—50 cm土层, 总体表现为免耕样地土壤磷酸酶活性最高, 荒地最低;其中在10—30 cm和40—50 cm, 免耕样地磷酸酶活性显著高于其他样地(图 1)。

|
| 图 1 不同土地管理方式下土壤酶活性 Fig. 1 Soil enzyme activities under different management practices MG:免耕No tillage land;FG:翻耕Conventional tillage land;MX:苜蓿Alfalfa land;HD:荒地Uncultivated land。图中不同字母表示同一土层不同样地之间P < 0.05差异显著 |
4个样地土壤微生物在门水平上细菌相对丰度大于1%的共有9个门, 分别是变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)、浮霉菌门(Planctomycetes)芽单胞菌门(Gemmmatimonadetes)、疣微菌门(Verrucomicrobia)、硝化螺旋菌门(Nitrospirae)和拟杆菌门(Bacteroidetes)。其中变形菌门所占的相对丰度最大(21.4%—23.0%), 其次是放线菌门和酸杆菌门, 相对丰度分别为18.7%—23.0%和17.5%—19.9%。4个样地的细菌在门水平上的群落分布存在一定的差异。对丰度最大的变形菌门在4个样地的相对丰度表现为:翻耕(25%)>荒地(23%)>苜蓿(22%)>免耕(21%);放线菌门、酸杆菌门和绿弯菌门在免耕样地相对丰度都最大(图 2)。4个样地在门水平上的优势真菌相对丰度大于1%的共有11个门, 主要包括子囊菌门(Ascomycota)、接合菌门(Zygomycota)和担子菌门(Basidiomycota), 相对丰度分别为61.4%—72.8%、1.3%—18.6%和4.6 %—14.1%, 未命名的菌门占0.7%—14.8%(图 3)。相对丰度最大的子囊菌门在4个样地的相对丰度表现为:苜蓿>免耕>翻耕>荒地。接合菌门在翻耕样地的相对丰度最大(19%), 荒地最小(1.3%);担子菌门在荒地的相对丰度最大(21%)(图 3)。

|
| 图 2 门水平细菌群落结构及分布 Fig. 2 Bacterial communities and distribution at the phylum level |

|
| 图 3 门水平真菌群落结构及分布 Fig. 3 Fungal communities and distribution at the phylum level |
对细菌群落的变形菌门(A)、放线菌门(B)、酸杆菌门(C)和绿弯菌门(D)以及真菌群落的子囊菌门(E)和担子菌门(F)的菌群在纲水平上进行分析(图 4), 4个样地中α-变形菌纲(Alphaproteobacteria)是变形菌门中相对丰度最大的菌群, 约占比例50.9%—64.0%, 其次是β-变形菌纲、γ-变形菌纲和δ-变形菌纲, 约占比例分别为13.8%—22.2%、9.6%—16.9%和9.8%—12.3%(图 4A)。组成放线菌门的菌群有7个纲, 相对丰度较高的为放线菌纲、嗜热菌纲、酸微菌纲和MB-A2-108, 放线菌纲和嗜热菌纲为优势放线菌, 所占比例分别为38.5%—53.8%和25.2%—35.9%, 而酸微菌纲和MB-A2-108所占比例较低(图 4B)。酸杆菌门中相对丰度较高的菌群是6-酸杆菌纲和Chloracidobacteria, 所占比例为60.2%—64.1%和25.2%—29.9%(图 4C)。绿弯菌门中主要有厌氧绳菌纲(Anaerolineae)、绿弯菌纲(Chloroflexi)、Ellin6529和Gitt-GS-136, 其中厌氧绳菌纲、绿弯菌纲和Ellin6529相对丰度较高, 分别占18.5%—30.0%、21.4%—29.1%和18.8%—37.6%(图 4D)。真菌群落的子囊菌门中古菌根菌纲(Archaeorhizomycetes)所占相对丰度比例最高, 4个样地所占比例基本相同, 其次为座囊菌纲(Dothideomycetes)、散囊菌纲(Eurotiomycetes)、粪壳菌纲(Sordariomycetes), 所占比例分别为3.37%—6.90%、1.60%—2.24%和17.9%—30.8%(图 4E)。组成担子菌门的菌纲主要有伞菌纲(Agaricomycetes)、银耳纲(Tremellomycetes), 以及未确定和命名分类的菌纲, 其中伞菌纲和银耳纲相对丰度较高, 分别为33.4%—76.2%和18.8%—59.7%(图 4F), 另外一个相对丰度较高的接合菌门, 其纲水平的菌种未确定。

|
| 图 4 纲水平土壤优势菌群落结构及分布 Fig. 4 Soil dominant microbes communities and distribution at the class level |
基于序列在97%的相似水平上进行OUT聚类, 4个样地微生物α多样性见表 2。Chao1和ACE用来衡量群落丰富度指数, 指数越大, 其丰富度越高, 而Shannon指数值越高, 表明群落的多样性越高。表 2所示, 苜蓿样地土壤细菌群落丰富度最高, Chao1和ACE指数分别为8064.43和8553.70, 其次为免耕样地, Chao1和ACE指数分别为7668.94和8311.15, 荒地最低(Chao1为5812.21, ACE为5911.68)。苜蓿样地土壤细菌群落Shannon指数最高(10.50), 说明苜蓿样地土壤细菌群落多样性最高, 其次为荒地(10.45)和免耕(10.42), 翻耕样地土壤细菌群落多样性最低, Shannon指数为10.32(表 2)。对真菌群落进行分析, 免耕样地土壤真菌群落丰富度最高, Chao1和ACE指数分别为926.00和1060.44, 其次是苜蓿和翻耕样地, 荒地最低。免耕样地的Shannon指数最高(6.92), 表明免耕样地土壤真菌群落多样性最高, 其次为苜蓿和荒地, 翻耕样地土壤真菌群落多样性最低, Shannon指数为6.00(表 2)。总体上, 苜蓿和免耕样地土壤细菌和真菌群落丰富度和多样性都较高, 翻耕样地土壤细菌和真菌群落丰富度较高, 但多样性较低, 而荒地土壤细菌和真菌群落丰富度较低, 但多样性较高。
| 样地Samples | 种类 Species |
序列数 Reads |
分类单元 Operational taxonomic units |
Chao1指数 Chao1Index |
ACE指数 Abundance-based coverage estimator index |
Shannon指数 Shannon index |
| MG | 细菌 | 42244 | 5719 | 7668.94 | 8311.15 | 10.42 |
| 真菌 | 45449 | 1074 | 926.00 | 1060.44 | 6.92 | |
| FG | 细菌 | 52179 | 5633 | 7231.22 | 7712.57 | 10.32 |
| 真菌 | 44989 | 760 | 677.00 | 759.66 | 6.00 | |
| MX | 细菌 | 47341 | 6040 | 8064.43 | 8553.70 | 10.50 |
| 真菌 | 44528 | 779 | 732.00 | 843.34 | 6.43 | |
| HD | 细菌 | 49477 | 4960 | 5812.21 | 5911.68 | 10.45 |
| 真菌 | 32292 | 449 | 417.00 | 417.00 | 6.35 |
对土壤微生物优势细菌和真菌群落相对丰度和土壤环境因子进行RDA冗余分析(图 5), 图中大写字母多对应的微生物种类见表 3, 分析结果显示, 细菌丰度分布在第一轴和第二轴累计解释变量分别达到74.2%和92.4%, 对于真菌丰度分布在第一轴和第二轴累计解释变量分别达到67.1%和95.3%。土壤细菌群落优势菌群的相对丰度与土壤环境因子冗余分析, 结果(图 5)显示酸杆菌门(Acidobacteria)主要集中在排序轴左侧, 与pH呈显著正相关关系, 与硝态氮、铵态氮和过氧化氢酶呈负相关关系;变形菌门中的主要变形菌纲B1与硝态氮、铵态氮和过氧化氢酶呈正相关关系, 而与pH呈负相关关系;放线菌门主要集中在排序轴右侧, 其与硝态氮、铵态氮和过氧化氢酶显著正相关关系;绿弯菌门中的D1、D3与pH呈正相关关系, 而D2、D4与硝态氮、铵态氮和过氧化氢酶呈负相关关系。土壤真菌群落优势菌群的相对丰度与土壤环境因子冗余分析, 结果(图 5)显示, 子囊菌门(Ascomycota)的优势菌集中在排序轴右侧, 与硝态氮、铵态氮和过氧化氢酶呈正相关关系, 而与脲酶和pH呈负相关关系, 其中E2、E4菌纲还与蔗糖酶和有机碳呈正相关关系;担子菌门(Basidiomycota)的优势菌菌纲F1与硝态氮、铵态氮、过氧化氢酶、蔗糖酶和有机碳呈正相关关系, 与脲酶和pH呈负相关关系, F2与脲酶和pH呈正相关关系, 与硝态氮、铵态氮和过氧化氢酶呈负相关关系;接合菌门(Zygomycota)与pH、脲酶和有机碳呈正相关关系, 而与硝态氮、铵态氮和过氧化氢酶呈负相关关系。

| 编号Number | 种类Species |
| 酸杆菌门Acidobacteria | |
| A1 | 6-酸杆菌刚Acidobacteria-6 |
| A2 | Chloracidobacteria |
| A3 | Sva0725 |
| A4 | iii1-8 |
| 放线菌门Actinobacteria | |
| C1 | 放线菌纲Actinobacteria |
| C2 | 嗜热菌纲Thermoleophilia |
| C3 | 酸微菌纲Acidimicrobiia |
| C4 | MB-A2-108 |
| 子囊菌门Ascomycota | |
| E1 | 古菌根菌纲Archaeorhizomycetes |
| E2 | 座囊菌纲Dothideomycetes |
| E3 | 散囊菌纲Eurotiomycetes |
| E4 | 粪壳菌纲Sordariomycetes |
| 变形菌门Alphaproteobacteria | |
| B1 | γ-变形菌纲Alphaproteobacteria |
| B2 | β-变形菌纲Betaproteobacteria |
| B3 | δ-变形菌纲Deltaproteobacteria |
| B4 | γ-变形菌纲Gammaproteobacteria |
| 绿弯菌门Chloroflexi | |
| D1 | 厌绳菌纲Anaerolineae |
| D2 | Ellin6529 |
| D3 | 绿弯菌纲Chloroflexi |
| D4 | Gitt-GS-136 |
| 担子菌门Basidiomycota | |
| F1 | 伞菌纲Agaricomycetes |
| F2 | 银耳纲Tremellomycetes |
| G | 接合菌门Zygomycota |
细菌群落多样性指数与环境因子的相关性矩阵(图 6)表明, 土壤细菌群落多样性与土壤环境因子具有一定相关性。土壤细菌Chao1指数与ACE指数、磷酸酶显著正相关, 与脲酶和过氧化氢酶活性显著负相关;土壤真菌ACE指数与磷酸酶活性显著相关, 与NH4+-N含量、脲酶和过氧化氢酶活性显著负相关。土壤细菌群落Shannon指数与磷酸酶、蔗糖酶、铵态氮均匀度以及土壤NH4+-N、NO3--N含量呈显著正相关, 与pH呈显著负相关。真菌群落多样性指数与环境因子的相关性矩阵显示, 土壤真菌Chao1指数与ACE指数、Shannon指数、过氧化氢酶和蔗糖酶活性、有机碳呈显著正相关, 与pH显著负相关;土壤真菌ACE指数与过氧化氢酶和蔗糖酶活性、有机碳呈显著正相关, 与pH显著负相关(图 7), 土壤真菌群落Shannon指数与NH4+-N含量、脲酶和过氧化氢酶活性显著负相关土壤细菌群落Shannon指数与磷酸酶、蔗糖酶、NO3--N和有机碳含量呈显著正相关, 与pH呈显著负相关(图 7)。

|
| 图 6 细菌群落多样性指数与环境因子的相关性 Fig. 6 Correlations between diversity index of fungus communities and environmental factors Chao1:Chao1指数Chao1 Index;ACE:ACE指数Abundance-based Coverage Estimator index;Shannon:Shannon多样性Shannon diversity;过氧化氢酶活性:Catalase activity;磷酸酶活性:Phosohatase activity;脲酶活性:Urease activity;蔗糖酶活性:Invertase activity;NH4+-N:铵态氮Ammonium nitrogen;NO3--N硝态氮Nitrate nitrogen;SOM:有机质Organic matter;pH值:pH value;* P<0.05, **P<0.01.四周数值表示环境因子的测量值, 其中纵坐标每组数值表示所在行环境因子的测量值, 横坐标每组数值表示所在列环境因子的测量值, 图中数值表示该数值所在行和列相对应的环境因子间的相关系数 |

|
| 图 7 真菌群落多样性指数与环境因子的相关性 Fig. 7 Correlations between diversity index of bacterial communities and environmental factors |
土壤环境因子是植物和微生物生存的载体[20], 由于不同土地管理方式下, 其地面覆被以及土壤收到的扰动(耕作和免耕)情况不同, 对土壤养分的富集和再分配作用, 以及减少水土流失引起的养分流失的作用不同, 进而会对土壤养分和酶活性及其在土壤剖面上的分布产生影响[21]。尤其是土壤有机碳氮含量主要取决于生物固氮及植被凋落物的归还量[22]有研究植被生长年限表层土壤养分含量和酶活性都高于深层土壤, 也有研究表明, 不同土地管理方式显著影响土壤有机碳和氮的含量[23]。本研究表明土壤有效氮(NH4+-N和NO3--N)和土壤有机碳含量和酶活性受土地管理方式和深度的共同影响, 总体上, 4种土地利用管理方式土壤有机碳、铵态氮和硝态氮含量随土壤深度的增加而降低, 4种土壤酶活性也呈同样变化趋势(表 1;图 1), 其中苜蓿和免耕样地土壤铵态氮、硝态氮和有机碳含量显著高于翻耕和荒地, 这是由于苜蓿和免耕样地土壤未受到扰动, 此外, 苜蓿为豆科植物, 根瘤菌可固定土壤氮素[24], 且地上和地下部分生物量较高, 而免耕样地由于地上作物秸秆和地下根茬还田量较高, 并且由于地表秸秆覆盖降低了土壤侵蚀, 从而提高了土壤碳氮含量[22]。这也表明植被恢复和无土扰动可改善土壤质量[25-26], 例如Wang等[27]在黄土高原地区研究植被恢复对土壤有机碳的影响, 发现草地能明显增加SOC含量。
土壤酶参与降解有机碳的分解和氮磷等元素的循环, 其活性可表征自然或人为活动对土壤的干扰程度[28], 并且影响土壤酶活性的因素主要是输入土壤中的有机物量, 以及土壤微生物的活性和数量[20]。本研究中苜蓿和免耕样地土壤过氧化氢酶、蔗糖酶、磷酸酶活性都较高, 而翻耕和荒地较低, 这是由于免耕和苜蓿样地凋落物较多且根系密集, 使土壤中供微生物吸收利用的物质源增加[17], 且由于凋落物的C/N较高, 促进土壤中异样微生物的组成和活性, 增强微生物代谢能力[24, 28-29]。脲酶活性与土壤氮素相关, 且土壤脲酶为好气性水解酶[30-31], 可将土壤中的有机化合物尿素分解转化为植物能利用的有效氮[32]。本研究中脲酶活性在苜蓿和翻耕样地显著高于免耕和对照样地(荒地), 这是由于苜蓿样地土壤氮素含量高[24], 而翻耕样地土壤通气性较好[33-34]。
3.2 土壤微生物群落多样性及土壤环境因子的关系土壤微生物的群落分布与其所处的土壤环境有关, 微生物对土壤环境因子的适应能力决定了其群落分布的多样性[35-36]。本研究中4种土地管理方式下土壤微生物群落在门和纲两个分类层次上的细菌和真菌菌群分布特征结果显示, 细菌丰度较大的为变形菌门、放线菌门和酸杆菌门, 真菌的子囊菌门丰度最大, 其中苜蓿和免耕样地土壤优势菌群(细菌和真菌)群落丰富度和多样性都较高, 主要是苜蓿和免耕样地土壤的碱性弱、有机碳和有效氮(NH4+-N和NO3--N)含量较高[37-38]。本研究显示, 土壤pH、硝态氮和铵态氮对细菌群落影响较大, pH和有机碳对真菌群落影响最大, 可见土壤pH值是土壤微生物的重要调节因子[18, 39-40]。由于真菌为慢周转方式, 偏好低营养、难分解的有机质, 对有机质的利用率高, 因此有机碳对真菌群落的影响较高[41]。RDA分析结果表明, 过氧化氢酶对细菌群落影响较大, 而蔗糖酶、脲酶和过氧化氢酶对真菌群落的影响都较大, 真菌对土壤酶活性的贡献率显著高于细菌群落。这与马文文[41]等的研究结果相符。
4 结论(1) 不同土地管理方式显著影响土壤有机碳、有效氮(NH4+-N、NO3--N)和土壤酶的活性, 并且随着土层深度的增加而减少, 其中苜蓿和免耕样地土壤有机碳、铵态氮和硝态氮含量显著高于翻耕和对照样地(荒地), 且苜蓿和免耕样地过氧化氢酶、蔗糖酶和磷酸酶活性较高, 而对照荒地过氧化氢酶活性最低;而脲酶活性在苜蓿和翻耕样地显著高于免耕和对照样地(荒地)。总体上, 苜蓿和免耕2种土地管理方式可显著提高表层土壤有机碳、铵态氮和硝态氮含量并增加土壤酶活性。
(2) 不同土地管理方式导致土壤环境差异, 进而影响土壤微生物的群落丰富度及多样性, 4种土地管理方式共有9个细菌门和11个真菌门, 细菌丰度较大的为变形菌门、放线菌门和酸杆菌门, 真菌的子囊菌门丰度最大;苜蓿和免耕样地土壤细菌和真菌群落丰富度和多样性都较高, 翻耕样地土壤细菌和真菌群落丰富度较高, 但多样性较低, 而荒地土壤细菌和真菌群落丰富度较低, 但多样性较高。
(3) 在纲水平上对土壤细菌和真菌群落组成进行RDA分析, 结果表明土壤pH、硝态氮和铵态氮对细菌群落影响较大, pH和有机碳对真菌群落影响最大;过氧化氢酶对细菌群落影响较大, 而蔗糖酶、脲酶和过氧化氢酶对真菌群落的影响都较大, 真菌对土壤酶活性的贡献率显著高于细菌群落。因此, 土壤细菌和真菌群落组成受到外部环境的显著影响, 如土地管理方式、pH、NH4+-N和NO3--N含量、有机碳碳含量、过氧化氢酶活性、蔗糖酶活性、脲酶活性。
(4) 综上, 本研究通过对晋西北丘陵区不同土地管理方式的土壤pH和碳氮含量、酶活性以及微生物细菌和真菌群落组成和结构特征进行系统的分析, 表明在所研究的四种土地管理方式中, 种植苜蓿和免耕是适用于该区的土地管理方式, 地方政府和当地农民可适当选用苜蓿田和保护性耕作田, 以保持农业的持续发展和生态环境安全。
| [1] |
王青杵, 王改玲, 石生新, 庄丽, 孙泰森. 晋北黄土丘陵区不同人工植被对水土流失和土壤水分含量的影响. 水土保持学报, 2012, 26(2): 71-74, 79-79. |
| [2] |
王雅, 刘爽, 郭晋丽, 刘兵兵. 黄土高原不同植被类型对土壤养分、酶活性及微生物的影响. 水土保持通报, 2018, 38(1): 62-68. |
| [3] |
Chapman S K, Newman G S. Biodiversity at the plant-soil interface:microbial abundance and community structure respond to litter mixing. Oecologia, 2010, 162(3): 763-769. DOI:10.1007/s00442-009-1498-3 |
| [4] |
陈龙, 姚拓, 柴晓虹, 王理德, 马剑平, 孙广正, 安雅君, 李发明. 石羊河中下游不同退耕年限次生草地土壤无机氮、酶及微生物量冬季动态研究. 草地学报, 2016, 24(2): 330-336. |
| [5] |
杨万勤, 王开运. 土壤酶研究动态与展望. 应用与环境生物学报, 2002, 8(5): 564-570. |
| [6] |
Boerner J R E, Brinkman J A, Smith A. Seasonal variations in enzyme activity and organic carbon in soil of a burned and unburned hardwood forest. Soil Biology and Biochemistry, 2005, 37(8): 1419-1426. DOI:10.1016/j.soilbio.2004.12.012 |
| [7] |
Balota E L, Kanashiro M, Filho A C, AndradeI D S, DickⅡ R P. Soil enzyme activities under long-term tillage and crop rotation systems in subtropical agro-ecosystems. Brazilian Journal of Microbiology, 2004, 35(4): 300-306. DOI:10.1590/S1517-83822004000300006 |
| [8] |
Zhang Y M, Zhou G Y, Wu N, Bao K K. Soil enzyme activity changes in different-aged spruce forests of the eastern Qinghai-Tibetan plateau. Pedosphere, 2004, 14(3): 305-312. |
| [9] |
BandickA K, Dick R P. Field management effects on soil enzyme activities. Soil Biology and Biochemistry, 1999, 31(11): 1471-1479. DOI:10.1016/S0038-0717(99)00051-6 |
| [10] |
Kandeler E, Stemmer M, Klimanek E M. Response of soil microbial biomass, urease and xylanase within particle size fractions to long-term soil management. Soil Biology and Biochemistry, 1999, 31(2): 261-273. DOI:10.1016/S0038-0717(98)00115-1 |
| [11] |
Williams MA, Rice C W, OwensbyC E. Carbon dynamics and microbial activity in tallgrass prairie exposed to elevated CO2 for 8 years. Plant and Soil, 2000, 227(1/2): 127-137. DOI:10.1023/A:1026590001307 |
| [12] |
曹瑞, 吴福忠, 杨万勤, 徐振锋, 谭波, 王滨, 李俊, 常晨晖. 海拔对高山峡谷区土壤微生物生物量和酶活性的影响. 应用生态学报, 2016, 27(4): 1257-1264. |
| [13] |
Gómez-Sagasti M T, Alkorta I, Becerril J M, Epelde L, Anza M, Garbisu C. Microbial monitoring of the recovery of soil qualityduring heavy metal phytoremediation. Water, Air, &Soil Pollution, 2012, 223(6): 3249-3262. |
| [14] |
Van Der Heijden M G A, Bardgett R D, Van Straalen N M. The unseen majority:soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecology Letters, 2008, 11(3): 296-310. DOI:10.1111/ele.2008.11.issue-3 |
| [15] |
付东磊, 刘梦云, 刘林, 张琨, 左进香. 黄土高原不同土壤类型有机碳密度与储量特征. 干旱区研究, 2014, 31(1): 44-50. |
| [16] |
鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科学出版社, 1999.
|
| [17] |
周礼恺. 土壤酶学. 北京: 科学出版社, 1987.
|
| [18] |
乔沙沙, 周永娜, 柴宝峰, 贾彤, 李毳. 关帝山森林土壤真菌群落结构与遗传多样性特征. 环境科学, 2017, 38(6): 2502-2512. |
| [19] |
德吉, 吴小妮, 索朗德庆, 多吉卓玛, 韦恩喜. 拉鲁湿地土壤细菌多样性初探. 西藏大学学报:自然科学版, 2014, 29(2): 44-50. |
| [20] |
王兵, 刘国彬, 薛萐. 退耕地养分和微生物量对土壤酶活性的影响. 中国环境科学, 2010, 30(10): 1375-1382. |
| [21] |
Sever H, Makineci E. Soil organic carbon and nitrogen accumulation on coal mine spoils reclaimed with maritime pine (PinuspinasterAiton) in Agacli-Istanbul. Environmental Monitoring and Assessment, 2009, 155(1/4): 273-280. |
| [22] |
熊汉锋, 王运华. 梁子湖湿地土壤养分的空间异质性. 植物营养与肥料学报, 2005, 11(5): 584-589. |
| [23] |
Zhao Z Q, Shahrour I, Bai Z K, Fan W X, Feng L R, Li H F. Soils development in opencast coal mine spoils reclaimed for 1-13 years in the West-Northern Loess Plateau of China. European Journal of Soil Biology, 2013, 55: 40-46. DOI:10.1016/j.ejsobi.2012.08.006 |
| [24] |
Yuan Z Q, Yu K L, Epstein H, Fang C, Li J T, Liu Q Q, Liu X W, Gao W J, Li F M. Effects of legume species introduction on vegetation and soil nutrient development on abandoned croplands in a semi-arid environment on the Loess Plateau, China. Science of the Total Environment, 2016, 541: 692-700. DOI:10.1016/j.scitotenv.2015.09.108 |
| [25] |
Barbera V, Poma I, Gristina L, Novara A, Egli M. Long-term cropping systems and tillage management effects on soil organic carbon stock and steady state level of C sequestration rates in a semiarid environment. Land Degradation & Development, 2012, 23(1): 82-91. |
| [26] |
Jiao F, WenZ M, An S S. Changes in soil properties across a chronosequence of vegetation restoration on the Loess Plateau of China. CATENA, 2011, 86(2): 110-116. DOI:10.1016/j.catena.2011.03.001 |
| [27] |
WangY F, FuB J, Lü Y H, Chen L D. Effects of vegetation restoration on soil organic carbon sequestration at multiple scales in semi-arid Loess Plateau, China. CATENA, 2011, 85(1): 58-66. DOI:10.1016/j.catena.2010.12.003 |
| [28] |
龙健, 黄昌勇, 滕应, 姚槐应. 矿区废弃地土壤微生物及其生化活性. 生态学报, 2003, 23(3): 496-503. |
| [29] |
戴全厚, 刘国彬, 姜峻, 薛楚, 翟胜. 黄土丘陵区不同植被恢复模式对土壤酶活性的影响. 中国农学通报, 2008, 24(9): 429-434. |
| [30] |
安韶山, 黄懿梅, 郑粉莉. 黄土丘陵区草地土壤脲酶活性特征及其与土壤性质的关系. 草地学报, 2005, 13(3): 233-237. |
| [31] |
刘家雄, 金陈斌, 张静, 汤珧华. 古树土壤酶活性与土壤肥力的关系. 安徽农业科学, 2018, 46(12): 146-148. |
| [32] |
Dequiedt S, Thioulouse J, Jolivet C, Saby N P A, Lelievre M, Maron P A, Martin M P, Prévost-Bouré N C, Toutain B, Arrouays D, Lemanceau P, Ranjard L. Biogeographical patterns of soil bacterial communities. Environmental Microbiology Reports, 2009, 1(4): 251-255. DOI:10.1111/emi4.2009.1.issue-4 |
| [33] |
袁建国, 李变梅, 万俊梅, 任亮仙. 晋西北干旱地区保护性耕作的研究. 山西农业大学学报:自然科学版, 2004, 24(4): 347-350. |
| [34] |
张德喜, 吴卿. 不同耕作方式对农田土壤养分含量及土壤酶活性的影响. 江苏农业科学, 2018, 46(11): 234-237. |
| [35] |
Ranjard L, Dequiedt S, Jolivet C, Saby N P A, Thioulouse J, Harmand J, Loisel P, Rapaport A, Fall S, Simonet P, Joffre R, Bouré N C P, Maron P A, Mougel C, Martin M P, Toutain B, Arrouays D, Lemanceau P. Biogeography of soil microbial communities:a review and a description of the ongoing French national initiative. Agronomy for Sustainable Development, 2010, 30(2): 359-365. DOI:10.1051/agro/2009033 |
| [36] |
陈法霖, 张凯, 郑华, 林学强, 欧阳志云, 屠乃美. PCR-DGGE技术解析针叶和阔叶凋落物混合分解对土壤微生物群落结构的影响. 应用与环境生物学报, 2011, 17(2): 145-150. |
| [37] |
Sinsabaugh RL, Lauber CL, Weintraub MN, Ahmed B, Allison SD, Crenshaw C, Contosta AR, Cusack D, Frey S, Gallo ME, Gartner TB, Hobbie SE, Holland K, Keeler BL, Powers JS, Stursova M, Takacs-Vesbach C, Waldrop MP, Wallenstein MD, Zak DR, Zeglin LH. Stoichiometry of soil enzyme activity at global scale. Ecology Letters, 2008, 11(11): 1252-1264. DOI:10.1111/ele.2008.11.issue-11 |
| [38] |
唐仕姗, 杨万勤, 何伟, 王海鹏, 熊莉, 聂富育, 徐振锋. 川西亚高山3种优势林木不同径级根系分解及木质素、纤维素降解特征. 应用与环境生物学报, 2015, 21(4): 754-761. |
| [39] |
Blagodatskay E V, Anderson T H A. Interactive effects of pH and substrate quality on the fungal-to-bacterial ratio and qCO2of microbial communities in forest soils. Soil Biology and Biochemistry, 1998, 30(10/11): 1269-1274. |
| [40] |
Lauber C L, Hamady M, Knight R, Fierer N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Applied and Environmental Microbiology, 2009, 75(15): 5111-5120. DOI:10.1128/AEM.00335-09 |
| [41] |
马文文, 姚拓, 靳鹏, 王国基, 张玉霞. 荒漠草原2种植物群落土壤微生物及土壤酶特征. 中国沙漠, 2014, 34(1): 176-183. |