 2019, Vol. 39
2019, Vol. 39文章信息
- 王晓荣, 曾立雄, 雷蕾, 潘磊, 肖文发
- WANG Xiaorong, ZENG Lixiong, LEI Lei, PAN Lei, XIAO Wenfa
- 抚育择伐对马尾松林主要树种空间分布格局及其关联性的短期影响
- Short-term effects of selective cutting on the spatial distribution and association of dominant tree species in Pinus massoniana stands
- 生态学报. 2019, 39(12): 4421-4431
- Acta Ecologica Sinica. 2019, 39(12): 4421-4431
- http://dx.doi.org/10.5846/stxb201801230177
-
文章历史
- 收稿日期: 2018-01-23
- 网络出版日期: 2019-04-01





2. 湖北省林业科学研究院, 武汉 430075;
3. 南京林业大学南方现代林业协同创新中心, 南京 210037
2. Forestry Institute of Hubei Province, Wuhan 430075, China;
3. Co-Innovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing 210037, China
群落结构由植物种类组成及其空间分布格局所决定, 物种间不同的空间关系会导致群落结构存在差异[1], 直接影响不同层次树种的空间分布格局和更新演替机制[2]。关于植物种群空间结构的研究多集中在未受干扰的自然林分, 重点研究自然群落结构形成和动态变化[3]、组配方式[4-5]、物种共存或竞争[6-7]、生物多样性维持等[8-9], 但对人为采伐干扰下的群落空间结构变化和种间相关性等方面研究明显不足[10-11]。
抚育择伐作为森林经营中重要的人为干扰方式, 往往会改变森林的物种组成和群落空间结构[12], 造成林内温度、光照、物种竞争、更新状况、特定生境条件和土壤等生态系统结构要素的改变[13-14]。已有研究发现, 适度择伐可促进森林结构优化, 对植物群落生长和更新演替产生有利影响[12, 15], 但以往经营管理更多关注择伐密度、择伐强度、次数以及采伐周期, 对采伐物种类型、种间关系等林分结构的改变考虑较少[10-11, 16]。同时, 不同择伐方式对森林结构的影响会随研究区域或树种差异而不同[17], 使得抚育择伐方式对森林结构变化的影响一直未有定论[14]。因此, 明确典型森林群落物种空间分布格局和种间关系对抚育择伐干扰的响应, 有助于揭示人为经营干扰对森林群落特性、稳定性和演替规律等的影响机制[4, 18-19], 为合理开展森林经营管理提供科学依据。
目前, 我国人工林面积全球最大, 根据第八次全国森林资源清查, 面积已达0.693亿hm2[20]。马尾松(Pinus massoniana)是我国亚热带特有乡土树种, 适应性强、耐干旱与瘠薄, 广泛分布于秦岭、淮河以南[21], 也是三峡库区分布面积最大和最主要的林分类型, 占库区森林面积34%[22]。以往马尾松林主要利用方式和管理措施包括采伐大径材马尾松、砍伐杂木以及林下植被抚育等3种模式, 长期受经营水平粗放和人为干扰强烈等多种因素的影响, 库区马尾松林分质量、稳定性及生态防护功能低下等问题十分突出。监测常规抚育择伐措施下马尾松林分结构组成特征及种间功能联系, 追踪抚育择伐后森林空间格局形成的生态学过程, 以确定合理的抚育择伐策略, 可促进林分结构优化发展。因此, 本研究以三峡库区马尾松林为研究对象, 模拟以往马尾松利用和管理措施, 采用未择伐、伐除优势马尾松、伐除非马尾松、除灌等方式[23]建立固定监测样地, 利用点格局法分析抚育择伐3年后马尾松林中主要树种结构、空间分布和种间关联性, 探讨不同抚育择伐方式对马尾松林分组成和结构的短期影响, 为三峡库区马尾松森林结构优化和林分质量提升以及制定合理的森林经营方案提供参考。
1 研究区概况研究地点位于湖北省宜昌市秭归县九岭头林场(30°59′—31°05′N, 110°45′—110°48′E), 海拔156—2030.6 m, 森林覆盖率95.2%。属于亚热带大陆性季风气候, 年均气温17.9℃, 年降水量1000—1250 mm, 多集中在4—9月份, 年日照时数1619.6 h, 无霜天260 d。土壤类型主要以黄壤、黄棕壤为主。研究区内马尾松林为20世纪70年代飞播造林, 长期以来人为干扰极少, 经过自然更新演替, 形成以马尾松为优势树种, 伴生有少量光皮桦(Betula luminifera)、漆树(Toxicodendron vernicifluum)、茅栗(Castanea seguinii)、盐肤木(Rhus chinensis)等的松阔混交林。灌木树种主要有绿叶胡枝子(Lespedeza buergeri)、桦叶荚蒾(Viburnum betulifolium)、宜昌荚蒾(Viburnum erosum)等, 草本以中日金星蕨(Parathelypteris nipponica)、狗脊(Cibotium barometz)、鱼腥草(Heartleaf Houttuynia)、野青茅(Deyeuxia arundinacea)、细梗苔草(Carex teinogyna)为主。
2 研究方法 2.1 样地设置2013年9月, 选择生长在同一坡面且林相完整的马尾松林, 按照未择伐(Non-cutting, NC)、伐除优势马尾松(Dominant Pinus massoniana-cutting, DPMC)、伐除非马尾松(No Pinus massoniana-cutting, NPMC)、除灌(Shrubs-cutting, SC)等4种管理措施, 各设置1块面积为50 m×50 m的正方形样地, 样地间相距20—30 m(当样地面积≥2500 m2时能够较真实的反映林木的空间分布格局[24])。伐除优势马尾松措施为采伐样地内平均胸径以上的马尾松, 采伐强度为70%(按胸高断面积计算);伐除非马尾松措施为采伐样地内胸径在4 cm以上的非马尾松(包括光皮桦、漆树等乔木, 以及部分木姜子、火棘、桦叶荚蒾等), 强度为15%;除灌措施为清除样地内林下灌木层, 但保留更新马尾松小树。采伐作业采用人力油锯伐木, 对采伐的树干进行移除[23]。
2.2 调查方法于2016年8月, 利用相邻格子调查方法, 将样地划分为25个10 m×10 m的样方, 对各样地内对所有胸径≥1 cm的乔木进行编号、挂牌并进行每木检尺, 记录样方号、树种、胸径。以每个样方为单元, 左下角为坐标原点测量所有胸径≥1 cm的乔木树种相对于坐标原点的平面直角坐标系的相对坐标(以X, Y的形式表示)。同时, 测定各样地的经纬度、海拔、坡度、坡向等立地因子。样地基本情况具体见表 1。
| 抚育择伐类型 Selective cutting types | 纬度 Latitude/N | 经度 Longitude/E | 海拔 Altitude/m | 坡度 Slope/° | 坡向 Aspect | 郁闭度 Canopy density | ≥5 cm树种 平均胸径 ≥5 cm mean DBH/cm | ≥5 cm树种 平均树高 ≥5 cm mean tree height/m |
| 未择伐Non-cutting | 30°59′20″ | 110°47′08″ | 1222.5 | 34 | 西北 | 0.7 | 12.10 | 9.23 |
| 伐除优势马尾松 Dominant Pinus massoniana-cutting | 30°59′23″ | 110°47′09″ | 1169 | 33 | 西北 | 0.3 | 9.45 | 8.85 |
| 伐除非马尾松 No Pinus massoniana-cutting | 30°59′21″ | 110°47′05″ | 1177.9 | 33 | 西北 | 0.6 | 15.18 | 11.36 |
| 除灌Shrubs-cutting | 30°59′23″ | 110°47′07″ | 1211 | 35 | 西北 | 0.7 | 14.35 | 10.41 |

|

|
Margalef丰富度指数(dM)
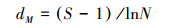
|
Shannon-Wiener多样性指数(H)
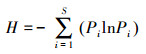
|
Simpson优势度指数(λ)
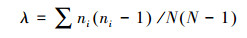
|
Pielou均匀度指数(Jsh)
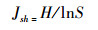
|
式中, S为物种数;N为所有物种的个体数之和;Pi=ni/N代表第i个物种的相对多度, ni为第i个物种的个体数。
2.3.3 乔木个体空间点格局分析首先, 将每个样方实测坐标进行全局化处理后得到林木个体坐标。然后, 采用Ripley′s K函数的改进型Ripley′s L函数[3, 10]分析不同择伐抚育措施主要树种的空间分布格局与种间空间关联性。具体公式如下:
Ripley′s K函数:
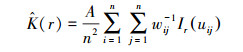
|
式中, A为研究区面积;uij为点i与j之间的距离;Ir(uij)为指示函数, 当uij≤r时, Ir(uij)=1, 当uij>r时, Ir(uij)=0;wij为权重值, 用于边缘校正。
Ripley′s L函数:
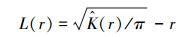
|
式中, L(r)=0时, 表示随机分布;L(r)>0时, 表示聚集分布;L(r) < 0时, 表示均匀分布。
利用Monte-Carlo函数来拟合检验计算上下包迹线即置信区间, 并以空间尺度r为横坐标, L(r)实际值和拟合值的上下包迹线为纵坐标作图。当分析单个种群时, 若种群实测值在包迹线以内, 则符合随机分布;若在包迹线上, 则为集群分布;若在包迹线下, 则为均匀分布。当分析两个种群间关联性时, 种群实际分布数据L12(r)高于上包迹线, 表示显著正相关, 在上下包迹线之间, 表示相关性不显著, 低于下包迹线则为显著负相关。
本研究随机模拟1000次, 达到99%的置信水平, 最大距离尺度为12.5 m, 为样地边长的1/4[14]。数据分析与作图利用R语言软件R-3.2.4中相关程序包以及Excel 13.0完成。
3 结果与分析 3.1 抚育择伐对马尾松林物种组成特征及物种多样性的影响抚育择伐3 a后, NC样地中乔木物种22种, DPMC 17种, NPMC 21种, SC17种。进一步统计出各措施下重要值≥5的乔木树种优势程度(表 2), NC样地中优势树种包括马尾松、茅栗、光皮桦、漆树、宜昌木姜子、盐肤木, 累积重要值占总体的82.34%;DPMC样地优势树种包括马尾松、香椿、光皮桦、漆树、盐肤木、宜昌木姜子、檫木、火棘, 累积重要值占总体的90.14%;NPMC样地优势树种包括马尾松、漆树、宜昌木姜子、盐肤木、香椿、火棘、檫木, 累积重要值占总体的87.23%;SC样地优势树种包括马尾松、漆树、光皮桦、香椿, 累积重要值占总体的73.3%。不同抚育择伐措施均在一定程度上降低了乔木层物种多样性(表 3), 与NC相比, DPMC、NPMC和SC的Margalef丰富度指数和Shannon-Wiener多样性指数分别降低28.43%、6.71%、12.79%和5.22%、2.17%、7.39%, Simpson优势度指数具有轻微增加, Pielou均匀度指数则无显著变化。不同抚育择伐措施均在一定程度上改变了物种组成以及部分树种的优势程度, 以DPMC和SC对林分物种组成影响最大。
| 物种名称 Species name | 未择伐 Non-cutting | 伐除优势马尾松 Dominant Pinus massoniana-cutting | 伐除非马尾松 No Pinus massoniana-cutting | 除灌 Shrubs-cutting |
| 马尾松Pinus massoniana | 30.97 | 17.9 | 41.03 | 38.52 |
| 茅栗Castanea seguinii | 13.84 | 3.24 | 3.73 | 4.85 |
| 光皮桦Betula luminifera | 11.82 | 12.38 | 2.66 | 12.39 |
| 漆树Toxicodendron vernicifluum | 11.62 | 12.37 | 10.94 | 13.01 |
| 宜昌木姜子Litsea ichangensis | 8.70 | 9.14 | 9.42 | 3.24 |
| 盐肤木Rhus chinensis | 5.39 | 10.12 | 7.77 | 1.79 |
| 火棘Pyracantha fortuneana | 3.59 | 5.77 | 5.77 | 0.37 |
| 化香Platycarya strobilacea | 3.57 | 0.23 | 2.93 | |
| 檫木Sassafras tzumu | 2.3 | 6.69 | 5.36 | 2.22 |
| 香椿Toona sinensis | 1.35 | 15.77 | 6.94 | 9.38 |
| 野樱桃Cornus chinensis | 1.35 | 0.91 | 0.67 | 4.93 |
| 君迁子Diospyros lotus | 1.22 | 0.87 | 0.37 | |
| 山胡椒Lindera glauca | 1.16 | 1.09 | 0.22 | |
| 青榨槭Acer davidii | 0.64 | 0.37 | ||
| 灯台树Bothrocaryum controversum | 0.50 | |||
| 白檀Symplocos paniculata | 0.38 | |||
| 山合欢Albizia kalkora | 0.33 | 0.22 | 1.09 | |
| 女贞Ligustrum lucidum | 0.30 | 1.43 | ||
| 香叶树Lindera communis | 0.28 | 0.44 | ||
| 短柄枹栎Quercus glandulifera | 0.24 | 0.22 | ||
| 朴树Celtis sinensis | 0.24 | |||
| 桤木Alnus cremastogyne | 0.24 | |||
| 合欢Albizia julibrissin | 0.37 | |||
| 槲栎Quercus aliena | 0.75 | |||
| 黄连木Pistacia chinensis | 0.44 | |||
| 杉木Cunninghamia lanceolata | 2.24 | 2.75 | ||
| 桃Amygdalus persica | 0.55 | 0.22 | ||
| 吴茱萸Evodia rutaecarpa | 1.03 | |||
| 腺柳Salix chaenomeloides | 1.47 | |||
| 香樟Cinnamomum camphora | 0.21 | |||
| 锥栗Castanea henryi | 0.47 | 0.22 |
| 抚育择伐类型 Selective cutting types | Margalef 丰富度指数 Margalef richness index(dM) | Shannon-Wiener 多样性指数 Shannon-Wiene species diversity index(H) | Simpson优势度指数 Simpson dominance index(λ) | Pielou均匀度指数 Pielou evenness index(Jsh) |
| 未择伐Non-cutting | 3.13 | 2.30 | 0.13 | 0.74 |
| 伐除优势马尾松 Dominant Pinus massoniana-cutting | 2.24 | 2.18 | 0.14 | 0.77 |
| 伐除非马尾松 No Pinus massoniana-cutting | 2.93 | 2.25 | 0.12 | 0.74 |
| 除灌Shrubs-cutting | 2.73 | 2.13 | 0.17 | 0.75 |
各样地内种群分布点图可以直观反映出各树种个体的空间分布状况。不同抚育择伐措施明显改变了各优势树种的空间格局, 且择伐强度和择伐对象选择的不同对优势树种空间分布格局的影响程度也存在差异(图 1)。

|
| 图 1 不同抚育择伐样地中主要乔木物种的空间分布散点图 Fig. 1 Spatial distribution pattern of major tree species in different selective cutting after three years NC:未择伐, Non-cutting;DPMC:伐除优势马尾松, Dominant Pinus massoniana-cutting;NPMC:伐除非马尾松, No Pinus massoniana-cutting;SC:除灌, Shrubs-cutting |
由图 2和图 3可知, 在NC样地中, 马尾松在5.5—6 m尺度上呈聚集分布, 其他尺度表现为随机分布;茅栗在0—12.5 m尺度上呈明显的聚集分布;漆树在0—7 m尺度上呈随机分布, 7—12.5 m尺度上表现为聚集分布;光皮桦在0—1.5 m尺度上呈聚集分布, >1.5 m的尺度上表现为随机分布;宜昌木姜子仅在0—0.5 m尺度上呈聚集分布, >0.5 m的尺度上表现为随机分布。在DMPC样地中, 马尾松在0—4.5 m尺度上呈随机分布, 4.5—12.5 m尺度上为聚集分布;漆树在0—0.5 m尺度和9—12.5 m尺度上呈随机分布, 而在0.5—9 m尺度上则表现为聚集分布;光皮桦、香椿、盐肤木在0—12.5 m尺度上均表现为聚集分布;宜昌木姜子和檫木在>0.5 m的尺度上表现为聚集分布;火棘在0—1.5 m尺度上呈聚集分布, 而>1.5 m尺度上表现为随机分布。在NPMC样地中, 马尾松在0—8 m尺度上呈聚集分布, 8—12.5 m尺度上表现为随机分布;漆树在1—12.5 m尺度上呈为聚集分布;宜昌木姜子在0—9.5 m和11.5 m尺度上呈随机分布, 9.5—11.5 m尺度上表现为聚集分布;香椿在0—8 m尺度和10.5—11.5 m尺度上呈聚集分布;盐肤木在0—12.5 m尺度上呈聚集分布;火棘在0—0.5 m和10.5—12.5 m尺度上呈聚集分布。在SC样地中, 马尾松在0—2 m尺度上呈随机分布, 2—8 m和11—12.5 m尺度上表现为轻微聚集分布;漆树和光皮桦在0—12.5 m上均表现为随机分布。

|
| 图 2 不同抚育择伐3 a后马尾松林4个优势树种的点格局分析 Fig. 2 Point pattern analysis of four dominant tree species in different selective cutting after three years NC:未择伐, Non-cutting;DPMC:伐除优势马尾松, Dominant Pinus massoniana-cutting;NPMC:伐除非马尾松, No Pinus massoniana-cutting;SC:除灌, Shrubs-cutting |

|
| 图 3 不同抚育择伐3 a后其他优势树种点格局分析 Fig. 3 Point pattern analysis of other dominant tree species in different selective cutting after three years |
比较不同抚育择伐措施中主要树种空间分布格局发现, NC样地中马尾松、漆树、光皮桦、宜昌木姜子等主要树种空间格局以随机分布为主。DMPC促使马尾松、光皮桦、香椿、盐肤木和火棘呈现出明显的聚集分布。NPMC导致光皮桦数量和地位严重下降, 且香椿和盐肤木主要以聚集分布格局为主, 其他树种空间分布格局与未择伐样地相似。SC进一步增加了林下更新阔叶树种呈明显的随机分布格局。
3.3 抚育择伐对马尾松林主要树种空间关联性的影响种间关联性是用来表示一定阶段内植物种群组成个体之间的空间相互关系和功能关系[25], 优势种间的关联性可以反映其空间依赖性[18], 分析研究人为抚育择伐干扰后的主要树种空间关联性, 有助于了解保留木在伐后群落中的作用。由图 4和图 5可以看出, 在NC样地中, 马尾松与漆树、马尾松与茅栗、光皮桦与漆树在0—12.5 m尺度内无明显空间相关性, 马尾松与光皮桦在5—11 m尺度上呈显著正相关, 宜昌木姜子与马尾松、漆树均在1—11 m尺度上呈显著正相关, 光皮桦与宜昌木姜子在4.5—11 m尺度上呈显著正相关。在DPMC样地中, 马尾松与漆树在8—12.5 m尺度上呈显著负相关, 与光皮桦在2—9 m尺度上呈显著负相关, 与盐肤木在2—8 m尺度上呈显著负相关;与香椿在10—12 m尺度上呈显著正相关;宜昌木姜子与马尾松、漆树在整个分析尺度上呈显著正相关;光皮桦与漆树在0—2.5 m尺度接近正相关, 而在4—12.5 m尺度接近或达到空间负相关, 与宜昌木姜子在1—9 m尺度上呈显著负相关。在NPMC样地中, 马尾松与漆树在1—12.5 m尺度上接近空间正相关, 与宜昌木姜子在0—5 m尺度上呈显著正相关, 与香椿在3—5.5 m尺度上呈显著负相关;马尾松与光皮桦、光皮桦与漆树、宜昌木姜子均在0—12.5 m尺度内无明显空间相关性, 与盐肤木在5—12.5 m尺度上呈显著负相关;漆树和宜昌木姜子在0—3.5 m尺度上呈显著正相关。在SC样地中, 仅马尾松与光皮桦在4—5.5 m和8—12.5 m尺度上呈接近或达到空间负相关, 而其他主要树种关联性呈明显的不相关。

|
| 图 4 不同抚育择伐3 a后马尾松与其他3个优势树种间空间关联性 Fig. 4 Interspecific spatial association between Pinus massoniana and other three dominant tree species in different selective cutting after three years |

|
| 图 5 不同抚育择伐3 a后其他优势树种间空间关联性 Fig. 5 Interspecific spatial association of other dominant tree species in different selective cutting after three years |
种群分布格局受多种因素影响, 如物种生物学特性、生境异质性、人为干扰因素等[1, 25-26]。抚育择伐往往会形成光照强弱、林隙效应以及土壤营养条件不均匀等空间异质性, 择伐对象不同势必会影响保留木的空间分布格局[11]。在本研究中, 未择伐马尾松林分的主要树种空间格局以随机分布为主, 优势树种间主要以不相关和正相关为主, 说明该阶段的林分树木已逐渐占据了有效的空间生态位, 种间相互竞争和排斥作用较弱, 林木空间分布格局已具备一般天然林分个体随机分布的特征[27]。然而, 伐除优势马尾松反而明显促使优势树种呈聚集分布, 且树种间的负相关性明显增加, 这是由于伐除优势马尾松影响了建群种和上林层结构, 林冠大开导致林下更新树种为了争夺更多的光照、水分、土壤、养分等生存条件, 加剧了树种间的竞争性。伐除非马尾松措施主要伐除个体较大的更新阔叶树种, 造成林内环境局部异质性增强[19], 导致漆树、香椿、盐肤木等目标择伐树种呈明显的聚集分布, 保留主要树种间以不相关为主, 在一定程度上降低了种间正相关性而呈现不相关, 降低了林分的稳定性。除灌措施主要去除林下植被层的灌木物种, 减少了林下层植被与更新树种的生存环境竞争, 促使更新阔叶树种随机分布程度增强, 致使更新树种间的生长空间扩大, 有利于林下树种的生长发育[28-29]。另外, 已有研究发现, 处于演替初期的群落中物种相关性主要以负相关和不相关为主, 随着植被演替的进行, 物种间负相关性逐渐减弱, 正相关性逐渐增强, 群落结构及物种组成也更加趋于稳定[2, 7]。可见, 抚育择伐导致林分发生逆向演替且对林分结构产生的影响各不相同, 可能主要由于不同物种或不同层次结构的物种在群落中的生物学特性、地位、作用、相互关系以及对外界干扰响应能力存在差异所产生的。
4.2 抚育择伐强度对马尾松林群落结构、物种空间关联性的短期影响择伐强度的大小严重制约了伐后林分的生长状况[15], 适度择伐有利于优化林分空间结构, 增强林分功能和稳定性[12, 30]。本研究中伐除优势马尾松(择伐强度为70%)属于强度择伐, 对林分扰动过大造成林内树种的聚集程度明显增加, 可能是由于强度择伐降低了上林层覆盖, 造成了大面积的林窗, 林窗内充足的资源促进了各个树种保留木和幼树的生长, 但同时树种竞争加剧又会制约优势树种的生长[28]。伐除非马尾松(择伐强度15%)属于中度择伐, 导致部分树种在大尺度上呈现聚集分布, 种间关联性以不相关为主, 可能是缘于该措施产生了有限的林窗, 促使某些树种呈聚集化有利于发挥功能群体效应, 利用有限的空间资源[10], 形成适合于自身生长的微环境, 从而抵御外部竞争[5], 而且伐后林分结构可在短期内恢复, 导致许多优势树种分布格局区域随机或轻微聚集分布。虽然本研究未统计除灌措施的择伐强度, 但由于林下层植被在森林总生物量中只占很少的一部分, 属轻度择伐。除灌进一步增强了各优势树种空间格局的随机分布以及树种空间关联性的不相关性。短期去除林下植被层暂时有利于加速林下更新速度, 但长期来看, 去除林下植被对上层乔木树种的生长不存在影响[31], 低强度择伐不能改变乔木树种对于绝大部分资源的竞争优势, 对林分结构优化改变的程度有限。同时, 森林群落组成和结构对抚育择伐活动的响应也存在一定的时间延迟[13-14], 短期择伐并未加速林分组成朝着演替后期的方向发展[11], 未来随着演替时间的发生, 抚育择伐后林分的群落组成结构将会进一步产生变化[7, 32]。
综上所述, 伐除优势马尾松、伐除非马尾松和除灌等抚育择伐措施均不同程度的改变了林木组成、空间关系、竞争状态。单纯调整某个树种或单个层次的结构来开展森林经营, 不利于林分空间结构的整体优化。因此, 未来针对三峡库区马尾松林经营管理时, 首先要基于林分空间格局和种间关联性判断林分是否需要人为经营手段的干预, 尽量减少人为干扰的程度和次数。其次, 择伐对象要优先选择清理衰老树、枯死木、生长势较弱的马尾松个体以及更新树种聚集分布区域和灌木层, 尽量保留有利于提高林分混交度的树种。再次, 遵循种群结构和空间格局分布规律, 保证林分地表的连续覆盖, 择伐强度应控制在15%左右, 防止林分结构剧烈变化, 通过产生适度的林窗, 改善林分生长环境, 促进马尾松林向针阔混异龄林发展, 进而实现森林结构优化和功能提升。
| [1] |
Hao Z Q, Zhang J, Song B, Ye J, Li B H. Vertical structure and spatial associations of dominant tree species in an old-growth temperate forest. Forest Ecology and Management, 2007, 252(1/3): 1-11. |
| [2] |
农友, 郑路, 贾宏炎, 卢立华, 黄德卫, 黄柏华, 雷丽群. 广西大青山次生林的群落特征及主要乔木种群的空间分布格局. 生物多样性, 2015, 23(3): 321-331. |
| [3] |
王鹏, 陈丽华, 卞西陈, 武巧英. 北沟林场天然次生林群落结构与种群分布格局. 应用生态学报, 2011, 22(7): 1668-1674. |
| [4] |
Masaki T, Ota T, Sugita H, Oohara H, Otani T, Nagaike T, Nakamura S. Structure and dynamics of tree populations within unsuccessful conifer plantations near the Shirakami Mountains, a snowy region of Japan. Forest Ecology and Management, 2014, 194(1/3): 389-401. |
| [5] |
龙成, 杨小波, 龙文兴, 李东海. 铜鼓岭热带常绿季雨矮林5种蒲桃属植物的种群结构及空间格局. 林业科学, 2015, 51(2): 18-27. |
| [6] |
Zunzunegui M, Esquivias M P, Oppo F, Gallego-Fernández J B. Interspecific competition and livestock disturbance control the spatial patterns of two coastal dune shrubs. Plant and Soil, 2012, 354(1/2): 299-309. |
| [7] |
韦博良, 袁志良, 牛帅, 刘霞, 贾宏汝, 叶永忠. 河南省宝天曼锐齿槲栎林树木死亡对空间格局及种间相关性的影响. 植物生态学报, 2017, 41(4): 430-438. |
| [8] |
Wang X G, Ye J, Li B H, Zhang J, Lin F, Hao Z Q. Spatial distributions of species in an old-growth temperate forest, northeastern China. Canadian Journal of Forest Research, 2010, 40(6): 1011-1019. DOI:10.1139/X10-056 |
| [9] |
郭屹立, 王斌, 向悟生, 丁涛, 陆树华, 黄俞淞, 黄甫昭, 李东兴, 李先琨. 广西弄岗北热带喀斯特季节性雨林监测样地种群空间点格局分析. 生物多样性, 2015, 23(2): 183-191. |
| [10] |
黄小波, 李帅峰, 苏建荣, 苏磊. 择伐对思茅松自然种群结构和空间分布格局的影响. 生态学报, 2015, 35(24): 8241-8250. |
| [11] |
齐麟, 赵福强. 不同采伐强度对阔叶红松林主要树种空间分布格局和物种空间关联性的影响. 生态学报, 2015, 35(1): 46-55. DOI:10.3969/j.issn.1673-1182.2015.01.010 |
| [12] |
Valverde-Barrantes O J, Rocha O J. Logging impacts on forest structure and seedling dynamics in a Prioria copaifera (Fabaceae) dominated tropical rain forest (Talamanca, Costa Rica). Revista De Biologia Tropical, 2014, 62(1): 347-357. |
| [13] |
Thorpe H C, Thomas S C, Caspersen J P. Residual-tree growth responses to partial stand harvest in the black spruce (Picea mariana) boreal forest. Canadian Journal of Forest Research, 2007, 37(9): 1563-1571. DOI:10.1139/X07-148 |
| [14] |
范春雨, 张春雨, 赵秀海. 择伐对吉林蛟河阔叶红松林群落结构及动态的影响. 生态学报, 2017, 37(20): 6668-6678. |
| [15] |
陈辉荣, 周新年, 蔡瑞添, 胡喜生, 巫志龙, 周成军. 天然林不同强度择伐后林分空间结构变化动态. 植物科学学报, 2012, 30(3): 230-237. |
| [16] |
吴建强, 王懿祥, 杨一, 朱婷婷, 朱旭丹. 干扰树间伐对杉木人工林林分生长和林分结构的影响. 应用生态学报, 2015, 26(2): 340-348. |
| [17] |
Ruel J C, Fortin D, Pothier D. Partial cutting in old-growth boreal stands:an integrated experiment. The Forestry Chronicle, 2013, 89(3): 360-369. DOI:10.5558/tfc2013-066 |
| [18] |
汤孟平, 周国模, 施拥军, 陈永刚, 吴亚琪, 赵明水. 天目山常绿阔叶林优势种群及其空间分布格局. 植物生态学报, 2006, 30(5): 743-752. DOI:10.3321/j.issn:1005-264X.2006.05.004 |
| [19] |
周建云, 李荣, 张文辉, 何景峰. 不同间伐强度下辽东栎种群结构特征与空间分布格局. 林业科学, 2012, 48(4): 149-155. |
| [20] |
陈幸良, 巨茜, 林昆仑. 中国人工林发展现状、问题与对策. 世界林业研究, 2014, 27(6): 54-59. |
| [21] |
葛晓改, 周本智, 肖文发. 马尾松人工林凋落物产量、养分含量及养分归还量特性. 长江流域资源与环境, 2014, 23(7): 996-1003. |
| [22] |
雷静品, 肖文发, 黄志霖, 曾立雄, 王怀清, 李良俊. 三峡库区上游马尾松生长及其与气候变化的关系. 华中农业大学学报, 2011, 30(4): 426-431. |
| [23] |
雷蕾, 肖文发, 曾立雄, 黄志霖, 谭本旺. 马尾松林土壤呼吸组分对不同营林措施的响应. 生态学报, 2016, 36(17): 5360-5370. |
| [24] |
李丽, 惠淑荣, 惠刚盈, 徐海, 胡艳波, 林天喜. 森林结构调查最小面积的研究. 林业资源管理, 2007(2): 47-51. DOI:10.3969/j.issn.1002-6622.2007.02.011 |
| [25] |
张兴旺, 张小平, 郭传友, 张强. 皖北石灰岩山地青檀种群不同发育阶段的点格局分析. 生态学杂志, 2013, 32(3): 542-550. |
| [26] |
张健, 郝占庆, 宋波, 叶吉, 李步杭, 姚晓琳. 长白山阔叶红松林中红松与紫椴的空间分布格局及其关联性. 应用生态学报, 2007, 18(8): 1681-1687. |
| [27] |
郑丽凤, 周新年. 择伐强度对中亚热带天然针阔混交林林分空间结构的影响. 武汉植物学研究, 2009, 27(5): 515-521. DOI:10.3969/j.issn.2095-0837.2009.05.011 |
| [28] |
杜忠, 蔡小虎, 包维楷, 陈槐, 潘红丽. 林下层植被对上层乔木的影响研究综述. 应用生态学报, 2016, 27(3): 963-972. |
| [29] |
Ares A, Neill A R, Puettmann K J. Understory abundance, species diversity and functional attribute response to thinning in coniferous stands. Forest Ecology and Management, 2010, 260(7): 1104-1113. DOI:10.1016/j.foreco.2010.06.023 |
| [30] |
Archambault L, Delisle C, Larocque G R. Forest regeneration 50 years following partial cutting in mixedwood ecosystems of southern Quebec, Canada. Forest Ecology and Management, 2009, 257(2): 703-711. DOI:10.1016/j.foreco.2008.09.056 |
| [31] |
Zhang J W, Oliver W W, Busse M D. Growth and development of ponderosa pine on sites of contrasting productivities:relative importance of stand density and shrub competition effects. Canadian Journal of Forest Research, 2006, 36(10): 2426-2438. DOI:10.1139/x06-078 |
| [32] |
缪宁, 刘世荣, 史作民, 马姜明, 王晖. 强度干扰后退化森林生态系统中保留木的生态效应研究综述. 生态学报, 2013, 33(13): 3889-3897. |