 2018, Vol. 38
2018, Vol. 38文章信息
- 贺燕燕, 王朝英, 袁中勋, 李晓雪, 杨文航, 宋虹, 李昌晓.
- HE Yanyan, WANG Chaoying, YUAN Zhongxun, LI Xiaoxue, YANG Wenhang, SONG Hong, LI Changxiao.
- 三峡库区消落带不同水淹强度下池杉与落羽杉的光合生理特性
- Photosynthetic characteristics of Taxodium ascendens and Taxodium distichum under different submergence in the hydro-fluctuation belt of the Three Gorges Reservoir
- 生态学报. 2018, 38(8): 2722-2731
- Acta Ecologica Sinica. 2018, 38(8): 2722-2731
- http://dx.doi.org/10.5846/stxb201704150673
-
文章历史
- 收稿日期: 2017-04-15
- 网络出版日期: 2018-01-08





三峡工程建成运行后, “冬蓄夏排”的水库调水方式给库区生态系统带来了巨大的负面影响[1], 消落带植被物种多样性降低[2], 水土流失严重[3], 进而导致消落带生态系统结构和功能受损。近年来, 消落带植被恢复受到社会各界的重视, 有研究表明人工植被重建是恢复消落带植被的有效方法之一[3-4]。消落带植物不仅要耐水淹, 且在退水后, 植物能否快速恢复其光合作用以制造和储存充分的碳水化合物将直接影响到其再次耐受水淹逆境后的能量供应及其在消落带的生存[5]。因此, 研究库区消落带适生植物在水淹后落干期的光合生理生态适应机制是消落带植被恢复的重要基础, 也是解决水库消落带生态环境问题的前提[6]。
水淹条件下, 缺氧使植物的生长、光合和代谢等过程受限。其对水淹敏感性植物的影响较大[7-8], 而耐淹植物可通过形成不定根、通气组织或茎伸长等方式适应水淹逆境[9-11]。作为植物生命活动最重要的过程之一, 光合响应变化已成为反映植物耐受胁迫能力的有效指标[12]。而光响应曲线的测定及其拟合有助于理解光合产物积累与环境因子的关系, 是植物光合生理生态学研究的重要手段之一[13-15]。通过光合光响应曲线可以获得光合作用的最大净光合速率(Pnmax)、表观量子效率(α)、光饱和点(LSP)、光补偿点(LCP)和暗呼吸速率(Rd)等生理参数, 而这些参数是植物生理生态学研究的基础[16-17], 有助于判定植物光合机构运转状况、光合作用能力和其受环境变化的影响程度[13, 18]。光响应曲线拟合模型则是光合作用对环境因子响应模拟系统的核心, 构建理想的光响应模型可更客观、准确地反映光响应过程[19]。国内外普遍采用的光响应曲线模型为:直角双曲线模型、非直角双曲线模型、直角双曲线修正模型和指数模型[13, 20-24]。由于不同植物对光照强度变化的敏感度及响应方式不同, 因而不同植物的最适光合光响应曲线的模型也不同。因此为了解植物的光合特性, 保证估测光合特性参数结果的准确性, 需首先对不同的光响应模型进行比较[25]。
池杉(T. ascendens)和落羽杉(T. distichum)为杉科落羽杉属落叶乔木。研究表明, 两树种具有较强的水淹耐受性[26], 是三峡库区消落带的适生乔木树种。目前关于两树种的研究多集中在模拟不同水淹环境、水淹胁迫解除后的生长、光合生理响应[27-28]、生理变化[29]和根系次生代谢[30]等, 尽管已有对三峡库区消落带原位落干期光合生理的研究[27], 但其研究仅反映了光合生理的瞬时响应, 而不能反映其对不同光强的利用情况, 缺乏应用光响应模型对两树种生长适应性的拟合研究。本研究以三峡库区消落带原位种植的池杉和落羽杉为研究对象, 测定其水淹后落干期的光响应曲线, 并采用直角双曲线模型、非直角双曲线模型、直角双曲线修正模型和指数模型4种模型对其进行拟合, 比较各模型拟合结果的差异并选出最优模型, 研究水淹后落干期两树种的光合生理变化, 探究其在消落带特殊生境下的适应机制, 以期为指导三峡库区消落带人工植被构建和管理提供理论依据。
1 材料与方法 1.1 试验地概况试验样地位于重庆市忠县共和村汝溪河消落带植被修复示范基地(107°32′—108°14′E, 30°03′—30°35′N), 面积0.133km2。汝溪河为长江一级支流, 流域属亚热带东南季风区山地气候。全年温度≥10℃, 年积温5787℃, 年均温18.2℃, 无霜期341d, 日照时数1327.5h, 日照率29%, 太阳总辐射能83.7×4.18kJ/cm2, 年降雨量1200mm, 相对湿度80%。
试验所选取的池杉和落羽杉树种位于消落带165—175m海拔范围内, 种植于2012年4月, 栽种时苗龄为2年生幼苗, 大小规格基本一致。种植时将样地分为20个宽10m、垂直于河床的条带, 每个条带栽植一个树种, 两个树种交替种植, 株行距为1m×1m。树苗栽好后立即浇水1次, 2012年6月中旬进行除草。此后无人为干扰自然生长。截至2016年7月试验开展时, 两树种已经历了4个水位波动周期, 并且长势良好。
1.2 试验设计于2016年7月中旬进行实地数据测定, 根据消落带不同海拔高程的淹水深度和淹水持续时间, 将样地划分为3个样带:浅淹(SS, 海拔175m, 对照)、中度水淹(MS, 海拔170m)和深度水淹(DS, 海拔165m), 各样带最大水淹深度及水淹天数见表 1。每个树种每个海拔高程分别选取4株长势相近的具有代表性植株, 每株选取3片完全舒展并且成熟、未受损的叶片(位于树冠的中上部)作为测定对象, 用于测定光响应曲线。
| 海拔 Elevation/m |
水淹深度 Submergence depth/m |
水淹天数 Submergence duration(d/a) |
|||
| 2012-06—2013-07 | 2013-06—2014-07 | 2014-06—2015-07 | 2015-06—2016-07 | ||
| 175 | 0 | 2 | 5 | 8 | 5 |
| 170 | 5 | 125 | 101 | 141 | 111 |
| 165 | 10 | 175 | 158 | 217 | 161 |
采用土壤氧化还原电位计DW-1型(江苏江分电分析仪器有限公司生产)测定样地土壤氧化还原电位(Eh)。将电位计探头插入距土壤表面10cm处, 持续几分钟待读数稳定后记录数值, 对每个海拔用于测定光响应曲线树种的生长土壤进行Eh的测定。通气良好及含氧量充分的土壤, 其Eh介于+400-(+700mV之间;淹水后Eh变化从+400到+72mV;而当Eh低于+350mV, 表明土壤氧气匮乏[31]。
1.4 光响应曲线的测定于2016年7月中旬选择晴朗天气进行原位测量, 测定时间在9:30—12:00。采用Li-6400便携式光合系统(Li-Cor 6400, Li-Cor Inc, USA)进行测定, 测定时用Li-6400-02B红蓝光源设定一系列光合有效辐射梯度:1800、1600、1400、1200、1000、800、600、400、200、100、50、0μmol m-2 s-1, 测定前用自然光对叶片进行充分诱导, 改变光照强度后, 最少稳定时间设定为120s。CO2浓度设置为(400±5)μmol/mol(仪器自带小钢瓶提供)。叶室温度和相对湿度均为自然背景值。实测值的计算参考郎莹等[20]的方法。
1.5 数据处理用Excel 2007和SPSS 22.0软件对光合测定仪Li-6400测得的数据进行处理。采用以下4种光合模型进行光响应曲线拟合, 得到各项光响应参数, 再分别对各项参数在模型间和处理组间做方差分析, 采用Duncan多重比较(Duncan′s multiple range test)进行显著性检验。同时, 将实测值和模型拟合值用Origin 9.0作图并比较。
1.5.1 直角双曲线模型[32]
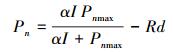
|
式中, Pn为净光合速率, I为光合有效辐射(结果与分析、讨论部分用PAR表示光合有效辐射), α为光响应曲线的初始斜率(即表观量子效率), Pnmax为最大净光合速率, Rd为暗呼吸速率。直线y =Pnmax与弱光下的线性方程相交, 交点对应x轴的值即为光饱和点LSP[33]。
1.5.2 非直角双曲线模型[34]
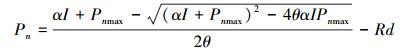
|
式中, θ为曲线的曲率, Pn、I、α、Pnmax和Rd的定义与前述相同。LSP的求算方法同直角双曲线模型。
1.5.3 直角双曲线修正模型[35]
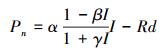
|
式中, β和γ为系数, Pn、I、α和Rd的定义与前述相同。拟合过程通过光合计算(版本4. 1. 1)完成, 由于拟合得到的LSP过大, 在表中未列出。
1.5.4 指数函数模型[20]
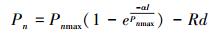
|
式中, Pn、I、α、Pnmax和Rd的定义与前述相同。其中LSP=Pnmax×ln(100C0)/α, 与90%Pnmax相对应的光照强度相比较求得[23]。C0为度量弱光下净光合速率趋于0的指标。
2 结果与分析 2.1 不同水淹处理组对池杉和落羽杉土壤氧化还原电位的影响3个水淹处理后两树种的Eh均有显著差异(表 2)。SS组的Eh始终高于420mV, 表明土壤通气良好。MS组的Eh在360.2—386.4mV范围波动, 而DS组的Eh低于350mV, 表明土壤氧气含量下降。
| 物种Species | 处理Treatment | ||
| SS | MS | DS | |
| 池杉T. ascendens | 420.60±5.51a | 386.40±5.09b | 346.40±3.80c |
| 落羽杉T.distichum | 425.60±3.54a | 360.20±2.46b | 331.60±11.42c |
| SS:浅淹, Shallow submergence;MS:中度水淹, Moderate submergence;DS:深度水淹, Deep submergence;不同小写字母表示同一物种在不同水淹处理组间差异显著(n=4, P < 0.05) | |||
经历4个水淹周期后的池杉和落羽杉长势良好。不同水淹处理组的两树种生长特征参数值变化趋势相似(表 3)。两树种的胸径与树高存在显著差异(P < 0.05), 均表现为SS组>MS组>DS组。随着水淹强度的增加, 两树种冠幅显著降低(P < 0.05)。
| 特征值Character | 池杉T. ascendens | 落羽杉T. distichum | |||||
| SS | MS | DS | SS | MS | DS | ||
| 胸径DBH/cm | 6.50±0.12a | 5.75±0.09b | 4.46±0.10c | 6.48±0.08a | 5.83±0.10b | 5.07±0.15c | |
| 冠幅Canopy/m2 | 5.38±0.24a | 2.86±0.32b | 2.45±0.09b | 8.87±0.78a | 6.62±0.23b | 5.52±0.27b | |
| 树高Tree height/m | 5.92±0.17a | 5.21±0.03b | 4.34±0.05c | 5.71±0.11a | 5.09±0.05b | 4.84±0.06c | |
| 不同小写字母表示同一物种在不同水淹处理组间差异显著(n=4, P < 0.05) | |||||||
不同水淹处理后的池杉和落羽杉采用不同模型拟合的变化趋势相似(图 1)。在弱光(PAR≤200μmol m-2 s-1)下, 4种模型拟合的光响应曲线变化规律均呈线性增长, 且各拟合值与实测值差值较小;随着光强的增加, 不同模型间的差异逐渐增大, 4种模型的净光合速率(Pn)均随着光强的增加而升高, 但逐渐趋于平缓。而不同模型间的变化幅度差异较大。

|
| 图 1 不同光响应模型对池杉和落羽杉光合速率光响应曲线的拟合 Fig. 1 Simulation of photosynthetic rate-light response curves of T. ascendens and T. distichum by 4 models under different submergence treatment SS:浅淹, Shallow submergence;MS:中度水淹, Moderate submergence;DS:深度水淹, Deep submergence;A:直角双曲线模型(池杉);A′:直角双曲线模型(落羽杉);B:非直角双曲线模型(池杉);B′:非直角双曲线模型(落羽杉);C:直角双曲线修正模型(池杉);C′:直角双曲线修正模型(落羽杉);D:指数模型(池杉);D′:指数模型(落羽杉) |
通过对不同模型的进一步分析(表 4)可知, 各模型的决定系数(R2)均在0.9以上, 表明4种模型均能较好地拟合池杉和落羽杉的光响应过程。在本研究不同水淹处理下, 两树种采用直角双曲线模型与非直角双曲线模型拟合的α、Pnmax、Rd与实测值相比显著偏大, 而LSP和LCP则显著低于实测值。修正模型的拟合精度较高, R2达到0.97以上, 可以准确地预测Pnmax;但修正参数β < 0且(β+γ)/β < 0[36], 求出LSP太大(表 4中未列出), 且α和Rd也均大于实测值, LCP低于实测值。指数模型拟合出的α和Rd最接近实测值, Pnmax和LSP虽无显著差异, 但相比其他模型更接近实测值。结合决定系数发现, R2的大小仅反映模型拟合程度, 并不能说明各模型拟合的参数值与实测值相符。上述研究结果表明, 指数模型为池杉和落羽杉光响应曲线拟合的最优模型, 故以此模型为基础分析两树种的光响应曲线特征。
| 树种 Species |
模型 Model |
处理 Treatment |
α/ (μmol/μmol) |
Pnmax /(μmol m-2 s-1) |
LSP/ (μmol m-2 s-1) |
LCP/ (μmol m-2 s-1) |
Rd/ (μmol m-2 s-1) |
R2 |
| 池杉 | 实测值 | SS | ≈ 0.031 | ≈ 8.55 | ≈1600.00 | ≈ 59.94 | ≈0.50 | — |
| T. ascendens | MS | ≈ 0.043 | ≈ 15.10 | ≈ 1640.00 | ≈ 42.30 | ≈0.60 | — | |
| DS | ≈ 0.044 | ≈ 18.14 | ≈ 1640.00 | ≈ 71.05 | ≈0.80 | — | ||
| Ⅰ | SS | 0.060±0.012Aa | 11.79±0.49 Ac | 484.46±74.15Bb | 53.87±8.95Aa | 2.33±0.41Aa | 0.968 | |
| MS | 0.073±0.006Aa | 18.73±1.99Ab | 548.54±66.31Aab | 38.50±12.59Aa | 2.19±0.53Aa | 0.985 | ||
| DS | 0.068±0.013Aa | 25.03±0.85Aa | 710.37±48.46Aa | 66.58±5.63Aa | 3.74±0.58Aa | 0.992 | ||
| Ⅱ | SS | 0.056±0.009Aa | 11.70±0.55Ac | 482.11±75.63Ba | 54.33±9.12Aa | 2.31±0.41Aa | 0.968 | |
| MS | 0.068±0.004Aa | 18.63±2.08Ab | 545.42±68.50Aab | 38.36±12.73Aa | 2.14±0.55Aa | 0.986 | ||
| DS | 0.068±0.013Aa | 25.03±0.85Aa | 710.56±48.48Aa | 66.55±5.65Aa | 3.74±0.58Aa | 0.992 | ||
| Ⅲ | SS | 0.060±0.013Aa | 9.51±0.85BCc | — | 53.70±9.16Aab | 2.31±0.46Aab | 0.968 | |
| MS | 0.062±0.007Aa | 16.02±2.08Ab | — | 37.77±12.75Ab | 1.92±0.55Ab | 0.983 | ||
| DS | 0.063±0.010Aa | 21.63±0.59Ba | — | 83.73±14.90Aa | 3.54±0.45Aa | 0.992 | ||
| Ⅳ | SS | 0.031±0.005Aa | 7.81±0.61Bc | 1185.17±187.3Ba | 51.26±9.26Aa | 1.27±0.09Aa | 0.952 | |
| MS | 0.043±0.004Ba | 13.60±1.22Ab | 1352.34±140.76Aa | 28.19±3.86Ab | 1.31±0.04Aa | 0.977 | ||
| DS | 0.039±0.005Aa | 16.74±0.47Ca | 1615.76±74.89Aa | 33.46±3.45Bab | 1.28±0.02Ba | 0.985 | ||
| 落羽杉 | 实测值 | SS | ≈0.023 | ≈5.98 | ≈1240.00 | ≈24.83 | ≈0.80 | — |
| T. distichum | MS | ≈0.026 | ≈7.24 | ≈1400.00 | ≈39.05 | ≈0.90 | — | |
| DS | ≈0.034 | ≈9.29 | ≈1400.00 | ≈41.61 | ≈1.30 | — | ||
| Ⅰ | SS | 0.103±0.033Aa | 10.00±0.00Ab | 488.48±47.12Aa | 29.43±7.43Aa | 2.46±0.72Aa | 0.916 | |
| MS | 0.055±0.010Aa | 10.41±0.25Ab | 444.27±19.94Aa | 36.30±3.73Aa | 1.72±0.35Aa | 0.981 | ||
| DS | 0.088±0.008Aa | 12.10±0.30Aa | 491.74±36.77Ca | 34.13±7.57Aa | 2.30±0.50Aa | 0.990 | ||
| Ⅱ | SS | 0.052±0.008ABa | 7.06±0.45Bc | 345.74±21.71Ab | 26.34±2.91Aa | 1.17±0.17Ba | 0.977 | |
| MS | 0.043±0.004ABa | 9.14±0.47Bb | 392.76±13.86Aab | 36.36±3.60Aa | 1.41±0.16Aa | 0.992 | ||
| DS | 0.062±0.008ABa | 11.23±0.42Aa | 458.70±30.06Ca | 37.78±8.44Aa | 2.02±0.49Aa | 0.992 | ||
| Ⅲ | SS | 0.060±0.006ABab | 5.86±0.35Cc | — | 25.09±2.17Aa | 1.25±0.17Ba | 0.975 | |
| MS | 0.049±0.005Ab | 7.42±0.19Cb | — | 34.45±2.94Aa | 1.47±0.20Aa | 0.991 | ||
| DS | 0.077±0.012Aa | 8.69±0.57Ba | — | 33.93±7.56Aa | 2.06±0.55Aa | 0.991 | ||
| Ⅳ | SS | 0.026±0.003Bb | 5.39±0.44Cc | 1120.11±58.31Aa | 39.33±3.22Aab | 1.20±0.04Ba | 0.962 | |
| MS | 0.030±0.004Bb | 6.90±0.32Cb | 1200.04±130.38Aa | 45.52±5.99Aa | 1.27±0.03Aa | 0.984 | ||
| DS | 0.047±0.004Ba | 8.45±0.57Ba | 1120.32±48.99Ba | 29.95±3.75Ab | 1.24±0.07Aa | 0.990 | ||
| Ⅰ:直角双曲线模型, Rectangular hyperbola model;Ⅱ:非直角双曲线模型, Non-rectangular hyperbola model;Ⅲ:直角双曲线修正模型, Modified model;Ⅳ:指数模型, Exponential equation model, α:表观量子效率, Apparent quantum yield;Pnmax:最大净光合速率, Maximum net photosynthetic rate;LSP:光饱和点, Light saturation point;LCP:光补偿点, Light compensation point;Rd:暗呼吸速率, Dark respiration rate;R2:决定系数, Determination coefficient; 数据为平均值±标准误, 采用Duncan法(n=4, P < 0.05)检验;不同小写字母表示同一树种相同模型在不同处理间显著差异, 大写字母表示同一树种相同处理在不同模型间显著差异 | ||||||||
在不同水淹处理组两树种的光响应曲线均有相似的变化规律(图 1), 即在同一光强下的Pn值均表现为DS组>MS组>SS组。但不同水淹处理组两树种光响应曲线的高低变化表现出较大差异。随着测定光强的增大, MS、DS组叶片的Pn持续增加, 而SS组叶片则在较低PAR时就达到了光饱和, 之后趋向平稳。在测定的光强范围内, 两树种并没有出现光抑制现象, 即在光强为1800μmol m-2 s-1时Pn仍有小幅度增长趋势, 依然没有达到光饱和(图 1), 且Pnmax和LSP均相对较高(表 4)。不同水淹处理组池杉的Pnmax、LSP和LCP均远大于落羽杉。
对光响应曲线拟合结果表明(表 4), 经历了4个水位周期性波动的两树种, 在不同水淹处理后, MS和DS组的各光合参数比SS组均有提高。对于池杉, 3个水淹处理组的α、LSP和Rd均无显著差异。而MS组的Pnmax比SS组显著提高了74.1%(P < 0.05), DS比MS组的Pnmax显著提高了23.1%(P < 0.05)。淹水后的LCP比SS组显著降低(P < 0.05), MS和DS组的LCP比SS组分别降低了35.4%和45.0%。对落羽杉而言, DS组的α比SS和MS组显著提高。MS和DS组的Pnmax比SS组分别提高了56.8%和28.0%, DS比MS组的Pnmax显著提高了23.0%(P < 0.05)。3个水淹处理组的LSP和Rd均无显著差异。MS和DS组的LCP分别比SS组降低了23.8%和9.7%。
3 讨论 3.1 光合光响应曲线拟合模型比较通过数学模型对光合光响应曲线的拟合, 可以快速求得植物叶片的特征参数, 对了解光合生理过程有重要意义[19]。但不同模型拟合得出的参数精确度存在一定差异, 本研究结果(图 1和表 4)也证实了该结论。另外, 遭受其他逆境胁迫下的山杏(Prunus sibirica L.)[22]、银杏(Ginkgo biloba)[23]和栾树(Koelreuteria paniculata Laxm.)[33]等植物多模型拟合的研究结果表明, 修正模型的拟合效果最好。本研究表明在LSP的拟合上修正模型存在不足, 这与邓云鹏等[37]通过多模型对不同种源栓皮栎(Quercus variabilis)的光响应曲线的拟合研究结果一致。因此, 模型的正确选择决定了光合特征参数的准确性, 才能真正反映植物的光合能力[38]。可见在研究植物的光响应曲线时, 应综合几个模型的结果进行分析, 提高对模型中各参数估计的准确性。综上所述, 指数模型的拟合结果更符合两树种的生理学意义。
3.2 水淹后落干期池杉和落羽杉的光合曲线的响应植物的光响应曲线最能反映植物对光照强度的适应特性, 同时也可以反映植物对环境的适应能力[39]。Pnmax是描述植物最大光合潜能的重要指标[40]。本研究表明, MS和DS组的池杉和落羽杉的Pnmax比SS组均有显著提高(表 4), 这可能与水淹结束后两树种的适应机制有关。有研究表明, 不同程度水淹后的中山杉(Taxodium hybrid′ zhongshansha′)的Pnmax比对照均有所提高, 表明其叶片光合潜力并未因淹水和淹水后落干期而降低, 相反, 水淹对其叶片的光合潜力的增强有一定的促进作用[28]。由此可以推断, 两树种表现出对水淹胁迫较强的适应能力, 这可能是因为中度水淹和深度水淹的两树种为适应下一次的水淹而通过增强光能利用效率去生产积累更多的光合产物。其中, 强光下池杉比落羽杉具有更高的Pnmax, 其光合同化潜力大, 表明落羽杉在水淹胁迫后的光合恢复能力弱于池杉。有研究表明, 在一定的环境条件下叶片的Pnmax决定于Rubisco活性与电子传递速率[41]。在本试验条件下, 水淹后引起两树种的Pnmax和α提高, 是否由于前期水淹胁迫导致PSⅡ电子传递速率及叶肉细胞Rubisco活性的提高, 还有待于进一步研究。
α值的高低, 反映了植物光合作用的光能利用效率, 尤其是对弱光的利用能力[40]。α越高, 说明其叶片光能转化效率越高。较多研究表明, 植物的αmax理论值在0.08—0.125μmol/μmol之间, 但在自然条件下的α值均低于理论值, 一般在0.03—0.05μmol/μmol之间[18]。水分胁迫会影响植物的α值[42], 但对于不同植物的α与土壤水分的定量关系还不清楚。池杉在3个水淹处理组的α并不具有显著性差异(表 4), 其范围在0.031—0.043μmol/μmol波动;而落羽杉的α范围在0.026—0.047μmol/μmol波动。与一般植物在适宜生长条件下的α基本一致。这说明两树种的叶片均具有较强的光能转化效率。在本研究中, 两树种在光饱和时的Pn和α均表现为DS组>MS组>SS组。有研究表明, LSP较高, LCP较低, 光饱和点时Pn和α较大, 表明植物在此条件下能最有效的利用光能, 表现为植物体的生长势以及光合生产能力强[43]。因此, 水淹后两树种较高的Pn和α有助于两树种充分利用光能, 积累更多光合产物, 这是其适宜生长在三峡库区消落带水淹-落干交替变化的特殊生境中的原因之一。
LSP和LCP是衡量植物需光特性的重要生理指标, 也反应植物对环境的适应能力[44]。LSP越大, 则植物利用强光的能力越强[45];LCP越小, 表明植物利用弱光的能力越强。LSP显著减小会影响植物对强光的利用, 影响植物的光合, 甚至会导致植物发生光抑制。夏季是三峡库区消落带内植物的生长期, LSP的高低将直接影响植物的存活和生长。本研究中, 两树种各水淹处理组间的LSP无显著差异, 而经历水淹后池杉的LCP均低于SS组, 表明池杉的LCP随水淹深度的增加有明显降低的趋势, 说明水淹不仅未对池杉对强光的利用产生影响, 还促进了池杉对弱光的利用能力。这是水淹后两树种能在消落带内持续生长的又一重要原因。
Rd值的高低, 与植物的生长状况有关[46], 而大部分植物的Rd均 < 2μmol m-2 s-1[5]。在3个处理组中, 两树种的Rd值均无显著差异, 但较浅淹组有所提高, 说明两树种在淹水后落干恢复期为维持正常生命活动需要更多的光合有机物, 这与裴顺祥等[6]对水淹后狗牙根(Hemarthria altissima)光响应曲线的研究结果一致。Rd的高低与植物生长速率关系密切, 生长快的植物呼吸速率会相应加快[47]。水淹后两树种的Rd升高, 说明其在落干期生长加快, 有利于其积累更多的有机物以抵御下一次水淹。
大量研究表明, 水淹致使土壤中气体交换速率明显下降, 植物的无氧呼吸逐渐替代了有氧呼吸, 植物的生长和形态发生明显变化[28-29, 48]。从不同水淹处理组池杉和落羽杉的胸径、冠幅和树高等3个生长参数(表 3)看, 均表现为SS组>MS组>DS组, 说明不同水淹强度对两树种的影响不同;水淹强度越大, 对两树种的生长影响也越大。植物在水淹后的恢复生长状况是衡量其耐受水淹胁迫能力的重要指标[49]。本研究中, 相比于SS组, 经历前期淹水的两树种在恢复期的光合速率非但没有降低, 反而均高于浅淹, 并呈水淹深度增大其増幅越大的变化趋势。这一现象与环境胁迫会限制生物体生长的理论不尽相同, 但用"中等胁迫理论"[50]来诠释, 也属于一种正常的生理现象。该理论认为, 生物体所受的胁迫若未超过其自身的耐性限度, 生物会在解除胁迫后, 迅速地启动恢复机制, 进而使各项生命活动正常化, 且足够强的胁迫还会诱导生物体产生最大程度的耐受能力, 这对其生理功能的进化有一定的促进作用, 反之, 不受胁迫的生物体则不具备这样的能力。
另外, DS组池杉和落羽杉的光合能力均高于MS组, 这可能与水淹后植物根系孔隙度的增加有关。王瑗等[51]对落羽杉(T. distichum)、薄壳山核桃(Carya illinoensis)和乌柏(Sapium sebiferum)耐涝渍的研究表明, 水淹会增加植物不定根和根系孔隙度的数量, 并随着水淹深度的増加, 二者显著增多。植物的不定根形成和根系孔隙度增加与植物耐淹性密切相关。也有研究表明长期生活在沼泽地带的水稻, 其根系具有很发达的细胞间隙和气道, 与茎、叶的细胞间隙相连, 可改善根系的通气状况, 较高的根系氧浓度提高了植物的根系活力, 促进植株对营养元素的吸收, 进而提髙水稻的光合作用[52]。在试验测定时, 本课题组也发现样地中两树种有明显膝状呼吸根的形成。这也可能是两树种水淹后光合能力增强的一个原因。表明两种植物通过形态改变来适应消落带恶劣环境。
4 结论三峡库区消落带人工重建植被体系中的池杉和落羽杉在经历了4a的水淹-落干循环后, 表现出良好的生长适应能力。两树种在水淹后落干期的光合能力提高, 可能与其遭受胁迫后的积极自我调节能力和光合补偿机制有关。在不同水淹处理组的两树种表现出了不同的光合特性。池杉表现出更高的光合活性, 其Pn、Pnmax、α、LSP、LCP及Rd值均比落羽杉高, 表明生长在同一生境中的池杉对光照、水分利用的恢复适应能力高于落羽杉。这与不同植物对水淹生理适应性的差异有关。
本研究虽为原位实验, 但仍存在一些不足, 主要包括以下两方面内容。首先, 本研究只对池杉和落羽杉落干期进行了一次测试工作, 而对其恢复期生长缺少全面监测;其次, 本研究仅对水淹后落干期两树种的光合生理特征方面做了初步研究。而植物对于三峡库区消落带原位生境的适应是一个非常复杂的过程, 因此需开展进一步的研究工作, 如糖代谢、信号传导机制以及基因表达等方面的研究, 从而全面揭示两树种在三峡库区消落带的水淹适应机制。这些也是本课题组目前正在进一步深入开展的工作。
| [1] | New T, Xie Z Q. Impacts of large dams on riparian vegetation:applying global experience to the case of China's Three Gorges Dam. Biodiversity & Conservation, 2008, 17(13): 3149–3163. |
| [2] | 刘维暐, 王杰, 王勇, 杨帆. 三峡水库消落区不同海拔高度的植物群落多样性差异. 生态学报, 2012, 32(17): 5454–5466. |
| [3] | 鲍玉海, 贺秀斌. 三峡水库消落带土壤侵蚀问题初步探讨. 水土保持研究, 2011, 18(6): 190–195. |
| [4] | 樊大勇, 熊高明, 张爱英, 刘曦, 谢宗强, 李兆佳. 三峡库区水位调度对消落带生态修复中物种筛选实践的影响. 植物生态学报, 2015, 39(4): 416–432. DOI:10.17521/cjpe.2015.0041 |
| [5] | 裴顺祥, 洪明, 郭泉水, 秦爱丽, 朱莉, 朱妮妮. 三峡库区消落带水淹结束后狗牙根的光合生理生态特性. 生态学杂志, 2014, 33(12): 3222–3229. |
| [6] | 揭胜麟, 樊大勇, 谢宗强, 张想英, 熊高明. 三峡水库消落带植物叶片光合与营养性状特征. 生态学报, 2012, 32(6): 1723–1733. |
| [7] | Mommer L, Pedersen O, Visser E J W. Acclimation of a terrestrial plant to submergence facilitates gas exchange under water. Plant, Cell & Environment, 2004, 27(10): 1281–1287. |
| [8] | Perata P, Armstrong W, Voesenek L A C J. Plants and flooding stress. New Phytologist, 2011, 190(2): 269–273. DOI:10.1111/j.1469-8137.2011.03702.x |
| [9] | Kozlowski T T, Pallardy S G. Acclimation and adaptive responses of woody plants to environmental stresses. The Botanical Review, 2002, 68(2): 270–334. DOI:10.1663/0006-8101(2002)068[0270:AAAROW]2.0.CO;2 |
| [10] | Colmer T D, Pedersen O. Underwater photosynthesis and respiration in leaves of submerged wetland plants:gas films improve CO2and O2 exchange. New Phytologist, 2008, 177(4): 918–926. DOI:10.1111/nph.2008.177.issue-4 |
| [11] | 王海锋, 曾波, 李娅, 乔普, 叶小齐, 罗芳丽. 长期完全水淹对4种三峡库区岸生植物存活及恢复生长的影响. 植物生态学报, 2008, 32(5): 977–984. |
| [12] | 刘泽彬. 三峡库区消落带两种植物对淹水环境适应性的模拟研究[D]. 北京: 中国林业科学研究院, 2014. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y2629960 |
| [13] | 叶子飘. 光合作用对光和CO2响应模型的研究进展. 植物生态学报, 2010, 34(6): 727–740. |
| [14] | Buckley T N, Diaz-Espejo A. Partitioning changes in photosynthetic rate into contributions from different variables. Plant, Cell & Environment, 2015, 38(6): 1200–1211. |
| [15] | 蒋冬月, 钱永强, 费英杰, 王正超, 孙振元. 柳属植物光合-光响应曲线模型拟合. 核农学报, 2015, 29(1): 169–177. DOI:10.11869/j.issn.100-8551.2015.01.0169 |
| [16] | 徐玲玲, 张宪洲, 石培礼, 于贵瑞. 青藏高原高寒草甸生态系统表观量子产额和表观最大光合速率的确定. 中国科学D辑地球科学, 2004, 34(S2): 125–130. |
| [17] | 张弥, 吴家兵, 关德新, 施婷婷, 陈鹏狮, 纪瑞鹏. 长白山阔叶红松林主要树种光合作用的光响应曲线. 应用生态学报, 2006, 17(9): 1575–1578. |
| [18] | 叶子飘. 光响应模型在超级杂交稻组合-Ⅱ优明86中的应用. 生态学杂志, 2007, 26(8): 1323–1326. |
| [19] | Sharp R E, Matthews M A, Boyer J S. Kok effect and the quantum yield of photosynthesis:light partially inhibits dark respiration. Plant Physiology, 1984, 75(1): 95–101. DOI:10.1104/pp.75.1.95 |
| [20] | Bassman J H, Zwier J C. Gas exchange characteristics of Populus trichocarpa, Populus deltoides and Populus trichocarpa×P. deltoides clones. Tree Physiology, 1991, 8(2): 145–159. DOI:10.1093/treephys/8.2.145 |
| [21] | Kyei-Boahen S, Lada R, Astatkie T, Gordon R, Caldwell C. Photosynthetic response of carrots to varying irradiances. Photosynthetica, 2003, 41(2): 301–305. DOI:10.1023/B:PHOT.0000011967.74465.cc |
| [22] | 郎莹, 张光灿, 张征坤, 刘顺生, 刘德虎, 胡小兰. 不同土壤水分下山杏光合作用光响应过程及其模拟. 生态学报, 2011, 31(16): 4499–4508. |
| [23] | 王欢利, 曹福亮, 刘新亮. 高温胁迫下不同叶色银杏嫁接苗光响应曲线的拟合. 南京林业大学学报:自然科学版, 2015, 39(2): 14–20. |
| [24] | 王小菲, 高文强, 刘建锋, 倪妍妍, 屈璐, 赵秀莲, 杨文娟, 邓云鹏, 江泽平. 不同生境对栓皮栎幼苗光合生理特性的影响. 生态学报, 2016, 36(24): 8062–8070. |
| [25] | 肖丹丹, 左力辉, 王进茂, 梁海永, 史亚朋, 杨润蕾, 杨敏生. 5种榆属植物光响应曲线模型对比分析. 中国农业科技导报, 2016, 18(4): 124–131. |
| [26] | 汪企明, 江泽平, 吕祥生, 张继凡, 鲁开基, 孙永召, 郭士祥, 吴孟军, 陆兴安. 落羽杉属种源研究:树种生物学特性. 江苏林业科技, 1995, 22(2): 14–18. |
| [27] | Wang C Y, Li C X, Wei H, Xie Y Z, Han W J. Effects of long-term periodic submergence on photosynthesis and growth of Taxodium distichum and Taxodium ascendens saplings in the hydro-fluctuation zone of the Three Gorges Reservoir of China. PloS One, 2016, 11(9): e0162867. DOI:10.1371/journal.pone.0162867 |
| [28] | 凌子然. 不同程度水淹对中山杉及亲本生长与光合生理恢复的影响[D]. 南京: 南京大学, 2016. http://cdmd.cnki.com.cn/Article/CDMD-10284-1016123461.htm |
| [29] | 李川, 周倩, 王大铭, 辜夕容. 模拟三峡库区淹水对植物生长及生理生化方面的影响. 西南大学学报:自然科学版, 2011, 33(10): 46–50. |
| [30] | 李昌晓, 钟章成, 陶建平. 不同水分条件下池杉幼苗根系的苹果酸、莽草酸含量及生物量. 林业科学, 2008, 44(10): 1–7. |
| [31] | Pezeshki S R. Root responses of flood-tolerant and flood-sensitive tree species to soil redox conditions. Trees, 1991, 5(3): 180–186. |
| [32] | Baly E C C. The kinetics of photosynthesis. Proceedings of the Royal Society B:Biological Sciences, 1935, 117(804): 218–239. DOI:10.1098/rspb.1935.0026 |
| [33] | 陈志成, 王荣荣, 王志伟, 杨吉华, 王华田, 耿兵, 张永涛. 不同土壤水分条件下栾树光合作用的光响应. 中国水土保持科学, 2012, 10(3): 105–110. |
| [34] | Thornley J H M. Mathematical Models in Plant Physiology. London: Academic Press, 1976. |
| [35] | Ye Z P, Yu Q. A coupled model of stomatal conductance and photosynthesis for winter wheat. Photosynthetica, 2008, 46(4): 637–640. DOI:10.1007/s11099-008-0110-0 |
| [36] | 梁文斌, 聂东伶, 吴思政, 柏文富, 沈素贞. 短梗大参光合作用光响应曲线及模型拟合. 经济林研究, 2014, 32(4): 38–44. |
| [37] | 邓云鹏, 雷静品, 潘磊, 王晓荣. 不同种源栓皮栎光响应曲线的模型拟合及参数比较. 生态学杂志, 2016, 35(2): 387–394. |
| [38] | 王秀伟, 毛子军. 7个光响应曲线模型对不同植物种的实用性. 植物研究, 2009, 29(1): 43–48. |
| [39] | 廖小锋, 刘济明, 张东凯, 靳勇, 张勇, 闫国华, 王敏. 野生小蓬竹的光合光响应曲线及其模型拟合. 中南林业科技大学学报, 2012, 32(3): 124–128. |
| [40] | 熊彩云, 曾伟, 肖复明, 曾志光, 涂圣勇, 江斌, 邱凤英, 伍艳芳. 木荷种源间光合作用参数分析. 生态学报, 2012, 32(11): 3628–3634. |
| [41] | Watling J R, Press M C, Quick W P. Elevated CO2 induces biochemical and ultrastructural changes in leaves of the C4 cereal sorghum. Plant Physiology, 2000, 123(3): 1143–1152. DOI:10.1104/pp.123.3.1143 |
| [42] | 陈建, 张光灿, 张淑勇, 王梦军. 辽东楤木光合和蒸腾作用对光照和土壤水分的响应过程. 应用生态学报, 2008, 19(6): 1185–1190. |
| [43] | 惠竹梅, 焦旭亮, 张振文. 渭北旱塬'赤霞珠'葡萄浆果膨大期光合特性研究. 西北农林科技大学学报:自然科学版, 2008, 36(4): 111-116, 122-122. |
| [44] | 陆銮眉, 林金水, 杜晓娜, 卞阿娜. 短叶金边虎尾兰的光合和叶绿素荧光特性研究. 漳州师范学院学报:自然科学版, 2010, 23(4): 108–112. |
| [45] | 伍维模, 李志军, 罗青红, 韩路. 土壤水分胁迫对胡杨、灰叶胡杨光合作用-光响应特性的影响. 林业科学, 2007, 43(5): 30–35. |
| [46] | 韩刚, 赵忠. 不同土壤水分下4种沙生灌木的光合光响应特性. 生态学报, 2010, 30(15): 4019–4026. |
| [47] | 潘瑞炽. 植物生理学(第四版). 北京: 高等教育出版社, 2001. |
| [48] | Simova-Stoilova L, Demirevska K, Kingston-Smith A, Feller U. Involvement of the leaf antioxidant system in the response to soil flooding in two Trifolium genotypes differing in their tolerance to waterlogging. Plant Science, 2012, 183(9): 43–49. |
| [49] | Mommer L, Lenssen J P M, Huber H, Visser E J W, de Kroon H. Ecophysiological determinants of plant performance under flooding:a comparative study of seven plant families. Journal of Ecology, 2006, 94(6): 1117–1129. DOI:10.1111/jec.2006.94.issue-6 |
| [50] | 常杰, 葛滢. 生态学. 杭州: 浙江大学出版社, 2001. |
| [51] | 王瑗. 涝渍对三个树种生长及生理生化的影响[D]. 南京: 南京林业大学, 2011. |
| [52] | 张达, 林杉, 黄永芬, 曹一平. 通气状况与氮素形态对水稻和旱稻生长的影响. 哈尔滨师范大学自然科学学报, 2002, 18(4): 97–102. |