 2018, Vol. 38
2018, Vol. 38文章信息
- 公绪云, 饶兴权, 周丽霞, 王晓玲, 朱小林, 蔡锡安.
- GONG Xuyun, RAO Xingquan, ZHOU Lixia, WANG Xiaoling, ZHU Xiaolin, CAI Xi'an.
- 尾叶桉林下5种植物的耐阴性、生物量及其个体消长
- Dynamics of shade tolerance, biomass, and individual growth of five understory plant species in Eucalyptus urophylla plantations
- 生态学报. 2018, 38(3): 1124-1133
- Acta Ecologica Sinica. 2018, 38(3): 1124-1133
- http://dx.doi.org/10.5846/stxb201612022484
-
文章历史
- 收稿日期: 2016-12-02
- 网络出版日期: 2018-10-18





2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
叶片形态的大小、寿命、冠层高度和光合特征等植物功能性状可以反映出植物对生长环境的响应和适应[1-6], 这些功能性状与植物对资源的获取利用效率及植株生物量密切相关[7]。叶片是种子植物制造有机养料的重要器官, 具有光合、蒸腾和呼吸的作用, 其形态大小和功能直接影响着植物对光的截取和对碳的获取, 并显著影响植物的相对生长速率[8-9]。已有的研究显示, 生长在光线充足生境中植物叶片的暗呼吸速率、比叶面积、光合能力及叶寿命均显著高于生长在光线较弱生境中的植物[10]。与喜光植物(或阳生叶)的叶片相比, 耐阴植物(或阴生叶)的叶片较薄, 比叶重较小、比叶面积较大, 光合能力、光饱和点、暗呼吸速率、叶绿素a/b偏低, 叶绿素浓度则较高[11-12]。植物耐阴性是植物为适应遮阴环境的一种复合性状, 在植物群落动态中起重要作用, 被认为是了解森林群落演替和动态的主要特征[13-14]。目前国内外对耐阴性的研究涵盖了从个体到生态系统水平的多个层次, 包括植物的形态特征、生理特征、耐阴性等级评估、耐阴性与个体发育等内容, 并有碳获得假说、胁迫耐受假说和碳储备假说等机理理论, 但植物耐阴性受特定的生态环境、生长季持续时间等多重压力的影响[13, 15], 植物耐阴性有很大的差异, 如何选择便于测定的指标还需要更深入的研究。以往植物耐阴性的研究主要集中在农作物、园林植物、地被植物、花卉、牧草等方面[12, 16], 在野外自然条件下, 对林下植物耐阴性的研究还较少报道。
桉树 (Eucalyptus urophylla)是联合国粮农组织推荐的三大速生树种之一, 也是我国南方重要的战略用材树种[17]。但由于长期以来为提高林木的生长量和出材率, 定期除草、抚育等人为干扰导致了桉树林生物多样性的锐减;另外, 桉树种植密度通常较大, 幼林在1年内可达到完全郁闭, 致使其林下植被的更新和生长受阻[18-20]。对华南地区尾叶桉林的林下植物生长调查发现, 在种植初期冠层未形成前, 芒萁、岗松、桃金娘、野牡丹等植物会快速生长, 但随着尾叶桉林冠层的郁闭, 梅叶冬青和黄栀子等种类得到发展, 林下植物种类、个体数量出现剧烈的消长, 推测这种林下个体数量的消长可能与林下环境的过滤和筛选作用密切相关[21-22]。因此, 研究尾叶桉林下植物的生长情况及其耐阴性, 不仅有利于认识尾叶桉林下植物生长过程对环境变化的响应和适应, 而且有利于了解林下植物在发展过程中的群落演替动态和生物多样性形成机理。基于此, 本研究开展尾叶桉林林下5种植物的叶片形态及其光合特征、以及植株生物量和个体数量消长等研究, 通过分析尾叶桉林林下植物的这些功能性状指标与耐阴性特征, 阐明尾叶桉林林下主要植物种类的生理生态特性和生长适应性, 为促进桉树林的可持续发展和合理经营提供参考。
1 研究地区概况本研究在广东鹤山森林生态系统国家野外科学观测研究站(鹤山站)共和样区开展。鹤山站位于112°53′E, 22°40′N, 低丘地势, 气候温暖多雨, 年均气温21.70℃, 其中最高月均温29.2℃, 最低月均温12.6℃。平均年降雨量1801mm, 年蒸发量1700mm, 年平均相对湿度78%。地带性土壤为赤红壤。土壤pH为4.0, 土壤有机质含量为0.56%—1.64%。
尾叶桉人工林是2005年春季种植, 株行距为3m×3m, 种植后前2年按正常的林业抚育管理, 其后自然生长。根据样方调查, 尾叶桉树人工林下的主要植物有山苍子(Litsea cubeba)、桃金娘(Rhodomyrtus tomentosa)、梅叶冬青(Ilex asprella)、栀子(Gardenia jasminoides)、野牡丹(Melastoma candidum)、鬼灯笼(Clerodendron fortunatum)、岗松(Baeckea frutescens)、米碎花(Eurya chinensis)、了哥王(Wikstroemia indica)等。草本植物种类主要有芒萁(Dicranopteris dichotoma)、扇叶铁线蕨(Adiantum flabellulatum)、芒草(Miscanthus sinensis)等。
2 研究方法 2.1 样方调查与生物量计算在尾叶桉林内建立3个30m×30m永久样地。于2006年10月和2011年10月分别调查了各个永久样地中的灌木层生长情况。用相邻格子法调查了9个5m×5m小样方内的所有灌木植物的种类、高度、地径、丛数以及每丛的分枝数。统计各种植物的丛(株)数、高度、地径和冠幅等。树高、冠幅用钢卷尺测定, 地径(离地面10cm)用游标卡尺测定。
灌木层植物的生物量测定用类似于乔木的异速生长式方法。根据2011年野外样方调查数据统计结果, 每种灌木各选择标准木3株, 大于和小于标准木各2株, 共7株进行全株收获, 以此数据建立枝、叶和根各器官的生物量与地径和株高的异速生长关系式, 再依生长式求算生物量[23]。
各器官的生物量与地径和株高的生长关系式为:
W=a(D2H)b
式中, W为灌木各器官的干生物量, D为基径, H为树高, a和b为常数。
再依样地调查数据(地径、高度和数量)求算各物种的生物量, 本文所述生物量均指干生物量。
2.2 光合参数测定根据前面的样方调查数据选择林下自然分布较多, 同时有较大经济价值和药用价值的植物种类桃金娘、栀子、梅叶冬青、山苍子和野牡丹等5种作为研究对象, 开展进一步的叶片形态特征和光合测定。于2013年10月, 用LI 6400便携式光合作用系统(LI COR, Inc, USA)分别测定了5种林下植物成熟叶片光合作用的光响应曲线。
尾叶桉人工林种植规律均匀, 根据2013年样方调查时测定, 3个固定样地的叶面积指数在1.12—1.3之间, 林内的光照环境较为相似。在光合测定时, 选取样方内林冠基本一致的5种植物进行原位连体测定。测定时间为晴朗天气的9:00—12:00。测定的叶片为适当远离桉树、植株健康的树冠上部长势优良的枝条第3—4片健康叶。测定前所有叶片材料均处在自然光下或用Li-6400自带光源(6400-02 LED)以1000μmol m-2 s-1光强充分诱导, 使用开放式气路, 用CO2小钢瓶供应CO2气体, 控制叶室内CO2浓度为400μmol/mol, 用仪器自带的温度控制器设定叶片温度为25℃左右。测定时光强由强到弱, 依次设定光量子通量密度为1500、1200、1000、800、500、300、200、100、80、60、40、20μmol m-2 s-1, 待数值稳定后, 记录每个光强下的Pn。测定结果用Von Bertalanffy非线性模型拟合出每种植物的Pn-PFD(光合-光响应)曲线方程[24]。
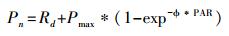
|
式中:Pn为实际测定的光合速率(μmol m-2 s-1), Rd为暗呼吸速率(μmol m-2 s-1), Pmax为最大光合速率(μmol m-2 s-1), ϕ为光合作用的表观量子效率(mol/mol)。
根据Pmax值(Psat, 为Pmax的95%)[25], 算出光饱和点(LSP, mol m-2 s-1)和光补偿点(LCP, mol m-2 s-1)[26]。
2.3 叶片特征的测定光合速率测定完成后, 在叶片中部用钻孔器取10片小圆片, 用80%丙酮提取, 采用紫外分光光度计(Lambda650, USA)在波长663、645、470nm下测定吸光值, 计算叶绿素叶a、b含量和类胡萝卜含量。
同时随机摘取成熟叶30片, 分别用电子天平称重, 用LI-3000C叶面积仪测定叶片的长、宽、叶面积, 用游标卡尺测定叶片中部的厚度(避开主叶脉)。上述项目测定完成后叶片在105℃处理30min, 再用80℃烘干24h至恒重, 称重并根据数据计算比叶面积(specific leaf area, SLA), SLA=叶片单面面积/叶片干重。
2.4 数据分析数据统计分析采用SPSS 22.0软件、Excel 2010软件和Sigma Plot 13.0, 测定结果以平均值表示, 采用单因素方差分析不同灌木种类各指标间的显著性差异。
3 结果与分析 3.1 5种林下植物叶片特征叶片特征指标是植物生长特征之一。耐阴植物对弱光照的适应性表现在叶面积的增加和非同化器官相对重量的减少, 叶片通常变薄, 比叶面积大[27-28]。由表 1可看出, 5种林下植物中, 野牡丹的宽度、厚度、单片叶面积等指标值最大, 梅叶冬青的长度、厚度、单位叶面积鲜重等指标值最小;其余3种, 桃金娘、栀子和山苍子介于野牡丹和梅叶冬青之间。梅叶冬青的比叶面积较大, 而桃金娘和野牡丹较小, 栀子和山苍子介于它们之间。从叶片大小、叶厚度、比叶面积等指标可粗略得出, 梅叶冬青比桃金娘和野牡丹较为耐阴。
| 项目 Items |
桃金娘 Rhodomyrtus tomentosa |
栀子 Gardenia jasminoides |
梅叶冬青 Ilex asprella |
山苍子 Litsea cubeba |
野牡丹 Melastoma candidum |
| 叶长Leaf length/cm | 8.31(0.84)C | 11.63(1.90)A | 5.60(0.72)D | 8.02(1.06)C | 10.03(1.26)B |
| 叶宽Leaf width/cm | 3.10(0.46)C | 4.70(0.70)B | 3.08(0.38)C | 2.65(0.40)D | 5.50(0.95)A |
| 单片叶面积Leaf area/cm2 | 16.40(3.18)C | 33.71(9.63)B | 10.36(2.48)C | 13.37(3.38)C | 38.73(10.68)A |
| 叶厚度Leaf thickness/mm | 0.38(0.04)B | 0.25(0.03)C | 0.17(0.03)E | 0.19(0.02)D | 0.56(0.05)A |
| 单片叶干重Leaf dry weight/g | 0.15(0.04)B | 0.16(0.04)B | 0.04(0.02)C | 0.06(0.02)C | 0.33(0.09)A |
| 含水量Water content/% | 57(3)C | 72(2)A | 72(3)A | 68(2)B | 68(2)B |
| 比叶面积Specific leaf area/(cm2/g) | 108.60(8.52)E | 203.77(21.53)C | 263.06(67.04)A | 223.79(24.31)B | 117.36(69.31)D |
| 数据为平均值(标准差), 样本n=30, 同行不同大写字母表示5种植物的叶片特性在(P≤0.01)水平上差异显著 | |||||
叶绿素含量也是植物叶片适应环境的特征之一。从表 2可看出, 5种灌木的叶绿素含量在29.57—45.40μg/cm2之间, 山苍子的叶绿素a、b和类胡萝卜素含量都明显比其他4种高, 野牡丹的叶绿素b含量比其他种明显低。由表 2还可看出, 除桃金娘与梅叶冬青之间的叶绿素a/b比值没有显著差异外, 其他种间的比值都有显著差异, 其中野牡丹的比值最高达3.85, 梅叶冬青最低为2.11。5个种中只有野牡丹的叶绿素a/b比值超过3, 其余的都在2—3之间。
| 项目 Items |
桃金娘 Rhodomyrtus tomentosa |
栀子 Gardenia jasminoides |
梅叶冬青 Ilex asprella |
山苍子 Litsea cubeba |
野牡丹 Melastoma candidum |
| 叶绿素a Chlorophyll-a | 23.11(1.24)B | 24.71(1.74)B | 23.55(1.20)B | 32.77(1.44)A | 23.48(1.40)B |
| 叶绿素b Chlorophyll-b | 10.67(0.48)B | 10.36(0.45)B | 11.18(0.61)B | 12.64(0.37)A | 6.09(0.37)C |
| 叶绿素a+b Chlorophyll a+b | 33.78(1.72)B | 35.07(2.12)B | 34.72(1.75)B | 45.40(1.80)A | 29.57(1.76)C |
| 叶绿素a/b Chlorophyll a/b | 2.16(0.02)D | 2.38(0.11)C | 2.11(0.06)D | 2.59(0.04)B | 3.85(0.04)A |
| 类胡萝卜素Carotenoids | 3.51(0.23)C | 4.26(0.26)B | 1.52(0.23)D | 6.4(0.29)A | 4.11(0.24)B |
| 数据为平均值(标准差), 样本n=3, 同行不同大写字母之间表示5种植物的叶绿素含量在(P≤0.01)水平上差异显著 | |||||
植物的光合作用是揭示植物生长的重要指标。从图 1可见, 5种林下植物叶片的净光合速率都随着光照的增强而增大。桃金娘、野牡丹和山苍子的净光合速率随光照的增强迅速增大, 其光饱和点较高;梅叶冬青和栀子的光合速率也随光照的增强而增大, 但其斜率比野牡丹和山苍子大, 并很快就达到光饱和点。

|
| 图 1 5种林下植物叶片的光合-光响应曲线(平均数±标准差, n=3) Fig. 1 The photosynthetic light response curve of five understory plants (mean±SD, n=3) |
从表 3中可见5种林下植物中, 最大净光合速率较高的种类有野牡丹、山苍子、桃金娘, 分别达到(16.42±0.31)、(14.64±4.64)、(14.53±1.79)μmol m-2 s-1, 梅叶冬青和栀子最大净光合速率较小, 分别为(9.50±0.41)μmol m-2 s-1和(11.25±0.16)μmol m-2 s-1。野牡丹的暗呼吸速率较大, 而桃金娘、山苍子、梅叶冬青、栀子的暗呼吸速率稍小, 且它们之间的差异不大。在光能利用方面, 梅叶冬青和栀子的表观量子效率较大, 而山苍子、野牡丹、栀子较小。5种林下植物中山苍子、野牡丹、桃金娘的光补偿点较大, 分别为(26±2)、(21±2)、(19±2)μmol m-2 s-1, 而栀子和梅叶冬青的光补偿点只有(15±1)μmol m-2 s-1和(16±1)μmol m-2 s-1。
| 项目 Items |
桃金娘 Rhodomyrtus tomentosa |
栀子 Gardenia jasminoides |
梅叶冬青 Ilex asprella |
山苍子 Litsea cubeba |
野牡丹 Melastoma candidum |
| Rd/(μmol m-2 s-1) | 0.98(0.053)b | 1.09(0.15)b | 0.72(0.05)c | 1.03(0.085)b | 1.37(0.12)a |
| Pnmax/(μmol m-2 s-1) | 14.53(1.79)b | 11.25(0.16)c | 9.50(0.41)d | 14.64(0.32)b | 16.42(0.31)a |
| LSP/(μmol m-2 s-1) | 716(166)ab | 462(8)c | 567(86)bc | 941(186)a | 669(137)bc |
| LCP/(μmol m-2 s-1) | 19(2)bc | 15(1)d | 16(1)cd | 26(2)a | 21(2)b |
| AQY/(mol/mol) | 0.0058(0.0008)ab | 0.0066(0.0001)a | 0.0065(0.0006)a | 0.0042(0.0003)c | 0.0052(0.0007)bc |
| Rd:暗呼吸速率Dark respiration rate;Pnmax:最大净光合速率Maximum net photosynthetic rate;LSP:光饱和点Light saturation point;LCP:光补偿点Light compensation point;AQY:表观量子效率Apparent quantum efficiency;数据为平均值(标准差), 样本n=3, 同行不同小写字母之间表示5种植物的光合参数在(P≤0.05)水平上差异显著 | |||||
植物耐阴性的影响因素众多, 从不同因素分析得到的结果可能不同。在生长环境、林下植被盖度和林冠郁闭度相似的前提下, 为了比较各指标的聚类分析结果, 本研究分别用叶片形态指标、光合参数指标和综合指标等作为变量对5种植物进行Person相关性组间联接聚类分析。其中叶片的形态结构指标为:叶片长、宽、厚度、单片叶面积、单片叶干重和比叶面积等;叶绿素含量指标为:叶绿素a、叶绿素b、叶绿素a/b值、类胡萝卜素、叶绿素a+b等;光合参数指标为:暗呼吸速率、最大净光合速率、光饱和点、光补偿点等;综合指标为:叶片长、宽、厚度、单片叶面积、单片叶干重、比叶面积、叶绿素a、叶绿素b、叶绿素a/b值、类胡萝卜素、叶绿素a+b、暗呼吸速率、最大净光合速率、光饱和点、光补偿点和表观量子率等。分析结果如图 2, 当用叶片形态结构指标聚合距离为25时可分2类:梅叶冬青、山苍子、桃金娘和栀子为一类, 野牡丹为单独一类(图 2A)。当用叶绿素含量指标时结果类似于叶片形态结构指标的结果(图 2B), 这二种结果都不能反映植物的实际情况,可见单独用叶片形态和叶绿素含量指标进行聚类分析不能很好地区分这5种植物的耐阴性。当用叶光合参数指标时梅叶冬青、山苍子、桃金娘为一类, 栀子和野牡丹为一类, 与叶形态和叶绿素含量指标聚类有差异, 栀子分为不耐阴生类与实际有些不符。当用综合指标聚类时, 5种林下植物也分为2类, 桃金娘、野牡丹、山苍子3种为1类, 这3种林下植物为较不耐阴;栀子、梅叶冬青为第2类, 这类植物较耐阴, 综合指标聚类分析结果与实际情况较相符。

|
| 图 2 5种植物耐阴性的聚类分析 Fig. 2 Dendrogram of shade-tolerance of plants A:叶形态指标聚类Cluster analysis of leaf morphology indicators;B:叶绿素含量指标聚类Cluster analysis of chlorophyll content indicators;C:为光合参数指标聚类Cluster analysis of photosynthetic parameters;D:为综合指标聚类Cluster analysis of composite indicators |
根据植株收获的数据建立了5种植物的各器官干生物量与树高和地径(胸径)的异速生长关系式。如表 4:
| 项目 Items |
桃金娘 Rhodomyrtus tomentosa |
栀子 Gardenia jasminoides |
梅叶冬青 Ilex asprella |
山苍子* Litsea cubeba |
野牡丹 Melastoma candidum |
| 枝W枝 Branch/WB | 0.0264(D2H)1.4868 | 0.4033(D2H)0.8641 | 3.9094(D2H)0.6362 | 10.1(D2H)1.0363 | 0.151(D2H)1.1474 |
| 叶W叶 Leaf/WL | 0.00001(D2H)2.5862 | 0.5262(D2H)0.5596 | 0.6865(H)0.5785 | 16.7(D2H)0.701 | 0.3305(D2H)0.6878 |
| 根W根 Root/WR | 0.2277(D2H)1.0502 | 0.2399(D2H)0.8814 | 3.5439(D2H)0.5916 | 18.9(D2H)0.8592 | 1.1407(D2H)0.6912 |
| *山苍子生长式的单位:W:干生物量Dry biomass/g, H:树高Height/m, D:胸径Diameter at breast height/cm;桃金娘、栀子、梅叶冬青、野牡丹生长式单位:W:干生物量Dry biomass/g, H:树高Height/cm, D:地径Ground diameter /cm | |||||
根据样方调查数据和上面的生长关系式计算, 2006年尾叶桉林下的桃金娘、栀子、梅叶冬青、山苍子和野牡丹等5种林下植物的总干生物量分别为:103.45、160.08、80.29、217.76、0.87kg/hm2(表 5), 其中山苍子生物量最大, 其次是栀子和桃金娘, 最小是野牡丹。2011年, 5种林下植物的生物量都有不同程度的增加, 其中增幅最大的是野牡丹, 增加达74倍, 从0.87kg/hm2增加到64.4kg/hm2;其次是桃金娘, 增加达18倍, 从103.45kg/hm2增加到1884.72kg/hm2;增幅最小的是栀子, 只增加1.3倍, 从160.08kg/hm2增加到211.18kg/hm2。山苍子和梅叶冬青分别增加6和5倍。结合表 6的个体数量变化分析可知, 野牡丹、桃金娘和山苍子的个体数量是减少的, 它们林下生物量的增加主要是由植株的高度和地径增加引起的。栀子和梅叶冬青生物量的增加,其个体数量、株高和地径增长都有贡献。2011年尾叶桉林下5种林下植物生物量从大到小的排序为:桃金娘>山苍子>梅叶冬青>栀子>野牡丹。
| 树种 Tree species |
2006年 | 2011年 | |||||
| 地上生物量 Aboveground biomass |
地下生物量 Underground biomass |
总生物量 Total biomass |
地上生物量 Aboveground biomass |
地下生物量 Underground biomass |
总生物量 Total biomass |
||
| 桃金娘Rhodomyrtus tomentosa | 68.63 | 34.82 | 103.45 | 244.78 | 1639.94 | 1884.72 | |
| 栀子Gardenia jasminoides | 31.40 | 128.68 | 160.08 | 49.89 | 161.29 | 211.18 | |
| 梅叶冬青Ilex asprella | 55.88 | 24.41 | 80.29 | 218.94 | 121.61 | 403.55 | |
| 山苍子Litsea cubeba | 171.50 | 46.17 | 217.67 | 1096.95 | 232.10 | 1329.05 | |
| 野牡丹Melastoma candidum | 0.27 | 0.61 | 0.87 | 38.57 | 25.83 | 64.40 | |
| 种类 Tree species |
2006年 | 2011年 | |||||||
| 密度 Density/(丛/hm2) |
高 Height/cm |
地径 Ground diameter/cm |
株数 Number of stems/丛 |
密度 Density/(丛/hm2) |
高 Height/cm |
地径 Ground diameter/cm |
株数 Number of stems/丛 |
||
| 桃金娘Rhodomyrtus tomentosa | 520 | 71.5 | 0.62 | 3.4 | 400 | 114.4 | 0.95 | 1.7 | |
| 栀子Gardenia jasminoides | 190 | 100.5 | 0.78 | 2.8 | 250 | 106.2 | 0.88 | 1.5 | |
| 梅叶冬青Ilex asprella | 120 | 141.9 | 1.09 | 3.2 | 190 | 213.1 | 1.99 | 3.5 | |
| 山苍子Litsea cubeba | 1170 | 210.8 | 1.31(胸径) | 1 | 170 | 476.1 | 4.31(胸径) | 1 | |
| 野牡丹Melastoma candidum | 130 | 63.5 | 0.29 | 1.2 | 70 | 114.8 | 0.70 | 1.7 | |
对鹤山共和样地尾叶桉林的林下植被样方调查数据分析显示(表 6):山苍子、桃金娘和野牡丹的密度(从2006年到2011年), 单位面积丛数分别减少了73.4%、23.1%和46.2%, 而栀子和梅叶冬青分别增加了31.6%和58.3%。从每丛的分枝数来看, 野牡丹和梅叶冬青平均每丛的分枝数增加, 而桃金娘和栀子的每丛分枝数正逐渐减小。说明尾叶桉林从第2年到第7年的生长过程中, 山苍子、桃金娘和野牡丹的个体数量不断减少, 栀子和梅叶冬青的个体数量不断增加。
在株高方面, 除栀子的株高变化不大外, 其他种的株高都有较大的增长, 特别是山苍子其株高在2011年比2006年有较大的增加, 这可能与其小乔木的生物学特性有关[29]。5种林下植物的地径(胸径)都有不同程度的增长, 其中山苍子胸径和梅叶冬青的地径增长最大。
4 讨论 4.1 5种林下植物的耐阴特性植物的耐阴性是由植物的遗传特性和植物对外部光环境变化的适应性决定的, 是植物的重要性状。乔木下层生境一般光照较弱, 空间资源相对较小。林下植物作为乔林下层空间的生存者, 大多演化成较好的适应耐阴特征。叶片特征如叶片大小、叶厚度、比叶面积等常常被作为衡量植物的耐阴特性。一般认为耐阴植物的叶大而薄, 比叶面积较大[27-28]。特别是比叶面积指标, 许多学者认为同一个体或群落内, 受光越弱, 比叶面积越大, 所以比叶面积可作为叶遮阴度的指数使用[29-32]。本研究的5种林下植物中, 野牡丹叶片的宽度、厚度、单片叶面积和单片叶干重等指标值最大, 梅叶冬青的长度、厚度、单位叶面积干重等指标值最小;其余3种, 桃金娘、栀子和山苍子介于野牡丹和梅叶冬青之间。野牡丹和桃金娘的比叶面积相对较小, 而梅叶冬青、山苍子和栀子的比叶面积相对较大, 它们之间都有显著差异。用叶片形态特征指标叶片长、叶片宽、单片叶面积、叶厚度、单片叶干重、含水量和比叶面积作为聚类分析的因素变量进行分析, 结果是梅叶冬青、山苍子、桃金娘和栀子为一类, 野牡丹单独一类(图 2A)。可见叶片的叶片形态特性只能粗略地分析叶片的耐阴性, 叶片形态很多指标表现并不一致, 也就是说用叶片的形态特性没法较好地划分植物的耐阴特性。如山苍子的比叶面积和叶厚度等指标与耐阴特性并不完全相一致。虽然有些学者认为SLA对光照响应呈现相反的变化趋势[33], 但本研究的结果显示, 比叶面积与植物耐阴性的相关性不明显, 这可能与植物群落的发育程度相关。
叶绿素在植物的光合作用中扮演着重要的角色, 主要由叶绿素a和叶绿素b组成。由于两者的理化性质和吸收光谱的特性有所差异, 因此叶绿素a、b及叶绿素a/b是反映植物耐阴性的重要指标[34]。一般来说, 耐阴性强的植物叶绿素含量较高, Chl a/b比值较低(常小于3), 能充分利用蓝紫光, 以适应在遮阴的环境中生长, 而阳生植物Chla/b比值可达3以上[30, 35-37]。根据这一点, 本研究中野牡丹的叶绿素a/b比值超过3属于阳生植物, 而山苍子、桃金娘、栀子和梅叶冬青的叶绿素a/b值都在2—3之间, 属非阳生植物。但综合考虑光合参数和聚类分析的结果表明, 野牡丹、桃金娘和山苍子都属于阳生植物。任海等[38]的研究也认为桃金娘、山苍子的生长需要充足的阳光, 不能在透光率低于32%以下的林冠下生长。以叶片叶绿素含量特征指标作为聚类分析的因素变量进行分析, 结果与叶片形态指标相似, 梅叶冬青、山苍子、桃金娘和栀子为一类, 野牡丹单独一类(图 2B)。可见单独以叶绿素含量指标和叶片形态指标都不能很好地区分植物的耐阴性。
植物的光合参数是植物生长情况的重要参考参数。植物能否适应环境的生长, 光合参数就能较好地反映出来。通常阳生植物最大净光合速率较高, 光饱和点和补偿点也较大, 而耐阴植物相反[35]。5种植物的光合测定表明, 野牡丹、山苍子和桃金娘最大净光合速率较大, 梅叶冬青和栀子最大净光合速率较小。山苍子、野牡丹和桃金娘的光补偿点较大, 而栀子和梅叶冬青的光补偿点较小。再以叶片光合参数指标作为聚类分析的因素变量进行分析, 结果为梅叶冬青、山苍子、桃金娘为一类, 栀子和野牡丹为一类(图 2C)。这也与实际情况不太一致, 可见仅以光合参数指标也没法很好地分类判别植物的耐阴性。
聚类分析是统计学中常用的分类方法, 并被广泛的应用于科学研究中。曾小平等应用此方法对25种植物的耐阴性进行综合分类, 其正确率达到96%, 认为此方法具有较高的可信度[30]。为了更准确地对5种植物进行聚类, 本研究选择叶片形态特征指标, 叶绿素含量指标, 光合参数等综合指标作为聚类分析的因素变量进行分析, 结果表明:5种林下植物的耐阴性可分为两类;第1类为较阳生性的野牡丹、山苍子、桃金娘;第2类为较耐阴生性的梅叶冬青和栀子(图 2D)。这一结论较符合5种植物的实际情况。植物耐阴性是由多种因素相互作用构成的一个较为复杂的综合性状, 用单一类的指标很难准确评定, 应用综合指标的已成为主流方法, 然而如何选择合适的指标还没有统一标准。本研究以叶片形态、叶绿素含量和光合参数等形态和生理指标作为评价指标, 运用聚类分析方法, 能较好地区分林下植物的耐阴性, 具有简便易行, 结果可靠等特点, 可在实践中推广。
4.2 5种林下植物个体数量的消长5种林下植物的生长特征表明, 山苍子、野牡丹和桃金娘的单位面积密度不断减小, 预计随着尾叶桉林冠层郁闭度越来越大, 这3种植物会逐渐消亡, 最终退出群落, 梅叶冬青和栀子作为耐阴植物, 其单位面积密度不断增加, 在林内能够长期保存, 最终发展为林下主要种类。其中山苍子为落叶小乔木, 喜温湿的环境, 原生长在阳坡、采伐迹地、荒山灌丛和疏残林地的边缘[39-40]。在尾叶桉树种植初期(2006年), 其单位面积密度和生长表现都较其他4个种大;但随着尾叶桉种植年限的增加(2011年), 其单位面积密度大大下降, 虽然保存下来的个体能长成小乔木, 但其个体生长没有尾叶桉快, 与尾叶桉竞争资源处于劣势, 只能在林窗或林缘形成乔木的亚冠层。因此, 在桉树的短轮伐期经营模式下, 林窗和林缘可适当保留山苍子。野牡丹和桃金娘为矮小常绿灌木, 是阳性先锋树种。野牡丹喜光, 耐炎热;桃金娘稍耐阴, 遮阴时间过长则开花结实会减少乃至消亡[41]。随着桉树人工林群落的发展, 野牡丹和桃金娘的单位面积密度也大大下降, 可预见野牡丹和桃金娘也会逐渐衰退。因此, 在尾叶桉种植初期, 可适当地清理野牡丹和桃金娘种类, 减少其与尾叶桉竞争养分。有资料显示, 梅叶冬青常生于山地疏林、路旁灌丛或阔叶林中, 有一定的耐阴性[42]; 栀子为常绿灌木, 喜四季分明的亚热带湿润气候, 耐旱, 幼苗时喜阴[43]。本研究结果也表明梅叶冬青和栀子2种林下植物的单位面积丛数呈增加的趋势, 这2种植物能在桉树林下正常生长。因此, 梅叶冬青和栀子是适合桉树林下生长的植物种类;同时, 梅叶冬青和栀子具有一定的药用价值, 为提高桉树人工林的生物多样性, 建议在桉树林下植被管理过程中可适当保护和种植梅叶冬青和栀子。
| [1] | Mcintyre S, Lavorel S. Livestock grazing in subtropical pastures:steps in the analysis of attribute response and plant functional types. Journal of Ecology, 2001, 89(2): 209–226. DOI:10.1046/j.1365-2745.2001.00535.x |
| [2] | Royer D L, Miller I M, Peppe D J, Hickey L J. Leaf economic traits from fossils support a weedy habit for early angiosperms. American Journal of Botany, 2010, 97(3): 438–445. DOI:10.3732/ajb.0900290 |
| [3] | Scoffoni C, Rawls M, McKown A, Cochard H, Sack L. Decline of leaf hydraulic conductance with dehydration:relationship to leaf size and venation architecture. Plant Physiology, 2011, 156(2): 832–843. DOI:10.1104/pp.111.173856 |
| [4] | 雷羚洁, 孔德良, 李晓明, 周振兴, 李国勇. 植物功能性状、功能多样性与生态系统功能:进展与展望. 生物多样性, 2016, 24(8): 922–931. DOI:10.17520/biods.2015295 |
| [5] | 龚时慧, 温仲明, 施宇. 延河流域植物群落功能性状对环境梯度的响应. 生态学报, 2011(20): 6088–6097. |
| [6] | 刘晓娟, 马克平. 植物功能性状研究进展. 中国科学:生命科学, 2015, 45(4): 325–339. |
| [7] | 宝乐, 刘艳红. 东灵山地区不同森林群落叶功能性状比较. 生态学报, 2009, 29(7): 3692–3703. |
| [8] | Wright I J, Reich P B, Westoby M, Ackerly D D, Braruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas ML, Niinemets V, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821–827. DOI:10.1038/nature02403 |
| [9] | Milla R, Reich P B. The scaling of leaf area and mass:the cost of light interception increases with leaf size. Proceedings of the Royal Society B:Biological Sciences, 2007, 274(1622): 2109–2114. DOI:10.1098/rspb.2007.0417 |
| [10] | Wright J P, Naeem S, Hector A, Lehman C, Reich P B, Schmid B, Tilman D. Conventional functional classification schemes underestimate the relationship with ecosystem functioning. Ecology Letter, 2006, 9(2): 111–120. DOI:10.1111/j.1461-0248.2005.00850.x |
| [11] | Niinemets U, Kull O, Tenhunen J D. An analysis of light effects on foliar morphology, physiology, and light interception in temperate deciduous woody species of contrasting shade tolerance. Tree Physiology, 1998, 18(10): 681–696. DOI:10.1093/treephys/18.10.681 |
| [12] | 安锋, 林位夫. 植物耐荫性研究的意义与现状. 热带农业科学, 2005, 25(2): 68–72. |
| [13] | Valladares F, Niinemets V. Shade tolerance, a key plant feature of complex nature and consequences. Annual Review of Ecology, Evolution, and Systematics, 2008, 39(1): 237–257. DOI:10.1146/annurev.ecolsys.39.110707.173506 |
| [14] | Baltzer J L, Thomas S C. Determinants of whole-plant light requirements in Bornean rain forest tree saplings. Journal of Ecology, 2007, 95(6): 1208–1221. DOI:10.1111/jec.2007.95.issue-6 |
| [15] | 殷东生, 沈海龙. 森林植物耐荫性及其形态和生理适应性研究进展. 应用生态学报, 2016, 27(8): 2687–2698. |
| [16] | 付景, 李潮海, 赵久然, 马丽, 刘天学. 玉米品种耐阴性指标的筛选与评价. 应用生态学报, 2009, 20(11): 2705–2709. |
| [17] | 韦东艳, 杜阿朋, 谢耀坚, 贺亮亮, 张沛健. "近自然林业"在桉树人工林经营中的应用. 桉树科技, 2010, 27(2): 48–54. |
| [18] | 项东云. 华南地区桉树人工林生态问题的评价. 广西林业科学, 2000, 29(2): 57-64, 86-86. |
| [19] | 陈秋波. 桉树人工林生物多样性研究进展. 热带作物学报, 2001, 22(4): 82–90. |
| [20] | 赵庭香, 麦昌金. 桉树林生态环境的变化. 桉树科技, 1988, 5(1): 31–33. |
| [21] | Díaz S, Noy-Meir I, Cabido M. Can grazing response of herbaceous plants be predicted from simple vegetative traits?. Journal of Applied Ecology, 2001, 38(3): 497–508. DOI:10.1046/j.1365-2664.2001.00635.x |
| [22] | 孟婷婷, 倪健, 王国宏. 植物功能性状与环境和生态系统功能. 植物生态学报, 2007, 31(1): 150–165. |
| [23] | 张志良, 瞿伟菁. 植物生理学实验指导(第三版). 北京: 高等教育出版社, 2003. |
| [24] | Horton J L, Neufeld H S. Photosynthetic responses of Microstegium vimineum (Trin.) A. Camus, a shade-tolerant, C4 grass, to variable light environments. Oecologia, 1998, 114(1): 11–19. DOI:10.1007/s004420050414 |
| [25] | Quero J L, Villar R, Marañón T, Zamora R. Interactions of drought and shade effects on seedlings of four Quercus species:physiological and structural leaf responses. New Phytologist, 2006, 170(4): 819–834. DOI:10.1111/nph.2006.170.issue-4 |
| [26] | 许大全. 光合作用效率. 上海: 上海科学技术出版社, 2002. |
| [27] | 采利尼克尔. 木本植物耐阴性的生理学原理. 王世绩, 译. 北京: 科学出版社, 1986. |
| [28] | 白伟岚, 任建武, 苏雪痕. 八种植物耐阴性比较研究. 北京林业大学学报, 1999, 21(3): 46–52. |
| [29] | 陈庆之, 蔡淑珍. 山苍子植物学特性的研究. 经济林研究, 1984, 2(2): 52–58. |
| [30] | 曾小平, 赵平, 蔡锡安, 饶兴权, 刘惠, 马玲, 李长洪. 25种南亚热带植物耐阴性的初步研究. 北京林业大学学报, 2006, 28(4): 88–95. |
| [31] | Westoby M, Falster D S, Moles A T, Vesk P A, Wright I J. Plant ecological strategies:some leading dimensions of variation between species. Annual Review of Ecology and Systematics, 2002, 33(1): 125–159. DOI:10.1146/annurev.ecolsys.33.010802.150452 |
| [32] | Wright S J, Muller-Landau H C, Condit R, Hubbell S P. Gap-dependent recruitment, realized vital rates, and size distributions of tropical trees. Ecology, 2003, 84(12): 3174–3185. DOI:10.1890/02-0038 |
| [33] | Malavasi U C, Malavasi M M. Leaf characteristics and chlorophyll concentration of Schyzolobium parahybum and Hymenaea stilbocarpa seedlings grown in different light regimes. Tree Physiology, 2001, 21(10): 701–703. DOI:10.1093/treephys/21.10.701 |
| [34] | 孙艳, 高海顺, 管志勇, 陈素梅, 房伟民, 陈发棣. 菊花近缘种属植物幼苗耐阴特性分析及其评价指标的确定. 生态学报, 2012, 32(6): 1908–1916. |
| [35] | Larcher W. Physiological Plant Ecology.4th ed. Berlin Heidelberg: Springer-Verla, 2003. |
| [36] | Kozlowski T T, Pallardy S G. Physiology of Woody Plants.2nd ed. New York: Academic Press, 1997. |
| [37] | 王沙生, 高荣孚, 吴贯明. 植物生理学(第二版). 北京: 中国林业出版社, 1991. |
| [38] | 任海, 彭少麟, 孙谷畴, 余作岳. 广东中部两种常见灌木的生态学比较. 植物生态学报, 1997, 21(4): 386–392. |
| [39] | 陈卫军, 龚洵胜, 游小敏. 山苍子播种繁殖及扦插育苗初探. 经济林研究, 2004, 22(4): 59–60. |
| [40] | 钟奕灵, 谢金兰, 林立. 山苍子的价值及造林技术. 农技服务, 2011, 28(7): 1070-1070, 1079-1079. |
| [41] | 赵志刚, 程伟, 郭俊杰. 桃金娘的资源利用与人工培育. 广西林业科学, 2006, 35(2): 70–72. |
| [42] | 中国科学院中国植物志编辑委员会. 中国植物志. 北京: 科学出版社, 1999. |
| [43] | 曾智勇, 袁平新. 黄栀子栽培. 中国林业, 2011: 58–58. |