 2018, Vol. 38
2018, Vol. 38文章信息
- 唐一思, 石慰, 曾文豪, 郑维艳, 曹坤芳.
- TANG Yisi, SHI Wei, ZENG Wenhao, ZHENG Weiyan, CAO Kunfang.
- 广西天然林中藤本植物区系组成及系统发育多样性
- Floristic composition and phylogenetic diversity of climbing plants in natural forests across Guangxi
- 生态学报. 2018, 38(24): 8750-8757
- Acta Ecologica Sinica. 2018, 38(24): 8750-8757
- http://dx.doi.org/10.5846/stxb201808021642
-
文章历史
- 收稿日期: 2018-08-02
- 修订日期: 2018-11-30

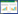




2. 广西大学, 亚热带农业生物资源保护与利用国家重点实验室, 南宁 530004
2. State Key Laboratory for Conservation and Utilization of Subtropical Agro-bioresources, Guangxi University, Nanning 530004, China
藤本植物是热带和亚热带森林的重要组分[1-5]。藤本植物依赖其它植物的支撑向上攀缘生长, 自己不长发达的支撑结构, 而把生物量更多地用于茎干的伸长生长, 以及叶片和根系生长, 因此, 在生态适应性和竞争力上具有独特优势[6], 对森林生态系统的生物多样性、结构、生产力及养分循环具有重要影响[7-8]。藤本植物对树木的缠绕会导致树木的物理损伤, 还会与其竞争有限的资源(如水分、养分和光)[9-10]。藤本植物通过影响共存树木的增补、生长、死亡, 进而影响森林的生物多样性、碳贮存与固定、生态系统服务与功能。
几个关键非生物因子, 如:温度、降雨、森林干扰等对天然林中藤本的多样性、丰富度和分布格局有重要影响[11-13]。温度是限制藤本向高纬度、高海拔分布的主要原因, 另有研究表明, 热带森林中藤本植物的丰富度与降水量呈负相关, 而与森林的降雨量的季节性强度正相关, 在具有明显干季的季节性热带森林中具有更高的丰富度[14]。
喀斯特是由碳酸盐岩构成的特殊生境, 中国喀斯特地貌分布广、面积大。其中以广西、贵州和云南东部所占的面积最大, 是世界上最大的喀斯特区之一。与非喀斯特生境相比, 喀斯特一方面生境异质性强, 岩石裸露, 土层薄, 多石缝、石沟, 因此保水能力差。另一方面, 由于碳酸盐岩易于风化、淋溶, 风化留下的碎屑少, 南方喀斯特地区土壤层浅, 生境破碎化严重, 生态系统抗干扰能力低。并且喀斯特土壤富含钙镁, 对有些植物有毒害作用。这些因素会限制一些植物的分布乃至森林的多样性, 尽管如此, 藤本植物在喀斯特植被中占有重要地位[15]。
藤本植物对于群落稳定性维持起着重要的作用, 但是, 长期以来对于藤本植物研究的关注度不够, 尤其对于其在生态系统生态过程中的作用缺乏系统深入的探讨。生物群落的构建是一个复杂的演替过程, 生态位理论认为生境过滤、竞争排斥等生态过程主导群落构建[16]。目前群落生态研究利用群落内物种的系统发育结构来揭示进化过程对群落的影响, 根据生态位保守理论, 如果是环境过滤为主的生态过程构建群落, 群落系统发育结构表现为聚集, 而系统发育结构发散可能是由竞争排斥作用主导构建群落[17-20]。随机的系统发育结构可能是生境过滤和竞争排斥共同作用的结果[21-22]。结合系统发育和功能性状结构的研究可以更全面地分析群落构建成因[23-24]。本研究以受太平洋热带季风强烈影响的广西地区天然林为研究对象, 通过整理与分析文献记载的森林群落藤本植物物种组成和多样性数据, 探究以下问题:(1)广西地区天然林中海拔1000 m以下的喀斯特和非喀斯特森林以及海拔1000 m以上非喀斯特森林中藤本植物区系组成有何特点?(2)3个不同生境中藤本植物系统发育多样性和结构有何差异?(3)3个生境的森林藤本物种多样性和系统发育多样性与结构指数与生长地点的年均温是否相关?
1 材料和方法 1.1 研究区域自然概况广西壮族自治区地处中国南方热带亚热带地区, 位于104°26′—112°04′E, 20°54′—26°24′N之间, 北回归线横贯全区中部。南北以贺州-东兰一线为界, 此界以北属中亚热带季风气候区, 以南属南亚热带季风气候区。全区各地极端最高气温为42.5℃, 极端最低气温为-8.4℃, 年平均气温在16.5—23.1℃之间。等温线基本上呈纬向分布, 气温由北向南递增, 由河谷平原向丘陵山区递减。广西是中国降水量最丰富地区之一。各地年降水量均在1070 mm以上, 大部分地区为1500—2000 mm。由于受冬、夏季风的交替影响, 广西降水量的季节变化不均, 干湿季分明。4—9月为雨季, 其降水量占全年降水量的70%—85%, 容易发生洪涝灾害;10月—次年3月为干季, 降水量仅占年降水量的15%—30%。
1.2 数据来源植物群落植物样地数据来源于《广西植被志要》[25]。共计355个样地[26], 海拔范围为10—2020 m, 纬度范围21.4°—25.9°N, 经度范围105.0°—111.9°E, 样地面积大部分为400 m2, 占总样地数的54.9%, 其次是500 m2和600 m2的样地, 合计占总样地数的39.1%。样方调查的森林主要为天然的成熟林, 将样地所涉及的森林分为三类生境类型:喀斯特森林、海拔1000 m以下非喀斯特森林、海拔1000 m以上非喀斯特森林。
根据《中国植物志》网络数据库(http://www.eflora.cn/)对同物异名和拉丁名进行了校正, 并把汇总后的藤本植物按科、属进行分类。根据《世界种子植物科的分布区类型系统》[26]、《中国被子植物科属综论》[27], 确定藤本植物的地理成分及科属的分布区类型, 并据此作统计分析。
从国家气象中心网站获取广西90个气象台站的气象监测数据, 通过每个台站1977—2006年的年均温计算得出每个台站30年的年均温。结合台站的海拔、纬度、经度构建回归方程(年平均温=-0.736×纬度-0.31×经度-0.005×海拔+72.794, R2=0.938, P < 0.001), 根据拟合出的回归方程, 通过样地的经纬度和海拔估测样地的年均温。
1.3 分析方法因为部分样方面积不一致, 采用Gleason(1926)指数来表示物种多样性的测度[28], 指数公式如下:
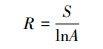
|
式中, A表示所研究的面积(m2);S表示面积A内的种数(个)。
本研究通过物种拉丁名, 利用Phylomatic和R软件构建系统发育树。将所有样地内全部藤本植物拉丁名输入到网页版Phylomatic中(http://phylodiversity.net/phylomatic/), 该软件以被子植物分类系统Ⅲ (APGⅢ)[29]为基础, 自动构建物种的系统发育拓扑结构[30]。在R软件中拟合分支结构, 得到系统发育树。计算每个样地的谱系多样性指数(PD), 分析物种在进化树中的位置, 计算物种间的谱系距离。假设物种分布随机, 在随机模型下将谱系距离标准化, 从而获得群落系统发育结构指数, 即净亲缘关系指数(NRI)和最近分类群指数(NTI)。群落系统发育结构指数的计算公式如下:
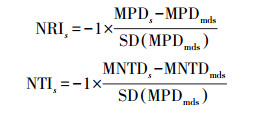
|
式中, NRIs和NTIs是样方群落系统发育结构指数, MPDs和MNTDs分别表示平均谱系距离的观测值和最近相邻谱系距离的观测平均值, 而MPDrnds和MNTDrnds表示通过软件随机模拟出的平均谱系距离和最近相邻谱系距离的平均值。最后用t检验判断3个不同类型生境系统发育是否有显著差异。
本研究数据利用Excel、SPSS与R语言3.4.2进行统计分析。
2 结果分析 2.1 广西天然林藤本植物区系组成本文收集到的广西地区天然林355个样方中藤本植物共计有344种, 55科, 130属。其中双子叶植物49科306种, 单子叶植物5科36种, 裸子植物仅有两种, 分别是买麻藤和小叶买麻藤。其中木质藤本300种, 占总种数的87.2%, 草质藤本44种, 占总种数的12.8%。
在科水平上, 豆科占比最大, 占该地区藤本总种数的12%(41种);蔷薇科、葡萄科、茜草科分别占总藤本种数的8%、8%、7%, 仅次于豆科。上述4科所含藤本总种数占该地区藤本物种数的35%。不低于10种的科还有薯蓣科、萝藦科等10科, 共有藤本193种, 占该地区藤本总种数的56.1%。不低于5种的科有紫金牛科、鼠李科等12科, 占藤本总种数的24.1%。其余的33科的物种数占总藤本总种数的19.8%。其中仅有一种的科有铁青树科、牛栓藤科等共13科。
2.2 不同生境的优势含藤科属广西喀斯特地区藤本植物共计230种, 45科, 95属。不低于10种以上的科依次是豆科、葡萄科、蔷薇科、萝藦科、薯蓣科、茜草科、夹竹桃科、防己科。在海拔1000 m以下非喀斯特的样地里藤本植物共计257种、47科、109属。不低于10种以上的科依次是豆科、葡萄科、茜草科、蔷薇科、番荔枝科、防己科、夹竹桃科、薯蓣科。在海拔1000米以上非喀斯特样地中藤本植物共计146种、35科、69属。不低于十种以上的科仅三科, 依次是豆科、蔷薇科、葡萄科(图 1)。

|
| 图 1 不同生境下藤本科组成 Fig. 1 The composition of climbing plants in different habitats |
仅在喀斯特森林样地出现的藤本植物科有翅子藤科、远志科、蓼科、爵床科、山柑科、马鞭草科。仅在非喀斯特海拔1000 m以下的样地中出现的植物科有海金沙科、菊科、五桠果科、龙胆科、檀香科、杨梅科、禾本科、梧桐科、樟科。而在海拔1000 m以上非喀斯特样地里所有科在其它两个生境都有分布。
在藤本植物科水平上, 喀斯特样地中世界广布型有11科, 热带型29科, 温带型3科, 亚洲型1科;在海拔1000 m以下的非喀斯特样地中, 世界广布型有13科, 热带型28科, 温带型3科, 亚洲型2科;在海拔1000 m以上的非喀斯特样地中, 世界广布型有9科, 热带型21科, 温带型3科, 亚洲型2科。
2.2.1 喀斯特森林在喀斯特森林样方出现的95个属的藤本植物可分为4大类分布类型和10个具体分布区类型(表 1)。其中世界属有4个, 占总属数的4.2%;热带分布属有75个, 占总属数的78.9%;温带分布属有10个, 占总属数的10.5%;亚洲分布属有6个, 占总属数的6.3%。
| 属分布区类型 Areal-types of genera | 喀斯特森林 Karst forests | 海拔1000 m以下的 非喀斯特森林 Non-karst forests with altitudes lower than 1000 m | 海拔1000 m以上的 非喀斯特森林 Non-karst forests with a ltitudes above 1000 m |
| 1 世界广布Widespread | 4.21 | 3.74 | 4.35 |
| 2 泛热带Pantropic | 29.47 | 30.84 | 30.43 |
| 3 东亚及热带南美间断Trop. & Subtr. E. Asia & (S.) Trop. Amer. disjuncted | 3.16 | 1.87 | 1.45 |
| 4 旧世界热带Old World Tropics | 13.68 | 14.95 | 15.94 |
| 5 热带亚洲至热带大洋洲Trop. Asia to Trop. Australasia Oceania | 7.37 | 5.61 | 4.35 |
| 6 热带亚洲至热带非洲Trop. Asia to Trop. Africa | 6.31 | 4.67 | 2.90 |
| 7 热带亚洲Trop. Asia | 18.95 | 18.69 | 14.49 |
| 8 北温带N. Temp. | 6.32 | 5.61 | 8.70 |
| 9 东亚及北美间断E. Asia & N. Amer. disjuncted | 4.21 | 5.61 | 5.80 |
| 10 东亚E. Asia | 6.32 | 7.48 | 10.14 |
| 11 中国特有Endemic to China | 0.93 | 1.45 |
就3个生境而言, 仅在喀斯特生境出现的属有球兰属(Hoya)、地锦属(Parthenocissus)、远志属(Polygala)、茅瓜属(Solena)、牛奶菜属(Marsdenia)、醉魂藤属(Heterostemma)、何首乌属(Fallopia)、马兰藤属(Dischidanthus)、眼树莲属(Dischidia)、老鼠簕属(Acanthus)、山柑属(Capparis)、扁蒴藤属(Pristimera)、翅子藤属(Loeseneriella)、花皮胶藤属(Ecdysanthera)、蓬莱葛属(Gardneria)、过江藤属(Phyla)。
2.2.2 海拔1000 m以下的非喀斯特森林该生境样方中出现107个属的藤本植物可分为4大类分布类型和11个具体分布区类型(表 1)。其中世界属有4个, 占总属数的3.7%, 热带分布属有82个, 占总属数的76.6%, 温带分布属有12个, 占总属数的11.2%, 亚洲分布属有9个, 占总属数的8.4%。
仅在该生境分布的属有:海金沙属(Lygodium)、斑鸠菊属(Vernonia)、葛属(Pueraria)、五味子属(Schisandra)、俞藤属(Yua)、黄藤属(Daemonorops)、锡叶藤属(Tetracera)、双蝴蝶属(Tripterospermum)、青牛胆属(Tinospora)、鹅掌柴属(Schefflera)、鸡爪簕属(Oxyceros)、榼藤属(Entada)、寄生藤属(Dendrotrophe)、马钱属(Strychnos)、裂果金花属(Schizomussaenda)、虎刺属(Damnacanthus)、黍属(Panicum)、紫玉盘属(Uvaria)、萝芙木属(Rauvolfia)、葎草属(Humulus)、土圞儿属(Apios)、昂天莲属(Ambroma)、黧豆属(Mucuna)、匙羹藤属(Gymnema)、纤冠藤属(Gongronema)、无根藤属(Cassytha)、盒果藤属(Operculina)。
2.2.3 海拔1000 m以上非喀斯特森林该生境森林样方中出现的69个属的藤本植物可分为4大类分布类型和11个具体分布区类型(表 1)。其中世界属有3个, 占总属数的4.3%;热带分布属有48个, 占总属数的69.6%;温带分布属有10个, 占总属数的14.5%;亚洲分布属有8个, 占总属数的11.6%。
仅在该生境分布的属有相思子属(Abrus)、罗汉果属(Siraitia)。
2.3 不同生境藤本植物物种多样性和系统发育多样性差异收集到的样地内的藤本物种丰富度和系统发育多样性(PD)呈极显著正相关关系(图 2)。协方差分析结果表明PD值与Gleason指数关系的回归线斜率在3种生境的森林中无显著差异, 而截距存在显著差异(P < 0.05)。在Gleason指数相同的情况下, 海拔在1000 m以下的非喀斯特森林中的PD值最高, 而在喀斯特森林中PD值最小。3个生境类型的群落Gleason指数(图 2)、PD与年平均温度不相关, NTI和NRI与年均温也不相关(图 3)。而3个生境类型的藤本群落Gleason指数与树木群落Gleason指数呈正的中度至弱相关关系(喀斯特森林R2=0.29, P < 0.0001;海拔小于1000 m的非喀斯特样地R2=0.10, P < 0.0001;海拔大于1000 m的非喀斯特样地R2=0.19, P < 0.0001)

|
| 图 2 不同生境下藤本植物Gleason指数与系统发育多样性和年均温之间的关系 Fig. 2 The relationship between Gleason index of climbing plants and the phylogenetic diversity, and relationship between mean annual temperature and Gleason index of climbing plants in the three habitats |

|
| 图 3 不同生境下年均温与藤本系统发育结构之间的关系 Fig. 3 The relationship between mean annual temperature and phylogenetic index of climbing plants in the three different forest habitats |
海拔1000 m以上的非喀斯特森林R2=95.7, P < 0.01;海拔1000 m以下的非喀斯特森林R2=91.1, P < 0.01;喀斯特森林R2=93.4, P < 0.01。
从样地内藤本植物系统发育结构来看, 海拔在1000 m以上的非喀斯特森林中76.5%的样地系统发育发散(NRI < 0), 23.5%的样地系统发育聚集(NRI>0);海拔在1000 m以下的非喀斯特森林中82.6%的样地系统发育发散(NRI < 0), 17.4%的样地系统发育聚集(NRI>0);喀斯特森林中29.4%的样地系统发育发散(NRI < 0), 70.6%的样地系统发育聚集(NRI>0)。
通过t检验分析, 不同生境的NRI、NTI和PD均存在着显著差异(P < 0.01)。
3 讨论 3.1 不同生境森林藤本植物区系组成差异相较于我国其他地区森林中的藤本植物区系组成, 广西地区的最大特点是热带型含藤属占比最大, 为75.4%, 而温带型藤本属占比较低, 仅占12.3%。藤本植物的导管比较宽大, 其水分运输易受低温和干旱胁迫的影响, 因而比较适宜热量丰富、雨量充足的气候条件[31]。广西地区地处北热带和亚热带, 全年温度偏高, 因此热带藤本植物在该地区占比大。
仅出现在一种生境的含藤属数量来看, 非喀斯特海拔1000 m以下生境内的含藤属最多, 共27个, 其中热带分布属有20个, 占总含藤属的74%, 而世界、温带、亚洲三类分布属仅有7属, 占总含藤属的26%。相较于喀斯特地区和非喀斯特1000 m以上地区, 海拔1000 m以下非喀斯特生境环境胁迫较弱, 物种多样性水平较高, 因此一些不适应低温、干旱胁迫生境的热带藤本属只能在该地区生长。
在非喀斯特1000 m以上地区, 温度下降明显, 森林类型由常绿阔叶林过度到常绿落叶阔叶混交林和少量针阔混交林, 仅出现在该生境的含藤属只有两种, 其中热带分布属和温带分布属各占一个, 由于藤本植物抵御气穴化的能力显著低于常绿树种, 在冬季面临着结冰的危险, 水分运输受阻, 从而限制了藤本在高海拔地区的生长[13]。所以在本研究中, 仅出现在海拔1000 m以上生境内的含藤属的数量显著下降, 热带分布属占比减小。喀斯特森林是一种容易缺水和富钙的生境[32], 限制了一些植物的分布, 研究结果显示仅出现在喀斯特地区的含藤属有16个。
3.2 不同生境类型系统发育间的差异藤本植物多样性与多种生态因子相关, 其中温度和降水量被认为是最关键的非生物因子[1, 5, 13], 也有研究表明藤本植物多样性随地形的变化而异, 与土壤pH值和岩石覆盖等重要的非生物因素有关[33]。本研究中藤本物种多样性和系统发育多样性(PD)与年均温均不相关, 可能是受到了不同样地内不同的地形以及乔木丰富度的影响。而在喀斯特森林内, 生境异质性明显和偏碱性、富钙镁的土壤同时影响着藤本植物物种多样性。
本研究结果显示, 在喀斯特森林藤本植物系统发育趋向聚集(图 3), 这是因为喀斯特地区独特的干旱生境以及破碎化的地形导致该地区植被生长相对困难, 多以小径级植物为主, 生境过滤作用在群落构建中占主导地位, 系统发育趋向于聚集。在非喀斯特地区大多是演替后期的成熟林, 树木径级较大, 群落竞争强, 竞争排斥作用占主导, 竞争排斥作用会使生态位相近的物种分离, 因此藤本植物系统发育表现为发散。从3种不同生境的PD值的比较来看, 在Gleason指数相同的情况下, 海拔在1000 m以下的非喀斯特森林中的藤本植物PD值最高, 而在环境胁迫严重的喀斯特森林中藤本PD值最小。季节性干旱显著的热带森林往往具有非常高的藤本植物丰富度[1, 14], 而喀斯特生境广泛分布的碳酸盐岩成分使得藤本植物难以利用深层地下水, 所以土壤层浅、保水性差的生境特点会使藤本植物受到一定程度的干旱胁迫, 土壤富钙镁对一些植物有毒害作用, 导致系统发育多样性降低。
由于温度的制约, 中山地带森林藤本多样性较低, 仅在中山地带森林中出现的含藤属很少, 温带成分占比大。本研究发现喀斯特森林藤本植物的系统发育结构趋向于聚集, 表明在该生境控制藤本植物的生态过程主要是生境过滤作用。在非喀斯特地区藤本植物的系统发育结构趋向于发散, 表明在非喀斯特地区竞争排斥作用在藤本植物群落构建中具有主导地位。本文的研究结果对于热带亚热带森林藤本植物的物种多样性组成和维持机制提供了新认识。
| [1] | Gentry A H. The distribution and evolution of climbing plants//Putz F E, Mooney H A, eds. The Biology of Vines. Cambridge: Cambridge University Press, 1991: 3-52. |
| [2] | Schnitzer S A, Mangan S A, Dalling J W, Baldeck C A, Hubbell S P, Ledo A, Muller-Landau H, Tobin M F, Aguilar S, Brassfield D, Hernandez A, Lao S, Perez R, Valdes O, Yorke S R. Liana abundance, diversity, and distribution on Barro Colorado island, Panama. PLoS One, 2012, 7(12): e52114. DOI:10.1371/journal.pone.0052114 |
| [3] | 蔡永立, 宋永昌. 中国亚热带东部藤本植物的多样性. 植物科学学报, 2000, 18(5): 390–396. DOI:10.3969/j.issn.2095-0837.2000.05.008 |
| [4] | 颜立红, 祁承经, 刘小雄. 中国亚热带中部藤本植物区系的基本特点. 中南林业科技大学学报, 2006, 26(4): 36–41. DOI:10.3969/j.issn.1673-923X.2006.04.008 |
| [5] | Hu L, Li M G, Li Z. Geographical and environmental gradients of lianas and vines in China. Global Ecology and Biogeography, 2010, 19(4): 554–561. |
| [6] | 祁承经, 颜立红, 彭春良. 异军突起的藤本植物. 武汉植物学研究, 2007, 25(4): 381–395. DOI:10.3969/j.issn.2095-0837.2007.04.012 |
| [7] | Schnitzer S A, Carson W P. Lianas suppress tree regeneration and diversity in treefall gaps. Ecology Letters, 2010, 13(7): 849–857. DOI:10.1111/ele.2010.13.issue-7 |
| [8] | Tymen B, Réjou-Méchain M, Dalling J W, Fauset S, Feldpausch T R, Norden N, Phillips O L, Turner B L, Viers J, Chave J. Evidence for arrested succession in a liana-infested Amazonian forest. Journal of Ecology, 2016, 104(1): 149–159. DOI:10.1111/1365-2745.12504 |
| [9] | 陈亚军, 曹坤芳, 蔡志全. 两种光强下木质藤本与树木幼苗的竞争关系. 植物生态学报, 2008, 32(3): 639–647. DOI:10.3773/j.issn.1005-264x.2008.03.013 |
| [10] | Van Der Heijden G M F, Phillips O L. Environmental effects on Neotropical liana species richness. Journal of Biogeography, 2009, 36(8): 1561–1572. DOI:10.1111/jbi.2009.36.issue-8 |
| [11] | Laurance W F, Andrade A S, Magrach A, Camargo J L C, Valsko J J, Campbell M, Fearnside P M, Edwards W, Lovejoy T E, Laurance S G. Long-term changes in liana abundance and forest dynamics in undisturbed Amazonian forests. Ecology, 2014, 95(6): 1604–1611. DOI:10.1890/13-1571.1 |
| [12] | Martínez-Izquierdo L, García M M, Powers J S, Schnitzer S A. Lianas suppress seedling growth and survival of 14 tree species in a Panamanian tropical forest. Ecology, 2016, 97(1): 215–224. DOI:10.1890/14-2261.1 |
| [13] | Balfour D A, Bond W J. Factors limiting climber distribution and abundance in a Southern African forest. The Journal of Ecology, 1993, 81(1): 93–100. DOI:10.2307/2261227 |
| [14] | Schnitzer S A, Bongers F. The ecology of lianas and their role in forests. Trends in Ecology & Evolution, 2002, 17(5): 223–230. |
| [15] | 李先琨, 苏宗明, 吕仕洪, 欧祖兰, 向悟生, 区智, 陆树华. 广西岩溶植被自然分布规律及对岩溶生态恢复重建的意义. 山地学报, 2003, 21(2): 129–139. DOI:10.3969/j.issn.1008-2786.2003.02.001 |
| [16] | Schnitzer S A. A mechanistic explanation for global patterns of liana abundance and distribution. The American Naturalist, 2005, 166(2): 262–276. DOI:10.1086/431250 |
| [17] | Tilman D. Resource Competition and Community Structure. Princeton:Princeton University Press, 1982: 1–296. |
| [18] | Hubbell S P. The Unified Neutral Theory of Biodiversity and Biogeography. Princeton:Princeton University Press, 2001: 340–348. |
| [19] | Vamosi S M, Heard S B, Vamosi J C, Webb C O. Emerging patterns in the comparative analysis of phylogenetic community structure. Molecular Ecology, 2009, 18(4): 572–592. DOI:10.1111/mec.2009.18.issue-4 |
| [20] | Webb C O, Ackerly D D, McPeek M A, Donoghue M J. Phylogenies and community ecology. Annual Review of Ecology and Systematics, 2003, 33: 475–505. |
| [21] | Webb C O, Donoghue M J. Phylomatic:tree assembly for applied phylogenetics. Molecular Ecology Notes, 2005, 5(1): 181–183. DOI:10.1111/men.2005.5.issue-1 |
| [22] | Kembel S W, Hubbell S P. The phylogenetic structure of a neotropical forest tree community. Ecology, 2006, 87(sp7): S86–S99. DOI:10.1890/0012-9658(2006)87[86:TPSOAN]2.0.CO;2 |
| [23] | Letcher S G. Phylogenetic structure of angiosperm communities during tropical forest succession. Proceedings of the Royal Society B:Biological Sciences, 2010, 277(1678): 97–104. DOI:10.1098/rspb.2009.0865 |
| [24] | Kraft N J B, Ackerly D D. Functional trait and phylogenetic tests of community assembly across spatial scales in an Amazonian forest. Ecological Monographs, 2010, 80(3): 401–422. DOI:10.1890/09-1672.1 |
| [25] | 王献溥, 郭柯, 温远光. 《广西植被志要》(上、下册). 生命世界, 2015(5): 2–2. |
| [26] | 曾文豪, 曹坤芳, 唐一思, 郑维艳, 石慰. 广西地区喀斯特与非喀斯特山地森林树木物种多样性及系统发育结构比较. 生态学报, 2018, 38(24): 8707–8716. |
| [27] | 吴征镒, 周浙昆, 李德铢, 彭华, 孙航. 世界种子植物科的分布区类型系统. 植物分类与资源学报, 2003, 25(3): 245–257. |
| [28] | 吴征镒, 路安民, 汤彦承, 陈之端, 李德铢. 中国被子植物科属综论. 北京: 科学出版社, 2003. |
| [29] | Gleason H A. The nonconcept of species diversity:A critique and alternative parameters. Ecology, 1971, 52(4): 577–586. DOI:10.2307/1934145 |
| [30] | THE ANGIOSPERM PHYLOGENY GROUP. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants:APG Ⅲ. Botanical Journal of the Linnean Society, 2009, 161(2): 105–121. DOI:10.1111/(ISSN)1095-8339 |
| [31] | Swenson N G, Enquist B J, Thompson J, Zimmerman J K. The influence of spatial and size scale on phylogenetic relatedness in tropical forest communities. Ecology, 2007, 88(7): 1770–1780. DOI:10.1890/06-1499.1 |
| [32] | 胡亮. 东亚温带藤本植物多样性及其格局. 生物多样性, 2011, 19(5): 567–573. |
| [33] | 肖兴艳, 刘方, 姚斌, 舒英格. 中国西南喀斯特土壤水分研究进展. 云南农业大学学报, 2015, 30(6): 958–964. |
| [34] | Yang S Z, Fan H, Li K W, Ko T Y. How the diversity, abundance, size and climbing mechanisms of woody lianas are related to biotic and abiotic factors in a subtropical secondary forest, Taiwan. Folia Geobotanica, 2018, 53(1): 77–88. DOI:10.1007/s12224-017-9306-z |