 2018, Vol. 38
2018, Vol. 38文章信息
- 白耀宇, 庞帅, 殷禄燕, 宋艾妮, 祝增荣.
- BAI Yaoyu, PANG Shuai, YIN Luyan, SONG Aini, ZHU Zengrong.
- 冬水田典型生境类型节肢动物群落多样性及生物量特征
- Arthropod species diversity and biomass of fallow waterlogged paddy fields in Chongqing, China
- 生态学报. 2018, 38(23): 8630-8651
- Acta Ecologica Sinica. 2018, 38(23): 8630-8651
- http://dx.doi.org/10.5846/stxb201801240187
-
文章历史
- 收稿日期: 2018-01-24
- 网络出版日期: 2018-09-29





2. 水稻生物学国家重点实验室, 农业部作物病虫分子生物学重点开放实验室, 浙江大学昆虫科学研究所, 杭州 310058
2. State Key Laboratory of Rice Biology, Ministry of Agriculture Key Laboratory of Molecular Biology of Crop Pathogens and Insects, Institute of Insect Sciences, Zhejiang University, Hangzhou 310058, China
在我国南方稻区, 冬水田是水稻长期栽培过程中形成的一种特殊稻田, 既是一种耕作制度, 又是一种大规模蓄水工程[1];特别对于缺乏水利工程及配套灌溉条件的丘陵山区, 具有重要的生态作用, 其地位和价值更是随着近年来频发的高温干旱, 受到了社会各界的重视[2]。目前, 冬水田广布于四川、重庆等西南丘陵山区[1, 3];仅重庆市面积就达l10万hm2, 占其水稻栽种面积的59.0%以上[4]。
由于冬水田在休耕期残留有大量的水稻植株秸秆(稻桩), 缺乏水稻主要害虫, 且植食性昆虫种类和数量均极度偏少, 导致稻田中捕食者的主要猎物以稻秆残体为食的腐食类弹尾虫等为主, 并形成了与水稻生育期完全不同的生存和觅食策略[5]。同时, 冬水田还田稻秆的难腐解以及由此导致的耕作困难, 特别是稻秆中有机养分如何能被快速分解也是一项重要且现实问题。而以弹尾纲/目(Collembola)为代表的腐食者(分解者)在植株残体撕碎、转化和分解过程中, 其种类组成和数量在很大程度上影响农田生态系统的物质循环[6-7]。此外, 在稻田系统节肢动物随着农事活动存在着群落重建的过程[8], 且稻田耕作和管理措施也对节肢动物群落及其功能团重建有不同程度影响[9-10];而非稻区生境的异质性对稻田节肢动物群落多样性和丰富度等的影响较大[11-15]。因此, 研究冬水田系统中节肢动物群落的多样性和功能特征及其影响因素将对翌年水稻生育期稻田建立积极有效的害虫生物防治体系, 以及稻田秸秆中的养分循环等都具有重要的理论参考价值。但目前这方面的研究还很欠缺。尽管有一些越冬期冬闲田节肢动物群落[16-18]或捕食者蜘蛛[19-20]和腐食者弹尾虫[21-22]调查的报道, 然而涉及休耕期不同冬水田节肢动物群落功能团多样性及其生物量与水稻秸秆、耕作措施及生境因素等相关性分析的研究还鲜有报道。
在重庆等西南山区, 冬水田的种植模式90%以上以年种一季中稻或再生稻为主;水稻从9月收割后蓄水保田休耕至翌年5月移栽的时间长达6—8个月[1, 23]。因此, 在这些单季稻区休耕季节, 冬水田中节肢动物群落多样性特征以及与稻田秸秆、耕作和管理措施等的关系如何, 都不得而知。基于此, 本研究在前期调查基础上, 选择了3个有代表性的重庆丘陵山地冬水田, 调查了休耕期该系统中节肢动物群落的多样性及生物量特征, 分析了稻田秸秆和耕作措施等与这些节肢动物发生的相关性, 以期为冬水田在可持续生态农业理论体系的建立等方面提供可参考的理论依据。
1 研究区概况璧山区(106°02′—106°20′E, 29°17′—29°53′N)位于重庆市主城以西, 处于中亚热带湿润季风气候区, 气候温暖湿润, 雨量充沛。年均气温17.9℃, 极端最高温和最低温分别为41.1℃和-3.0℃;年均降雨量1047 mm, 日照时数1296 h, 无霜期337 d。
八塘镇和河边镇属于璧山区的农业大镇, 分别位于璧山区的最北部和西北部, 农作物的种植以蔬菜、水稻和玉米等为主, 正在积极建设现代都市农业示范基地;其冬水田生境类型属于重庆地区典型的丘陵沟壑稻田(沟壑地带种植水稻, 周边丘陵坡地为蔬菜、退耕还林地或荒地)。该冬水田稻区的种植模式90%以上以年种一季中稻或再生稻为主[23]。研究表明, 环境的异质性能导致稻田节肢动物群落多样性的差异[11, 24], 因此, 在前期调查基础上本研究选取了上述2个镇3类冬水田生境(均为免耕稻田)作为调查的代表样点。另外, 丘陵沟壑冬水田往往处在一狭长不平坦的沟谷地带, 尽管不同田块间可通过加固田埂来保证稻田蓄水量, 但不同位置田块的地势高度落差会造成较长不落雨期间蓄水量的变化, 如地势高的上游田块蓄水量较少, 而地势低的下游田块积水较多。同时, 各个样点内的不同田块往往属于不同农户, 这些田块耕作和管理制度的不同亦会造成田块间的小生境差异。基于此, 本研究在每个调查点地势由高到低的上游、中游和下游分别设置了一个田块作为小区, 试图进一步深入分析各样点内部小生境条件及耕作和管理制度不同对节肢动物群落多样性特征的影响。在当地政府技术人员指导下, 这些稻田在水稻生育期的水肥管理和病虫害的防治措施方面基本相似。3类生境稻田各田块收割前的水稻品种基本一致, 主要为川香优和隆优近似系列, 收割期均为2016年8月下旬。其中样点1(106°19′43″E, 29°50′23″N)和样点2(106°18′5″E, 29°50′31″N)分别位于八塘镇的石苗村和三元村;样点3(106°10′22″E, 29°40′7″N)位于河边镇的浸口村, 其基本情况见图 1和表 1。

|
| 图 1 重庆璧山区不同研究样点冬水田分布及生境图 Fig. 1 Distribution and habitats of FWPF plots of different sampling sites in investigation area of Bishan County, Chongqing FWPF:冬水田Fallow waterlogged paddy fields |
| 样点 Sites |
农田生境类型 Farmland habitat types |
小区分类 Classification of plots |
冬水田信息The information of fallow waterlogged paddy fields (FWPF) | |||||
| 种植模式 Planting patterns |
收割方式 Harvesting methods |
蓄水量评价 Evaluation of water quantity |
稻桩密度 Density of rice stubs/m2 |
稻桩高度 Height of rice stubs/cm |
稻桩生物量 Biomass of rice straws/(g/m2) |
|||
| 样点1 First sampling site |
水稻、蔬菜和 荒地耕作区 |
RVW1 | M | HM | M | 24.98±3.52a | 45.23±2.82a | 0.29±0.06a |
| RVW2 | M | HM | AW | |||||
| RVW3 | M | HM | MW | |||||
| 样点2 Second sampling site |
水稻和蔬菜 耕作区 |
RV1 | MR | HH | MW | 14.98±5.73ab | 35.90±1.54a | 0.15±0.02ab |
| RV2 | MR | HH | MW | |||||
| RV3 | M | HM | MW | |||||
| 样点3 Third sampling site |
水稻和林地 耕作区 |
RF1 | MR | HH | MW | 8.23±1.14b | 36.3±3.54a | 0.13±0.05b |
| RF2 | MR | HH | MW | |||||
| RF3 | MR | HH | MW | |||||
| M:一季中稻种植模式, The planting patterns of medium hybrid rice;MR:一季中稻+再生稻种植模式, The planting patterns of medium hybrid rice and ratoon rice;HM:机械化收割, Harvested by machine;HH:人工收割, Harvested by hand;M:稻田水少但土壤湿润, The soil is reasonably moist in paddy plots;AW:稻田有水但未完全淹没地表, There is a small amount of water in paddy plots;MW:稻田蓄水量多且完全淹没土表, There is much water in paddy plots; 同一列中具不同字母者表示差异显著(P < 0.05) | ||||||||
试验于2017年2月上旬至4月中旬开展。在参考俞晓平等[25]和施波等[26]陷阱法原理基础上, 设置陷阱进行冬水田节肢动物调查(图 2)。在稻田中将玻璃瓶(口径9.0 cm, 底径11 cm, 深24.0 cm, 容积约1050 mL)放入事先挖好坑的土壤中, 保持陷阱瓶口与土表或水面在一个水平面(图 2);然后在杯中倒入少许洗衣粉水液, 即制成捕捉土表或水面以上节肢动物的陷阱。3类冬水田生境均设置了各3个面积为0.2 hm2田块作为研究样地;为了进一步深入分析各个样点不同田块间在蓄水量等生境条件以及耕作和管理制度方面的差异可能导致的影响, 分别在各样点地势由高到低的上游、中游和下游各设置了一个田块作为小区, 每个样点3个, 总共9个(表 1);每个小区均匀放置3—4个陷阱, 陷阱间的距离约为15 m。每3 d取样1次(如遇雨天顺延), 共计18次。

|
| 图 2 重庆璧山区冬水田生境陷阱法设置图 Fig. 2 Pitfall traps in FWPF plots of Bishan County, Chongqing |
由于受降雨的影响或人为/动物的破坏, 各个样点放置的有效回收陷阱(玻璃瓶)总共524个, 其中, 样点1为245个, 样点2和样点3分别为161个和118个。每次调查时用同样规格的玻璃瓶将原陷阱中的玻璃瓶替换, 然后将替换下来的玻璃瓶连同采集到的节肢动物水溶液一起带回实验室。在实验室将其中的节肢动物挑出并保存在75%酒精中进行鉴定和统计数量。蜘蛛鉴定到种, 其他昆虫类群受分类限制均鉴定到科。
2.2 生物量测定冬水田水稻秸秆生物量:在3个样点的每个调查小区选取代表性的样方(4 m×4 m), 将样方内的水稻植株残体齐地面刈割, 除去粘附的土壤和砾石等杂质后放置在室内干燥环境下阴干, 然后称重。此外, 在3个样点每个小区选取的代表性样方秸秆刈割前, 分别测量样方内稻桩的数量和秸秆高度。
稻田节肢动物生物量:将鉴定和统计数量后的所有节肢动物放置在65℃烘箱(型号DGG-9140A, 上海齐欣科学仪器有限公司)中干燥, 大型节肢动物烘干时间12 h, 中小型为6 h;然后将所有节肢动物称重。对于大型节肢动物直接称重记录其生物量(g);但受电子天平(型号FA2004, 上海舜宇恒科学仪器有限公司)精度限制(d值0.1 mg, e值0.1 mg)不能直接称重的中小型种类或类群, 先通过该类群单位数量的生物量推算出单头个体生物量, 然后根据该类群样本个体数计算总的生物量[27]。
2.3 数据处理与分析根据Berger-Parker的优势度指数[28]的计算结果对物种的优势等级进行划分;将其划分为5个等级:D≥0.1时为优势种;当0.05≤D < 0.1时为丰盛种;当0.01≤D < 0.05时为常见种;当0.001≤D < 0.01时为偶见种, 当D < 0.001时为稀少或罕见种;其计算公式为:
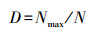
|
式中, Nmax为某类群的个体数;N为群落全部类群的个体数。
冬水稻田节肢动物群落内多样性指数主要采用Simpson优势集中性指数、Shannon-Wiener多样性指数、Pielou均匀度指数和Margalef丰富度指数来进行计算;计算公式分别如下:
Simpson优势集中性指数(C):
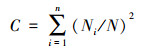
|
Shannon-Wiener多样性指数(H′):
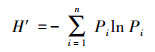
|
Pielou均匀度指数(J):
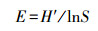
|
Margalef丰富度指数(D):
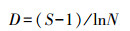
|
式中, Ni为第i类群的个体数;N为群落中所有类群的个体数;Pi=Ni/N, S为类群数。
冬水稻田节肢动物群落组成比较分析主要采用Gower相似性系数来进行计算;该系数主要反映两类生境稻田节肢动物类群上的相似程度;其计算公式为:
Gower系数(Gjk):
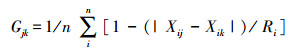
|
式中, 其中Xij和Xik分别为群落j和群落k中类群i的个体数, Ri是所有比较群落中第i个类群最大值和最小值的差;n为所有比较群落的总类群数。Gjk最大值为1, 最小值为0。Gjk值越低, 其相似程度越高。
节肢动物群落及其各功能团密度和生物量数据均为采集的节肢动物个体数换算成的平均密度(个/m2)和平均重量(g/m2)。数据分析前, 对节肢动物密度数据进行Lg (x+1)对数转换;对节肢动物生物量进行Sqrt (x)平方根转换。对服从正态分布和方差齐性的节肢动物群落参数进行单因素方差分析(One-way ANOVA)。而对于不服从正态分布的数据, 采用非参数检验中Kruskal Wallis单因素ANOVA秩和检验(H)进行总体分析, 若有统计学意义(P < 0.05), 进一步用该方法中的两两成对比较法分析。在环境因素、节肢动物及其各功能团特征参数符合正态分布基础上, 相关性分析采用Pearson法两两比较分析;若不符合正态分布, 则采用Spearman法两两比较分析。在对节肢动物密度和生物量、捕食者与猎物在密度和生物量上的相关性分析基础上, 建立了回归方程并拟合了方程曲线。
应用SPSS软件中的聚类分析程序对3个样点9个小区在节肢动物群落相似性方面进行了系统聚类法分组比较。该聚类法在方法上采用平方Euclidean距离法测量, 每两样本间用组间连接法连结。
主成分分析(Principal component analysis, PCA):应用CANOCO软件在对不同冬水稻田环境因子及节肢动物群落参数分析的基础上, 根据这些影响因素的PCA得分对不同冬水田样点进行排序分析。
采集地生境图的获得:在ArcGIS软件系统中, 先将样点坐标经过Albers投影添加到地形图中, 同时结合Google Map遥感数据和现场照片相结合进行制图。
以上分析和作图采用Microsoft Excel 2007、SPSS 24.0、CANOCO 5以及ArcGIS 9.2软件完成。
3 结果与分析 3.1 冬水田节肢动物群落组成及优势类群(种)通过调查共捕获冬水田节肢动物108367只, 隶属于3纲119科(种)。通过单位面积(m2)转换, 这些节肢动物在3类冬水田的密度和生物量统计结果见表 2。由表 2可知, 冬水田节肢动物群落按功能团可分为捕食者、腐食者、植食者、寄生者和杂食者。由密度计算的相对多度结果比较, 3个样点的均值为腐食者>捕食者>植食者>杂食者>寄生者, 腐食者相对多度超过了97%, 是占绝对主导位置的功能群;腐食者的相对多度以RF(样点3)的99.72%为最高, 其后依次是RV(样点2)和RVW(样点1)的98.41%和93.17%;捕食者和植食者的相对多度均为RVW最高, 分别为3.21%和3.22%, 其后依次为RV和RF, 分别为0.93%和0.59%, 0.17%和0.07%;寄生者和杂食者的在3个样点均较小, 为0.01%—0.28%。由生物量计算的各功能团生物量占节肢动物群落生物量的百分比结果比较, 3个样点的均值为捕食者>腐食者>植食者>寄生者>杂食者;捕食者和腐食者的相对生物量分别为46.64%和39.64%, 平均总和达到了86.29%, 均为主要功能群;捕食者以RF的53.67%为最高, 其后依次是RV和RVW的52.09%和34.16%;腐食者以RF的59.92%为最高, 然后依次为RV和RVW的39.08%和19.93%;植食者以RVW的25.75%为最高, 其次分别为RV和RF的7.98%和5.65%;寄生者和杂食者在3个样点均较小, 为0.11%—0.46%。3类冬水田节肢动物类群(种)数以RVW的97个为最高, 而RV和RF的接近, 分别为75个和69个。
| 序号 Sequence number |
类群或种类 Groups and species |
样点1/Site 1(RVW) | 样点2/Site 2(RV) | 样点3/Site 3(RF) | 平均值/Mean | ||||||||||||||||
| /(个/m2) | /% | /(g/m2) | /% | /(个/m2) | /% | /(g/m2) | /% | /(个/m2) | /% | /(g/m2) | /% | /(个/m2) | /% | /(g/m2) | /% | ||||||
| 功能团Functional groups:捕食者Predator | |||||||||||||||||||||
| 蛛形纲Arachnida | |||||||||||||||||||||
| 圆蛛科Araneidae | |||||||||||||||||||||
| 1 | 角园蛛Araneus cornutus | 2812.5 | 0.16 | 3.81 | 0.75 | 781.3 | 0.03 | 0.83 | 0.34 | 625.0 | 0.01 | 0.40 | 0.11 | 1406.3 | 0.06 | 1.68 | 0.40 | ||||
| 2 | 悦目金蛛Argiope amoena | 468.8 | 0.03 | 0.48 | 0.10 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.01 | 0.16 | 0.03 | ||||
| 3 | 横纹金蛛Argiope bruennichi | 1093.8 | 0.06 | 1.48 | 0.29 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 364.6 | 0.02 | 0.49 | 0.10 | ||||
| 4 | 黄金肥蛛Larinia argiopiformis | 156.3 | 0.01 | 0.16 | 0.03 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 52.1 | 0.00 | 0.05 | 0.01 | ||||
| 5 | 黄褐新园蛛Neoscona doenitzi | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.01 | 0.14 | 0.06 | 0.0 | 0.00 | 0.00 | 0.00 | 52.1 | 0.00 | 0.05 | 0.02 | ||||
| 6 | 嗜水新园蛛Neoscona nautica | 156.3 | 0.01 | 0.03 | 0.01 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 52.1 | 0.00 | 0.01 | 0.00 | ||||
| 7 | 棒络新妇Nephila clavata | 312.5 | 0.02 | 0.34 | 0.07 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 104.2 | 0.01 | 0.11 | 0.02 | ||||
| 8 | 四点亮腹蛛Singa pygmaea | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.00 | 0.14 | 0.04 | 52.1 | 0.00 | 0.05 | 0.01 | ||||
| 管巢蛛科Clubionidae | |||||||||||||||||||||
| 9 | 棕管巢蛛Clubiona japonicola | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.01 | 0.17 | 0.07 | 312.5 | 0.00 | 0.18 | 0.05 | 156.3 | 0.00 | 0.12 | 0.04 | ||||
| 10 | 四斑锯螯蛛Dyschiriognatha quabrimaculata | 156.3 | 0.01 | 4.59 | 0.91 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 52.1 | 0.00 | 1.53 | 0.30 | ||||
| 皿蛛科Linyphiidae | |||||||||||||||||||||
| 11 | 隆背微蛛Erigone prominens | 2187.5 | 0.12 | 0.18 | 0.04 | 1718.8 | 0.06 | 0.17 | 0.07 | 625.0 | 0.01 | 0.16 | 0.04 | 1510.4 | 0.06 | 0.17 | 0.05 | ||||
| 12 | 齿螯额角蛛Gnathonarium dentatum | 156.3 | 0.01 | 0.01 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 52.1 | 0.00 | 0.00 | 0.00 | ||||
| 13 | 驼背额角蛛Gnathonarium gibberum | 468.8 | 0.03 | 0.06 | 0.01 | 156.3 | 0.01 | 0.06 | 0.02 | 0.0 | 0.00 | 0.00 | 0.00 | 208.3 | 0.01 | 0.04 | 0.01 | ||||
| 14 | 草间小黑蛛Hylyphantes graminicola | 2187.5 | 0.12 | 0.12 | 0.02 | 625.0 | 0.02 | 0.03 | 0.01 | 625.0 | 0.01 | 0.04 | 0.01 | 1145.8 | 0.05 | 0.06 | 0.01 | ||||
| 15 | 黑侏儒蛛Meioneta nigra | 156.3 | 0.01 | 0.01 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 52.1 | 0.00 | 0.00 | 0.00 | ||||
| 16 | 食虫沟瘤蛛Ummeliata insecticeps | 3281.3 | 0.18 | 1.04 | 0.21 | 312.5 | 0.01 | 0.13 | 0.05 | 156.3 | 0.00 | 0.01 | 0.00 | 1250.0 | 0.07 | 0.39 | 0.09 | ||||
| 17 | 东京沟瘤蛛Ummeliata tokyoensis | 625.0 | 0.03 | 0.03 | 0.01 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 208.3 | 0.01 | 0.01 | 0.00 | ||||
| 狼蛛科Lycosidae | |||||||||||||||||||||
| 18 | 星豹蛛Pardosa astrigera | 1250.0 | 0.07 | 9.68 | 1.91 | 937.5 | 0.03 | 7.80 | 3.20 | 937.5 | 0.01 | 11.04 | 2.89 | 1041.7 | 0.04 | 9.50 | 2.67 | ||||
| 19 | 沟渠豹蛛Pardosa laura | 4375.0 | 0.24 | 60.61 | 11.96 | 1406.3 | 0.05 | 12.39 | 5.09 | 312.5 | 0.00 | 1.48 | 0.39 | 2031.3 | 0.10 | 24.83 | 5.81 | ||||
| 20 | 拟环纹豹蛛Pardosa pseudoannulata | 781.3 | 0.04 | 10.25 | 2.02 | 312.5 | 0.01 | 3.81 | 1.56 | 156.3 | 0.00 | 1.44 | 0.38 | 416.7 | 0.02 | 5.17 | 1.32 | ||||
| 21 | 丁纹豹蛛Pardosa tinsignita | 2343.8 | 0.13 | 18.36 | 3.62 | 625.0 | 0.02 | 9.06 | 3.72 | 156.3 | 0.00 | 1.89 | 0.49 | 1041.7 | 0.05 | 9.77 | 2.61 | ||||
| 22 | 真水狼蛛Pirala piraticus | 1875.0 | 0.10 | 23.48 | 4.64 | 781.3 | 0.03 | 8.03 | 3.30 | 2031.3 | 0.02 | 16.39 | 4.29 | 1562.5 | 0.05 | 15.97 | 4.07 | ||||
| 23 | 类水狼蛛Pirata piratoides | 4531.3 | 0.25 | 34.75 | 6.86 | 4062.5 | 0.15 | 31.89 | 13.09 | 3593.8 | 0.03 | 26.55 | 6.95 | 4062.5 | 0.14 | 31.06 | 8.96 | ||||
| 24 | 拟水狼蛛Pirata subpiraticus | 4531.3 | 0.25 | 50.13 | 9.89 | 3437.5 | 0.12 | 41.98 | 17.23 | 2812.5 | 0.02 | 35.61 | 9.32 | 3593.8 | 0.13 | 42.57 | 12.15 | ||||
| 25 | 奇异獾蛛Trochosa ruricola | 937.5 | 0.05 | 12.84 | 2.53 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 312.5 | 0.02 | 4.28 | 0.84 | ||||
| 球腹蛛科Theridiidae | |||||||||||||||||||||
| 26 | 四突瘤腹蛛Chrosiothes sudabides | 156.3 | 0.01 | 0.16 | 0.03 | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.00 | 0.01 | 0.00 | 104.2 | 0.00 | 0.06 | 0.01 | ||||
| 27 | 八斑鞘腹蛛Coleosoma octomaculatum | 156.3 | 0.01 | 0.01 | 0.00 | 156.3 | 0.01 | 0.01 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 104.2 | 0.00 | 0.01 | 0.00 | ||||
| 蟹蛛科Thomisidae | |||||||||||||||||||||
| 28 | 三突花蛛Misumenops tricuspidatus | 1406.3 | 0.08 | 0.64 | 0.13 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 468.8 | 0.03 | 0.21 | 0.04 | ||||
| 29 | 条纹蝇虎Plexippus setipet | 156.3 | 0.01 | 0.84 | 0.17 | 156.3 | 0.01 | 0.15 | 0.06 | 0.0 | 0.00 | 0.00 | 0.00 | 104.2 | 0.00 | 0.33 | 0.08 | ||||
| 30 | 斜纹花蟹蛛Xysticus saganus | 156.3 | 0.01 | 0.02 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 52.1 | 0.00 | 0.01 | 0.00 | ||||
| 肖蛸科Tetragnathidae | |||||||||||||||||||||
| 31 | 鳞纹肖蛸Tetragnatha squamata | 312.5 | 0.02 | 4.59 | 0.91 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 104.2 | 0.01 | 1.53 | 0.30 | ||||
| 跳蛛科Salticidae | |||||||||||||||||||||
| 32 | 微菱头蛛Bianor aenescens | 156.3 | 0.01 | 2.34 | 0.46 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 52.1 | 0.00 | 0.78 | 0.15 | ||||
| 33 | 纵条蝇狮Marpissa magister | 156.3 | 0.01 | 0.25 | 0.05 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 52.1 | 0.00 | 0.08 | 0.02 | ||||
| 昆虫纲Insecta | |||||||||||||||||||||
| 鞘翅目Coleoptera | |||||||||||||||||||||
| 34 | 步甲科Carabidae | 468.8 | 0.03 | 0.42 | 0.08 | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.00 | 0.12 | 0.03 | 208.3 | 0.01 | 0.18 | 0.04 | ||||
| 35 | 瓢甲科Coccinellidae | 312.5 | 0.02 | 0.69 | 0.14 | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.00 | 0.12 | 0.03 | 156.3 | 0.01 | 0.27 | 0.06 | ||||
| 36 | 鼓甲科Gyrinidae | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.01 | 0.02 | 0.01 | 0.0 | 0.00 | 0.00 | 0.00 | 52.1 | 0.00 | 0.01 | 0.00 | ||||
| 37 | 隐翅甲科Staphylinidae | 7656.3 | 0.43 | 8.45 | 1.67 | 3125.0 | 0.11 | 3.72 | 1.53 | 1875.0 | 0.02 | 2.00 | 0.52 | 4218.8 | 0.19 | 4.72 | 1.24 | ||||
| 双翅目Diptera | |||||||||||||||||||||
| 38 | 食虫虻科Asilidae | 937.5 | 0.05 | 0.54 | 0.11 | 312.5 | 0.01 | 0.28 | 0.12 | 312.5 | 0.00 | 0.44 | 0.11 | 520.8 | 0.02 | 0.42 | 0.11 | ||||
| 39 | 长足虻科Dolichopodidae | 625.0 | 0.03 | 0.39 | 0.08 | 1875.0 | 0.07 | 1.04 | 0.43 | 156.3 | 0.00 | 0.09 | 0.02 | 885.4 | 0.03 | 0.50 | 0.18 | ||||
| 40 | 沼蝇科Sciomyzidae | 1875.0 | 0.10 | 7.88 | 1.55 | 625.0 | 0.02 | 2.63 | 1.08 | 1250.0 | 0.01 | 5.06 | 1.32 | 1250.0 | 0.05 | 5.19 | 1.32 | ||||
| 41 | 食蚜蝇科Syrphidae | 156.3 | 0.01 | 0.07 | 0.01 | 156.3 | 0.01 | 0.07 | 0.03 | 0.0 | 0.00 | 0.00 | 0.00 | 104.2 | 0.00 | 0.05 | 0.01 | ||||
| 42 | 虻科Tabanidae | 3593.8 | 0.20 | 3.74 | 0.74 | 2812.5 | 0.10 | 1.63 | 0.67 | 1093.8 | 0.01 | 0.62 | 0.16 | 2500.0 | 0.10 | 2.00 | 0.52 | ||||
| 43 | 剑虻科Therevidae | 312.5 | 0.02 | 0.23 | 0.05 | 312.5 | 0.01 | 0.18 | 0.07 | 0.0 | 0.00 | 0.00 | 0.00 | 208.3 | 0.01 | 0.14 | 0.04 | ||||
| 半翅目Hemiptera | |||||||||||||||||||||
| 44 | 田鳖科Belostmatidae | 156.3 | 0.01 | 1.45 | 0.29 | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.00 | 11.38 | 2.98 | 104.2 | 0.00 | 4.28 | 1.09 | ||||
| 45 | 划蝽科Corixidae | 3437.5 | 0.19 | 3.33 | 0.66 | 781.3 | 0.03 | 0.60 | 0.25 | 1250.0 | 0.01 | 0.94 | 0.25 | 1822.9 | 0.08 | 1.62 | 0.38 | ||||
| 46 | 水黾科Gerridae | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.00 | 0.03 | 0.01 | 52.1 | 0.00 | 0.01 | 0.00 | ||||
| 47 | 猎蝽科Reduviidae | 312.5 | 0.02 | 2.28 | 0.45 | 0.0 | 0.00 | 0.00 | 0.00 | 1562.5 | 0.01 | 12.02 | 3.14 | 625.0 | 0.01 | 4.77 | 1.20 | ||||
| 48 | 宽黾蝽科Veliidae | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.01 | 0.08 | 0.03 | 0.0 | 0.00 | 0.00 | 0.00 | 52.1 | 0.00 | 0.03 | 0.01 | ||||
| 汇总 | 57500.0 | 3.21 | 271.94 | 53.67 | 26093.8 | 0.93 | 126.91 | 52.09 | 21093.8 | 0.17 | 130.54 | 34.16 | 34895.8 | 1.44 | 176.46 | 46.64 | |||||
| 功能团Functional groups:腐食者Detritivore | |||||||||||||||||||||
| 弹尾纲Collembola | |||||||||||||||||||||
| 49 | 长角 科Entomobryidae 科Entomobryidae |
626406.3 | 35.01 | 34.89 | 6.89 | 865781.3 | 31.02 | 50.86 | 20.88 | 526562.5 | 4.27 | 28.24 | 7.39 | 672916.7 | 23.43 | 38.00 | 11.72 | ||||
| 50 | 球角科Hypogastruridae |
468.8 | 0.03 | 0.01 | 0.00 | 312.5 | 0.01 | 0.02 | 0.01 | 781.3 | 0.01 | 0.02 | 0.00 | 520.8 | 0.01 | 0.01 | 0.00 | ||||
| 51 | 等节科Isotomidae |
468.8 | 0.03 | 0.03 | 0.01 | 1562.5 | 0.06 | 0.13 | 0.05 | 0.0 | 0.00 | 0.00 | 0.00 | 677.1 | 0.03 | 0.05 | 0.02 | ||||
| 52 | 圆科Sminthuridae |
749062.5 | 41.87 | 11.60 | 2.29 | 1702188.0 | 60.98 | 24.77 | 10.17 | 11597344.0 | 93.97 | 167.01 | 43.70 | 4682865.0 | 65.61 | 67.79 | 18.72 | ||||
| 昆虫纲Insecta | |||||||||||||||||||||
| 鞘翅目Coleoptera | |||||||||||||||||||||
| 53 | 坚甲科Colydioidae | 2343.8 | 0.13 | 7.81 | 1.54 | 2031.3 | 0.07 | 1.77 | 0.73 | 937.5 | 0.01 | 1.61 | 0.42 | 1770.8 | 0.07 | 3.73 | 0.90 | ||||
| 54 | 皮蠹科Dermestidae | 156.3 | 0.01 | 0.14 | 0.03 | 0.0 | 0.00 | 0.00 | 0.00 | 312.5 | 0.00 | 2.89 | 0.76 | 156.3 | 0.00 | 1.01 | 0.26 | ||||
| 55 | 牙甲科Hydrophilidae | 156.3 | 0.01 | 1.47 | 0.29 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 52.1 | 0.00 | 0.49 | 0.10 | ||||
| 56 | 筒蠹科Lymexylidae | 468.8 | 0.03 | 5.13 | 1.01 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.01 | 1.71 | 0.34 | ||||
| 57 | 蚁甲科Pselaphidae | 312.5 | 0.02 | 0.12 | 0.02 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 104.2 | 0.01 | 0.04 | 0.01 | ||||
| 58 | 埋葬甲科Silphidae | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 312.5 | 0.00 | 2.63 | 0.69 | 104.2 | 0.00 | 0.88 | 0.23 | ||||
| 59 | 拟步甲科Tenebrionidae | 312.5 | 0.02 | 0.31 | 0.06 | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.00 | 0.20 | 0.05 | 156.3 | 0.01 | 0.17 | 0.04 | ||||
| 双翅目Diptera | |||||||||||||||||||||
| 60 | 花蝇科Anthomyiidae | 2343.8 | 0.13 | 1.35 | 0.27 | 468.8 | 0.02 | 0.33 | 0.14 | 781.3 | 0.01 | 0.50 | 0.13 | 1197.9 | 0.05 | 0.73 | 0.18 | ||||
| 61 | 毛蚊科Bibionidae | 937.5 | 0.05 | 0.51 | 0.10 | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.00 | 0.08 | 0.02 | 364.6 | 0.02 | 0.20 | 0.04 | ||||
| 62 | 网蚊科Blepharoceridae | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 312.5 | 0.00 | 0.03 | 0.01 | 104.2 | 0.00 | 0.01 | 0.00 | ||||
| 63 | 蚊科Culicidae | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 312.5 | 0.00 | 0.01 | 0.00 | 104.2 | 0.00 | 0.00 | 0.00 | ||||
| 64 | 细蚊科Dixidae | 22500.0 | 1.26 | 0.67 | 0.13 | 16406.3 | 0.59 | 0.67 | 0.27 | 4218.8 | 0.03 | 0.18 | 0.05 | 14375.0 | 0.63 | 0.50 | 0.15 | ||||
| 65 | 蕈蚊科Mycetophilidae | 129375.0 | 7.23 | 10.71 | 2.11 | 28281.3 | 1.01 | 2.40 | 0.99 | 40156.3 | 0.33 | 3.29 | 0.86 | 65937.5 | 2.86 | 5.47 | 1.32 | ||||
| 66 | 瘿蚊科Mycetophilidae | 7187.5 | 0.40 | 0.84 | 0.16 | 7812.5 | 0.28 | 0.69 | 0.28 | 2187.5 | 0.02 | 0.17 | 0.04 | 5729.2 | 0.23 | 0.56 | 0.16 | ||||
| 67 | 蛾蚋科Psychodidae | 34218.8 | 1.91 | 3.68 | 0.73 | 37343.8 | 1.34 | 2.17 | 0.89 | 11562.5 | 0.09 | 0.62 | 0.16 | 27708.3 | 1.11 | 2.16 | 0.59 | ||||
| 68 | 蚋科Simuliidae | 40937.5 | 2.29 | 1.91 | 0.38 | 42343.8 | 1.52 | 1.74 | 0.72 | 89531.3 | 0.73 | 4.52 | 1.18 | 57604.2 | 1.51 | 2.73 | 0.76 | ||||
| 69 | 大蚊科Tipulidae | 21250.0 | 1.19 | 16.15 | 3.19 | 10312.5 | 0.37 | 5.09 | 2.09 | 19218.8 | 0.16 | 16.50 | 4.32 | 16927.1 | 0.57 | 12.58 | 3.20 | ||||
| 70 | 寡脉蝇科Asteiidae | 937.5 | 0.05 | 0.60 | 0.12 | 4062.5 | 0.15 | 2.32 | 0.95 | 0.0 | 0.00 | 0.00 | 0.00 | 1666.7 | 0.07 | 0.97 | 0.36 | ||||
| 71 | 丽蝇科Calliphoridae | 312.5 | 0.02 | 0.19 | 0.04 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 104.2 | 0.01 | 0.06 | 0.01 | ||||
| 72 | 蠓科Ceratopogouidae | 312.5 | 0.02 | 0.20 | 0.04 | 156.3 | 0.01 | 0.09 | 0.04 | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.01 | 0.10 | 0.03 | ||||
| 73 | 摇蚊科Chironomidae | 20937.5 | 1.17 | 0.45 | 0.09 | 21562.5 | 0.77 | 0.45 | 0.18 | 4218.8 | 0.03 | 0.09 | 0.02 | 15572.9 | 0.66 | 0.33 | 0.10 | ||||
| 74 | 躅大蚊科Cylindrotomidae | 625.0 | 0.03 | 0.18 | 0.04 | 156.3 | 0.01 | 0.09 | 0.04 | 0.0 | 0.00 | 0.00 | 0.00 | 260.4 | 0.01 | 0.09 | 0.02 | ||||
| 75 | 沼大蚊科Limoniidae | 0.0 | 0.00 | 0.00 | 0.00 | 468.8 | 0.02 | 0.02 | 0.01 | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.01 | 0.01 | 0.00 | ||||
| 76 | 尖翅蝇科Lonchopteridae | 0.0 | 0.00 | 0.00 | 0.00 | 312.5 | 0.01 | 0.09 | 0.04 | 0.0 | 0.00 | 0.00 | 0.00 | 104.2 | 0.00 | 0.03 | 0.01 | ||||
| 77 | 蝇科Muscidae | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.01 | 0.09 | 0.04 | 468.8 | 0.00 | 0.20 | 0.05 | 208.3 | 0.00 | 0.10 | 0.03 | ||||
| 78 | 菌蚊科Mycetophilidae | 0.0 | 0.00 | 0.00 | 0.00 | 312.5 | 0.01 | 0.01 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 104.2 | 0.00 | 0.00 | 0.00 | ||||
| 79 | 粗脉毛蚊科Pachyneuridae | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.01 | 0.17 | 0.07 | 0.0 | 0.00 | 0.00 | 0.00 | 52.1 | 0.00 | 0.06 | 0.02 | ||||
| 80 | 蚤蝇科Phoridae | 781.3 | 0.04 | 0.63 | 0.12 | 156.3 | 0.01 | 0.01 | 0.00 | 156.3 | 0.00 | 0.04 | 0.01 | 364.6 | 0.02 | 0.23 | 0.05 | ||||
| 81 | 毛蠓科Psychodidae | 1406.3 | 0.08 | 0.90 | 0.18 | 1718.8 | 0.06 | 0.86 | 0.35 | 0.0 | 0.00 | 0.00 | 0.00 | 1041.7 | 0.05 | 0.59 | 0.18 | ||||
| 82 | 粪蚊科Scastopsidae | 937.5 | 0.05 | 0.27 | 0.05 | 156.3 | 0.01 | 0.02 | 0.01 | 0.0 | 0.00 | 0.00 | 0.00 | 364.6 | 0.02 | 0.10 | 0.02 | ||||
| 83 | 水虻科Stratiomyidae | 468.8 | 0.03 | 0.14 | 0.03 | 468.8 | 0.02 | 0.28 | 0.11 | 0.0 | 0.00 | 0.00 | 0.00 | 312.5 | 0.01 | 0.14 | 0.05 | ||||
| 蜉蝣目Ephemeroptera | |||||||||||||||||||||
| 84 | 蜉蝣科Ephemeridae | 1093.8 | 0.06 | 0.03 | 0.01 | 1250.0 | 0.04 | 0.04 | 0.02 | 6875.0 | 0.06 | 0.22 | 0.06 | 3072.9 | 0.05 | 0.10 | 0.03 | ||||
| 汇总 | 1666719.0 | 93.17 | 100.92 | 19.93 | 2745938.0 | 98.41 | 95.18 | 39.08 | 12306563.0 | 99.72 | 229.04 | 59.92 | 5573074.0 | 97.08 | 141.73 | 39.65 | |||||
| 功能团Functional groups:植食者Herbivore | |||||||||||||||||||||
| 鞘翅目Coleoptera | |||||||||||||||||||||
| 85 | 吉丁甲科Buprestidae | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.01 | 0.09 | 0.04 | 0.0 | 0.00 | 0.00 | 0.00 | 52.1 | 0.00 | 0.03 | 0.01 | ||||
| 86 | 叶甲科Chrysomelidae | 6718.8 | 0.38 | 81.78 | 16.14 | 781.3 | 0.03 | 7.51 | 3.08 | 1406.3 | 0.01 | 4.92 | 1.29 | 2968.8 | 0.14 | 31.41 | 6.84 | ||||
| 87 | 象甲科Curculionidae | 312.5 | 0.02 | 0.23 | 0.05 | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.00 | 0.11 | 0.03 | 156.3 | 0.01 | 0.11 | 0.02 | ||||
| 88 | 叩甲科Elateridae | 156.3 | 0.01 | 0.17 | 0.03 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 52.1 | 0.00 | 0.06 | 0.01 | ||||
| 89 | 金龟科Scarabaeidae | 781.3 | 0.04 | 3.19 | 0.63 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 260.4 | 0.01 | 1.06 | 0.21 | ||||
| 双翅目Diptera | |||||||||||||||||||||
| 90 | 潜蝇科Agromyzidae | 937.5 | 0.05 | 0.54 | 0.11 | 468.8 | 0.02 | 0.25 | 0.10 | 1250.0 | 0.01 | 0.76 | 0.20 | 885.4 | 0.03 | 0.52 | 0.14 | ||||
| 91 | 秆蝇科Chloropidae | 12968.8 | 0.72 | 11.44 | 2.26 | 5156.3 | 0.18 | 3.18 | 1.31 | 1718.8 | 0.01 | 1.26 | 0.33 | 6614.6 | 0.31 | 5.29 | 1.30 | ||||
| 92 | 水蝇科Ephydridae | 11718.8 | 0.66 | 5.45 | 1.08 | 2187.5 | 0.08 | 1.26 | 0.52 | 1562.5 | 0.01 | 0.73 | 0.19 | 5156.3 | 0.25 | 2.48 | 0.59 | ||||
| 93 | 实蝇科Tephritidae | 2031.3 | 0.11 | 1.18 | 0.23 | 1718.8 | 0.06 | 1.09 | 0.45 | 156.3 | 0.00 | 0.10 | 0.02 | 1302.1 | 0.06 | 0.79 | 0.24 | ||||
| 半翅目Hemiptera | |||||||||||||||||||||
| 94 | 蚜科Aphididae | 9218.8 | 0.52 | 0.37 | 0.07 | 312.5 | 0.01 | 0.01 | 0.01 | 156.3 | 0.00 | 0.01 | 0.00 | 3229.2 | 0.18 | 0.13 | 0.03 | ||||
| 95 | 叶蝉科Cicadellidae | 6875.0 | 0.38 | 2.63 | 0.52 | 2187.5 | 0.08 | 0.34 | 0.14 | 2968.8 | 0.02 | 0.36 | 0.09 | 4010.4 | 0.16 | 1.11 | 0.25 | ||||
| 96 | 缘蝽科Coreidae | 156.3 | 0.01 | 0.10 | 0.02 | 312.5 | 0.01 | 0.81 | 0.33 | 156.3 | 0.00 | 0.10 | 0.03 | 208.3 | 0.01 | 0.34 | 0.13 | ||||
| 97 | 飞虱科Delphacidae | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.01 | 0.02 | 0.01 | 156.3 | 0.00 | 0.02 | 0.00 | 104.2 | 0.00 | 0.01 | 0.00 | ||||
| 98 | 蜡蝉科Fulgoridae | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.01 | 0.12 | 0.05 | 0.00 | 0.00 | 0.00 | 0.00 | 52.1 | 0.00 | 0.04 | 0.02 | ||||
| 99 | 长蝽科Lygaeidae | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.01 | 0.97 | 0.40 | 0.00 | 0.00 | 0.00 | 0.00 | 52.1 | 0.00 | 0.32 | 0.13 | ||||
| 100 | 角蝉科Membracidae | 937.5 | 0.05 | 0.62 | 0.12 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 312.5 | 0.02 | 0.21 | 0.04 | ||||
| 101 | 盲蝽科Miridae | 1562.5 | 0.09 | 0.77 | 0.15 | 468.8 | 0.02 | 0.33 | 0.13 | 1093.8 | 0.01 | 0.63 | 0.17 | 1041.7 | 0.04 | 0.58 | 0.15 | ||||
| 102 | 木虱科Psyllidae | 1093.8 | 0.06 | 0.08 | 0.02 | 312.5 | 0.01 | 0.08 | 0.03 | 312.5 | 0.00 | 0.02 | 0.01 | 572.9 | 0.02 | 0.06 | 0.02 | ||||
| 鳞翅目Lepidoptera | |||||||||||||||||||||
| 103 | 细蛾科Gracilariidae | 0.0 | 0.00 | 0.00 | 0.00 | 625.0 | 0.02 | 1.30 | 0.53 | 312.5 | 0.00 | 10.68 | 2.79 | 312.5 | 0.01 | 3.99 | 1.11 | ||||
| 104 | 弄蝶科Hesperiidae | 0.0 | 0.00 | 0.00 | 0.00 | 312.5 | 0.01 | 2.06 | 0.85 | 0.0 | 0.00 | 0.00 | 0.00 | 104.2 | 0.00 | 0.69 | 0.28 | ||||
| 105 | 夜蛾科Noctuidae | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 468.8 | 0.00 | 1.92 | 0.50 | 156.3 | 0.00 | 0.64 | 0.17 | ||||
| 直翅目Orthoptera | |||||||||||||||||||||
| 106 | 蟋蟀科Gryllidae | 468.8 | 0.03 | 4.02 | 0.79 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.01 | 1.34 | 0.26 | ||||
| 107 | 蝼蛄科Gryllotalpidae | 156.3 | 0.01 | 1.52 | 0.30 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 52.1 | 0.00 | 0.51 | 0.10 | ||||
| 108 | 菱蝗科Tetrigidae | 1406.3 | 0.08 | 16.35 | 3.23 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 468.8 | 0.03 | 5.45 | 1.08 | ||||
| 缨翅目Thysanoptera | |||||||||||||||||||||
| 109 | 蓟马科Thripidae | 0.0 | 0.00 | 0.00 | 0.00 | 468.8 | 0.02 | 0.01 | 0.00 | 468.8 | 0.00 | 0.01 | 0.00 | 312.5 | 0.01 | 0.01 | 0.00 | ||||
| 汇总 | 57500.6 | 3.22 | 130.44 | 25.75 | 15938.0 | 0.59 | 19.43 | 7.98 | 12344.3 | 0.07 | 21.63 | 5.65 | 28594.2 | 1.30 | 57.18 | 13.13 | |||||
| 功能团Functional groups:寄生者Parasite | |||||||||||||||||||||
| 双翅目Diptera | |||||||||||||||||||||
| 110 | 寄蝇科Tachinidae | 156.3 | 0.01 | 0.11 | 0.02 | 781.3 | 0.03 | 0.23 | 0.09 | 0.0 | 0.00 | 0.00 | 0.00 | 312.5 | 0.01 | 0.11 | 0.04 | ||||
| 膜翅目Hymenoptera | |||||||||||||||||||||
| 111 | 茧蜂科Braconidae | 937.5 | 0.05 | 0.78 | 0.15 | 937.5 | 0.03 | 0.62 | 0.26 | 156.3 | 0.00 | 0.12 | 0.03 | 677.1 | 0.03 | 0.51 | 0.15 | ||||
| 112 | 小蜂科Chalalcididae | 468.8 | 0.03 | 0.36 | 0.07 | 0.0 | 0.00 | 0.00 | 0.00 | 312.5 | 0.00 | 0.25 | 0.06 | 260.4 | 0.01 | 0.20 | 0.05 | ||||
| 113 | 瘿蜂科Cynipidae | 156.3 | 0.01 | 0.12 | 0.02 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 52.1 | 0.00 | 0.04 | 0.01 | ||||
| 114 | 环腹瘿蜂科Figitidae | 156.3 | 0.01 | 0.13 | 0.03 | 156.3 | 0.01 | 0.11 | 0.05 | 156.3 | 0.00 | 0.13 | 0.03 | 156.3 | 0.01 | 0.13 | 0.04 | ||||
| 115 | 姬蜂科Ichneumonidae | 312.5 | 0.02 | 0.23 | 0.04 | 0.0 | 0.00 | 0.00 | 0.00 | 0.0 | 0.00 | 0.00 | 0.00 | 104.2 | 0.01 | 0.08 | 0.01 | ||||
| 116 | 窄腹细蜂科Roproniidae | 156.3 | 0.01 | 0.13 | 0.03 | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.00 | 0.11 | 0.03 | 104.2 | 0.00 | 0.08 | 0.02 | ||||
| 汇总 | 2343.8 | 0.13 | 1.86 | 0.37 | 1875.0 | 0.07 | 0.96 | 0.40 | 781.3 | 0.01 | 0.60 | 0.16 | 1666.7 | 0.07 | 1.14 | 0.31 | |||||
| 功能团Functional groups:杂食者Omnivore | |||||||||||||||||||||
| 鞘翅目Coleoptera | |||||||||||||||||||||
| 117 | 出尾甲科Nitidulidae | 781.3 | 0.04 | 1.42 | 0.28 | 1093.8 | 0.04 | 0.96 | 0.40 | 312.5 | 0.00 | 0.26 | 0.07 | 729.2 | 0.03 | 0.88 | 0.25 | ||||
| 膜翅目Hymenoptera | |||||||||||||||||||||
| 118 | 蚁科Formicidae | 0.0 | 0.00 | 0.00 | 0.00 | 156.3 | 0.01 | 0.16 | 0.06 | 156.3 | 0.00 | 0.16 | 0.04 | 104.2 | 0.00 | 0.10 | 0.04 | ||||
| 缨尾目Thysanura | |||||||||||||||||||||
| 119 | 衣鱼科Lepismatidae | 4218.8 | 0.24 | 0.14 | 0.03 | 312.5 | 0.01 | 0.01 | 0.00 | 312.5 | 0.00 | 0.01 | 0.00 | 1614.6 | 0.08 | 0.05 | 0.01 | ||||
| 汇总 | 5000.0 | 0.28 | 1.55 | 0.31 | 1562.5 | 0.06 | 1.13 | 0.46 | 781.3 | 0.01 | 0.42 | 0.11 | 2447.9 | 0.11 | 1.04 | 0.29 | |||||
| 3个样地汇总 | 1789062.5 | 100.00 | 506.67 | 100.00 | 2791406.3 | 100.0 | 243.62 | 100.00 | 12341562.5 | 100.00 | 382.20 | 100.00 | 5640677.1 | 100.00 | 377.50 | 100.00 | |||||
| 总类群数(科和种类) | 97 | 75 | 69 | 119 | |||||||||||||||||
| Berger-Parker优势度指数(D) | 3, 0, 6, 31, 57 | 2, 0, 4, 13, 32 | 1, 0, 1, 4, 63 | ||||||||||||||||||
| Berger-Parker的优势度指数(D)排序依次为:优势类群(种)、丰盛类群(种)、常见类群(种)、偶见类群(种)和罕见或稀少类群(种) | |||||||||||||||||||||
Berger-Parker优势度指数结果指出, 冬水田节肢动物群落中的罕见或稀少种(类群)数量最多, 其后依次是偶见种(类群)、常见种(类群)和优势种(类群);优势类群仅有腐食功能团中的圆
冬水田3个样点的节肢动物平均密度为RF>RV>RVW, RF显著大于RV和RVW(P < 0.05), RVW和RV差异不显著(P>0.05);其中RVW中的RVW1和RVW3显著差异(P < 0.05), RV中的RV1分别与RV2和RV3差异显著(P < 0.05), RF中的RF1和RF3均远大于RF2, 且差异显著(P < 0.05)(图 3)。腐食者在3个样点间的平均密度为RF显著大于RVW和RV(P < 0.05);RV中的RV2和RV3显著大于RV1(P < 0.05);RF中的RF3显著大于RF2(P < 0.05);其他样点及其小区间的差异均不显著(P>0.05)。捕食者在3个样点间的平均密度均差异不显著(P>0.05);RVW1显著大于RVW3(P < 0.05), 其他样点各小区间均差异不显著(P>0.05)。植食者中的RVW显著大于RV和RF(P < 0.05);其他样点及其各小区间的差异均不显著(P>0.05)(图 3)。

|
| 图 3 不同休耕期冬水田调查点节肢动物群落密度(平均值±标准误) Fig. 3 Densities of arthropod community in FWPFs and their plots of three sites (Mean±SE) RVW1:水稻、蔬菜和荒地耕作区小区1, The first plot of mixed areas of rice, vegetables and wasteland;RVW2:水稻、蔬菜和荒地耕作区小区2, The second plot of mixed areas of rice, vegetables and wasteland;RVW3:水稻、蔬菜和荒地耕作区小区3, The third plot of mixed areas of rice, vegetables and wasteland;RV1:水稻和蔬菜耕作区小区1, The first plot of mixed areas of rice and vegetables;RV2:水稻和蔬菜耕作区小区2, The second plot of mixed areas of rice and vegetables;RV3:水稻和蔬菜耕作区小区3, The third plot of mixed areas of rice and vegetables;RF1:水稻和林地耕作区小区1, The first plot of mixed areas of rice and forestland;RF2:水稻和林地耕作区小区2, The second plot of mixed areas of rice and forestland;RF3:水稻和林地耕作区小区3, The third plot of mixed areas of rice and forestland;RVW:水稻、蔬菜和荒地耕作区, The mixed areas of rice, vegetables and wasteland;RV:水稻和蔬菜耕作区, The mixed areas of rice and vegetables;RF:水稻和林地耕作区, The mixed areas of rice and forestland; 同一组图柱上字母不相同者表示在0.05显著水平上的差异显著 |
节肢动物在3个样点的平均生物量为RF显著大于RVW和RV(P < 0.05), RVW和RV差异不显著(P>0.05);3个样点各小区间差异均不显著(P>0.05)(图 4)。腐食者在RF和RV的平均生物量显著大于RVW(P < 0.05);RV中的3个小区间均差异显著(P < 0.05);RF中的RF3显著大于RF1和RF2(P < 0.05);其他样点及其小区间的差异均不显著(P>0.05)。捕食者在3个样点间的平均生物量均差异不显著(P>0.05);RVW中的RVW1和RVW2显著大于RVW3(P < 0.05);其他样点各小区间差异均不显著(P>0.05)。植食者在RVW的平均生物量显著大于RV和RF(P < 0.05);其他样点及其小区间的差异均不显著(P>0.05)(图 4)。

|
| 图 4 三类冬水田调查点节肢动物群落及其功能团生物量(平均值±标准误) Fig. 4 Biomass of arthropod community in FWPFs and their plots of three sites (Mean±SE) |
Shannon-Wiener多样性指数在3个样点间, RVW和RV显著大于RF(P < 0.05), RVW和RV间差异不显著(P>0.05);其中, RVW中的RVW1和RVW2显著大于RVW3(P < 0.05), RV中的RV1显著大于RV2和RV3(P < 0.05), RF中的RF2显著大于RF1和RF3(P < 0.05), 其他样点各小区间差异均不显著(P>0.05)。对于Margalef丰富度指数, RVW显著大于RF(P < 0.05), 其他样点间差异不显著(P>0.05);其中, RVW中的RVW1和RVW2显著大于RVW3(P < 0.05), 其他样点各小区间差异均不显著(P>0.05)。对于Pielou均匀度指数, RVW和RV显著大于RF(P < 0.05), RVW和RV间差异不显著(P>0.05);其中, RVW中的RVW1和RVW2显著大于RVW3(P < 0.05), RV中的RV1显著大于RV2和RV3(P < 0.05), RF中的RF1和RF2显著大于RF3(P < 0.05), 其他样点各小区间差异均不显著(P>0.05)。对于Simpson优势集中性指数, RF显著大于RVW和RV(P < 0.05), RVW和RV间差异不显著(P>0.05);RVW和RV各自小区间均相互差异显著(P < 0.05), 其他样点中的各小区间差异均不显著(P>0.05)(图 5)。

|
| 图 5 三类冬水田调查点节肢动物群落多样性(平均值±标准误) Fig. 5 Diversities of arthropod community in FWPFs and their plots of three sites (Mean±SE) |
以3个样点各小区节肢动物群落及其主要功能团的密度和生物量作相关性分析, 结果显示, 两个参数间显著相关性的为节肢动物(总密度和总生物量)、腐食者和植食者(P < 0.05), 捕食者相关性不显著(P>0.05);通过回归分析结果比较, 它们在密度和生物量间的相关性模型均可用二元线性方程式表示(表 3)。
| 节肢动物及其功能团 Arthropod and its groups |
相关系数 Correlation coefficient |
显著水平 Significant level |
R2 | 回归方程* Regression equation |
| 节肢动物Arthropod | 0.846 | 0.001 | 0.716 | Y=-91097.8+59667.1X |
| 捕食者Predator | 0.537 | 0.072 | — | — |
| 腐食者Detritivore | 0.989 | 0.000 | 0.978 | Y=-13542.2+64141.1X |
| 植食者Herbivore | 0.908 | 0.000 | 0.824 | Y=73.5+216.1X |
| Y:节肢动物密度, Density of arthropod;X:节肢动物生物量, Biomass of arthropod | ||||
以3个样点各小区捕食者及其猎物的生物量(包括平均值)作相关性分析, 结果显示, 捕食者与其猎物腐食者和植食者及它们的组合间均存在极显著的相关性(P < 0.01);根据回归结果比较分析, 它们的相关性模型均适合用二元线性方程式表示;另外, 腐食者和植食者的猎物组合与它们单独作为猎物相比, 其与捕食者线性关系的拟合程度要更好(R2=0.959, P < 0.001)(表 4)。
| 捕食者及其猎物组合 Predator and its prey combination |
相关系数 Correlation coefficient |
显著水平 Significant level |
R2 | 回归方程* Regression equation |
| 捕食者和腐食者 Predator and detritivore |
0.867 | 0.002 | 0.756 | Y=0.001+1.992X1 |
| 捕食者和植食者 Predator and herbivore |
0.810 | 0.008 | 0.657 | Y=0.003+1.525X2 |
| 捕食者和腐食者+植食者 Predator and detritivore+herbivore |
0.979 | 0.000 | 0.959 | Y=-0.001+1.873X1+1.161X2 |
| Y:捕食者, Predator;X:腐食者或植食者或两者组合, Detritivore or herbivore or the combination of them | ||||
以3个样点各小区捕食者及其猎物的密度(包括平均值)作相关性分析, 结果显示, 捕食者只与植食者间存在极显著的相关性(P < 0.01)(表 5), 与腐食者及腐食者和植食者组合间的相关性均不显著(P>0.05);根据回归结果比较分析, 捕食者与植食者间的关系模型适合用二次方程式拟合表示(R2=0.929, P < 0.001)(图 6)。
| 捕食者及猎物组合 Predator and its prey combination |
相关系数 Correlation coefficient |
显著水平 Significant level |
R2 | 回归方程* Regression equation |
| 捕食者和腐食者 Predator and detritivore |
-0.422 | 0.171 | — | — |
| 捕食者和植食者 Predator and herbivore |
0.927 | 0.000 | 0.870 | Y=139.186+0.079X+0.002X2 |
| 捕食者和腐食者+植食者 Predator and detritivore+herbivore |
-0.422 | 0.172 | — | — |
| Y:捕食者, Predator;X:植食者, Herbivore | ||||

|
| 图 6 冬水田节肢动物捕食者与植食者密度的二次方程拟合曲线 Fig. 6 The fitted curves of quadratic equation between the densities of predators and herbivores in FWPF plots |
以3个样点的节肢动物群落个体数计算Gower系数, 结果显示, RV和RF间的节肢动物群落组成相似度最高, 其次是RVW和RF, 而RVW和RV差异最大;它们的Gjk分别为0.18、0.20和0.25。以3个样点9个小区的节肢动物群落个体数计算Gower系数, 结果显示, 这9个小区间的相似性可分为4个层次, 相似性最高的两对为RVW3和RF3与RV1和RF3, Gjk均为0.19;其次是Gjk介于0.21—0.28的23对;第3层次是Gjk介于0.30—0.36的10对;最后1对RVW1和RVW2的Gjk最大, 为0.44(表 6)。
| 各样点小区 Plots of three sites |
RVW1 | RVW2 | RVW3 | RV1 | RV2 | RV3 | RF1 | RF2 | RF3 |
| RVW1 | — | ||||||||
| RVW2 | 0.44 | — | |||||||
| RVW3 | 0.31 | 0.35 | — | ||||||
| RV1 | 0.28 | 0.32 | 0.23 | — | |||||
| RV2 | 0.30 | 0.34 | 0.27 | 0.25 | — | ||||
| RV3 | 0.28 | 0.33 | 0.28 | 0.22 | 0.28 | — | |||
| RF1 | 0.28 | 0.32 | 0.24 | 0.24 | 0.26 | 0.24 | — | ||
| RF2 | 0.31 | 0.36 | 0.25 | 0.25 | 0.26 | 0.23 | 0.26 | — | |
| RF3 | 0.26 | 0.30 | 0.19 | 0.19 | 0.23 | 0.21 | 0.25 | 0.23 | — |
| RVW1:水稻、蔬菜和荒地耕作区小区1, The first plot of mixed areas of rice, vegetables and wasteland;RVW2:水稻、蔬菜和荒地耕作区小区2, The second plot of mixed areas of rice, vegetables and wasteland;RVW3:水稻、蔬菜和荒地耕作区小区3, The third plot of mixed areas of rice, vegetables and wasteland;RV1:水稻和蔬菜耕作区小区1, The first plot of mixed areas of rice and vegetables;RV2:水稻和蔬菜耕作区小区2, The second plot of mixed areas of rice and vegetables;RV3:水稻和蔬菜耕作区小区3, The third plot of mixed areas of rice and vegetables;RF1:水稻和林地耕作区小区1, The first plot of mixed areas of rice and forestland;RF2:水稻和林地耕作区小区2, The second plot of mixed areas of rice and forestland;RF3:水稻和林地耕作区小区3, The third plot of mixed areas of rice and forestland | |||||||||
以稻田7个环境因子和12个节肢动物群落参数进行相关性分析, 结果显示, 节肢动物群落参数受环境因子显著影响最大的是生境类型(FH), 其次是稻田蓄水量(EWQ)和稻桩生物量(BR), 而种植模式(PP)和收割方式(HM)影响最小;其他各因子对节肢动物群落的参数影响均不显著。其中, FH与AI、AB、C、DB和DI显著正相关(P < 0.01或0.05), 与H′、D、J和PI显著负相关(P < 0.01或0.05)。EWQ与H′(P < 0.05)和D、HB、PI和HI(P < 0.01)显著或极显著负相关;BR与H′、D、PI和HI(P < 0.05)显著正相关, 与C显著负相关(P < 0.05);PP和HM均与PI显著负相关(P < 0.05)(表 7)。
| 节肢动物群落参数 Parameters of arthropod community |
生境类型 Farmland habitat types |
种植模式 Planting patterns |
收割方式 Harvesting methods |
蓄水量 Evaluation of water quantity |
稻秆高度 Height of rice stubs |
稻桩密度 Density of rice stubs |
稻桩生物量 Biomass of rice straws |
| AI | 0.896** | 0.546 | 0.546 | 0.361 | -0.219 | -0.479 | -0.305 |
| AB | 0.685* | 0.613 | 0.613 | 0.086 | -0.216 | -0.643 | -0.298 |
| H′ | -0.843** | -0.639 | -0.639 | -0.757* | 0.562 | 0.456 | 0.704* |
| D | -0.791* | -0.595 | -0.595 | -0.828** | 0.664 | 0.385 | 0.731* |
| J | -0.791* | -0.528 | -0.528 | -0.577 | 0.457 | 0.410 | 0.575 |
| C | 0.843** | 0.641 | 0.641 | 0.621 | -0.502 | -0.441 | -0.674* |
| PB | 0.158 | 0.193 | 0.193 | -0.344 | -0.018 | -0.281 | -0.012 |
| DB | 0.791* | 0.520 | 0.520 | 0.365 | -0.215 | -0.487 | -0.300 |
| HB | -0.369 | -0.461 | -0.461 | -0.832** | 0.503 | 0.459 | 0.464 |
| PI | -0.685* | -0.686* | -0.686* | -0.950** | 0.019 | 0.456 | 0.790* |
| DI | 0.791* | 0.503 | 0.503 | 0.357 | -0.128 | -0.467 | -0.236 |
| HI | -0.211 | -0.608 | -0.608 | -0.824** | 0.608 | 0.293 | 0.703* |
| *P<0.05;**P<0.01;AI:节肢动物群落密度, Density of arthropod;AB:节肢动物群落生物量, Biomass of arthropod;H′:Shannon-Wiener多样性指数, Shannon-Wiener diversity index;D:Margalef丰富度指数, Margalef richness index;J:Pielou均匀度指数, Pielou evenness index;C:Simpson优势集中性指数, Dominant concentration index of Simpson;PB:捕食者生物量, Biomass of predator;DB:腐食者生物量, Biomass of detritivore;HB:植食者生物量, Biomass of herbivore;PI:捕食者密度, Density of predator;DI:腐食者密度, Density of detritivore;HI:植食者密度, Density of herbivore | |||||||
由于表 7中19个环境因子和节肢动物群落参数中多数变量间存在高度的线性相关以及样本量少于指标, 无法形成正定矩阵, 结合该表中环境因子与节肢动物群落参数间相关性分析结果, 选取8个对冬水田生境异质性影响较大的代表因子进行主成分分析, 包括4个环境因子FH、PP、EWQ和BR以及4个节肢动物群落参数H′、AI、PI和DI。由主成分分析可知, KMO取样足够度为0.70, Bartlett的球形度检验概率小于0.01(χ2=70.66, P=0.000), 说明利用这些因子可以进行主成分分析。本研究选取的前2个主成分累积方差贡献率达86.49%, 认为一个2因子模型可解释试验数据内容的绝大部分信息(表 8)。由于总方差的73.73%的贡献率来自第1主成分, 因此轴1(第1主成分)各因子的影响力大小可认为是对冬水田各样点生境异质性起决定性的影响因素, 影响力从大到小依次是H′、FH、AI、PI、DI、PP、EWQ和BR;其中H′、PI和BR为负相关, 其他为正相关;而综合排序结果显示, 影响力最大的前3位因子是AI、DI和FH(表 9)。
| 主成分 Principal component |
特征根 Eigenvalues |
方差贡献率/% Variance contribution |
累积方差贡献率/% Cumulative variance contribution |
| 1 | 5.90 | 73.73 | 73.73 |
| 2 | 1.02 | 12.77 | 86.49 |
| 指标 Index |
主成分1 Principal component 1 |
主成分2 Principal component 2 |
综合分值 Comprehensive values |
排序 Rank |
| FH | 0.383 | 0.043 | 0.333 | 3 |
| PP | 0.331 | -0.342 | 0.232 | 4 |
| EWQ | 0.315 | -0.302 | 0.224 | 5 |
| BR | -0.310 | 0.505 | -0.190 | 6 |
| AI | 0.379 | 0.380 | 0.379 | 1 |
| H′ | -0.393 | -0.186 | -0.362 | 8 |
| PI | -0.373 | 0.161 | -0.294 | 7 |
| DI | 0.333 | 0.575 | 0.369 | 2 |
| FH:生境类型, Farmland habitat types;PP:种植模式, Planting patterns;EWQ:蓄水量, Evaluation of water quantity;BR:稻桩生物量, Biomass of rice straws | ||||
基于上述8个生物和非生物影响因素PCA得分的冬水田排序结果如图 7所示。由图 7可知,3个冬水田样点排序图显示出了明显的生境异质性差异, RVW、RV和RF三者间的差异均较大;其中,RVW受BR、PI和EWQ影响较大, 且正或负相关;RF受DI和AI影响较大, 均呈正相关;RV受这些因子的影响均较小, 但与EWQ和H′关系较密切, 均呈正相关, 说明小区间的生境条件差异很大。此外, 由图 7还可知, 节肢动物群落参数PI和BR、AI和DI、FH和PP等的相关性均较强;对第一轴(第1主成分)影响最大的是H′、FH和AI, 各因子对第二轴(第2主成分)的影响均较小。利用上述19个生物和非生物影响因素对9个冬水田小区聚类, 结果可分为3类, RF1和RF3聚为一类, RV1、RV2和RF3聚为一类, RVW1、RVW1、RVW3和RV3聚为一类;RV和RF小区生境差异较大, 而RVW间差异最小;该结果进一步说明了3类冬水田各小区(小生境)间也存在明显的生境异质性(图 8)。

|
| 图 7 基于节肢动物群落多样性特征参数及环境因子PCA得分的冬水田类型排序图 Fig. 7 PCA ordination diagram of different sites based on characteristic parameters of community diversity of arthropods and environmental factors in FWPF RVW:水稻、蔬菜和荒地耕作区;RV:水稻和蔬菜耕作区;RF:水稻和林地耕作区;AI:节肢动物群落密度;BR:稻桩生物量;DI:腐食者密度;EWQ:蓄水量;FH:生境类型;H′:Shannon-Wiener多样性指数;PI:捕食者密度;PP:种植模式 |

|
| 图 8 基于环境因素及节肢动物群落特征参数的冬水田不同生境类型小区聚类分析 Fig. 8 The analysis map of hierarchical cluster on the similarity of different plots in three sites based on the environmental factors and community parameters of arthropods in FWPF plots |
目前, 鲜有单季中稻区休耕期冬水田节肢动物群落的研究报道。本研究发现, 重庆单季稻区休耕期冬水田节肢动物群落隶属于3纲119科(种), 按功能团可分为腐食者、捕食者、植食者、寄生者和杂食者。但从个体数或密度分析, 腐食者相对多度超过了97%, 是占绝对主导位置的功能群;而从生物量分析, 捕食者和腐食者相对生物量之和超过了86%, 是对冬水田节肢动物群落的发生起主要作用的功能群。另外, 蓄水较少的冬水田杂草较多, 这也造成植食者的数量和生物量所占比重较大。优势度指数结果指出, 冬水田节肢动物群落以罕见或稀少种(类群)数量最多以及优势类群数量稀少且突出为最主要特征。优势类群主要是腐食者中的圆
另外, 本研究也发现, 节肢动物群落密度和生物量间存在显著线性相关性, 如节肢动物(总密度和总生物量)、腐食者和植食者;因此本研究中用节肢动物群落的密度分析了节肢动物群落的多样性特征参数;尽管如此, 在分析捕食者和猎物间的营养关系时, 发现生物量比密度更能准确地反映2者间的营养关系。师光禄等[31]认为以物种生物量的大小作为多样性测度指数, 能较客观地反映物种在生物群落中的作用, 研究得出的结果灵敏性更高。另外, 戈峰等[32]通过个体数和生物量等数据计算, 比较了节肢动物群落的数量与能流多样性特征, 认为能流量多样性指数比数量多样性指数变化更为敏感, 更能反映害虫与天敌的群落特征。
4.2 冬水田水稻秸秆与节肢动物群落的生态关系分析与水稻生育期稻田相比, 冬水田系统中大量存在的水稻秸秆(稻桩)决定了该季节稻田系统中节肢动物群落功能团间的基本生态关系。本研究发现, 重庆冬水田系统腐食者以弹尾虫为主, 且圆
目前在稻田生态系统中, 研究弹尾虫的生态功能主要集中于水稻生育期, 且将弹尾虫的生态功能定义为“中性”[40-41]。但在水稻收割后冬水田休耕季节, 主要害虫(或植食性)种类密度显著降低甚至于再生稻采收后数量非常稀少, 而弹尾虫的数量或密度却非常大;由此可知这些腐食者的生态功能已不是水稻生育期建立在“水稻-害虫-天敌”三营养系统下的“中性”或主要作为天敌的猎物, 而是承担着冬水田休耕季节主要营养系统“水稻稻秆(稻桩)-腐食类(弹尾虫)-捕食者”中承上启下的双重生态功能, 即能促进对水稻残体的分解又作为捕食者的主要猎物, 这就决定了其在该系统的“中心”位置, 是影响系统的关键因子, 可看作系统“关键或中心”节肢动物类群。我们前期的研究表明, 冬水田休耕季节稻田6科11种蜘蛛均捕食弹尾虫, DNA阳性检出率超过了69.0%[5]。此外, 作物生育期大量的研究表明, 无论是间接的途径还是直接的捕食证据都说明, 弹尾虫是蜘蛛等捕食者的重要猎物源, 如通过田间观察[9, 42-43]、室内捕食功能反应[44]、单克隆抗体[45]、稳定同位素[46]以及DNA分子技术[47-48]等。同时, 在植物残体降解过程中, 弹尾虫担负着重要的分解者的角色。如在室内模拟“凋落物质量-土栖动物对凋落物的破碎化-弹尾虫密度”3者间多营养相互关系的研究表明, 弹尾虫的存在可提高凋落物质量损失率和C、N养分的矿化率, 从而加速凋落物的降解[49]。在分子水平上, 利用同位素示踪法研究发现, 弹尾虫能调节进入土壤中的有机质养分, 特别是直接作用于植物残体中有机C养分的土壤转移[50]。
4.3 环境因素对冬水田节肢动物群落组成及多样性特征的影响本研究结果表明, 冬水田典型生境类型间在节肢动物及其功能团的密度和生物量上均出现了显著性差异;而节肢动物群落多样性特征参数在各类型冬水田中呈明显的规律变化, 也出现了显著差异, 包括Shannon-Wiener多样性指数、Margalef丰富度指数、Pielou均匀度指数和Simpson优势集中性指数;而造成这些变化的原因应该与各冬水田生境条件的不同密切相关,特别是稻田周围的非作物生境。由于短期农作物生境内节肢动物群落存在群落重建的过程, 而非作物生境内的节肢动物群落又称之为作物生境内节肢动物群落的种库[8, 51];因此, 3类冬水田节肢动物群落组成和多样性特征参数的变化也会因为它们非稻田生境节肢动物群落的不同而受到重大影响。研究也指出, 越冬节肢动物群落在冬闲田和非稻田生境优势类群和多样性特征参数上明显不同[16]。另外, 水稻生育期的大量研究也表明, 稻田与非稻田生境区节肢动物及其功能团在种类组成及多样性参数上既表现出了相似性, 也出现了较大差异[12-15]。而在同一地区不同稻田生境, 节肢动物群落的结构是基本相似的, 但不同生境区的景观及生态条件也会导致群落的差异[11]。冬季油菜田周围不同非作物景观结构能特殊地影响菜田豹蛛Pardosa agrestis的活动密度和适合度相关的特性(体型大小和繁殖力);当菜地周围带状杂草路长度最大且距离相邻杂草休耕地最小时, 该豹蛛的活动密度达最高;豹蛛体型大小及其活动密度与这个带状路长度呈负相关, 但与菜地近邻的林分面积正相关;豹蛛窝卵数与其体型大小正相关[24]。
利用节肢动物群落相似性比较不同冬水田及其小区的生境异质性时发现, 冬水田节肢动物组成变化较小, 但受环境因素的影响也出现了较大不同。在这些环境因素中, 节肢动物群落参数受环境因子显著影响最大的是生境类型, 其次是稻田蓄水量和稻桩生物量, 而种植模式和收割方式影响最小。主成分分析结果指出, 3类冬水田生境异质性差异较大, 而影响这种差异的生物和非生物因素存在明显不同;而聚类分析结果进一步表明了这种差异也与它们小区的微生境异质性变化存在紧密的相关性。研究指出, 作物种植、管理和收割过程中的所有耕作措施或方式以及生境条件均不同程度地影响到农作物生境中节肢动物群落及其功能团的重建[9-10, 51];要特别对蜘蛛等捕食天敌开展保护策略[52]。另外, 稻田生境中水的有无也能对稻田蜘蛛捕食功能产生明显影响[53]。研究发现, 稻田有机物的增加与捕食者的丰度正相关, 主要原因是这些有机物促进了天敌的猎物即腐食者的种群数量[54]。在小麦生长后期, 保护性耕作农田的弹尾虫丰度显然要高于传统耕作田, 但不同种类的弹尾虫对这两种耕作系统有不同偏好性, 如土
| [1] | 陈桂权. 四川冬水田的历史变迁. 古今农业, 2014(1): 83–91. DOI:10.3969/j.issn.1672-2787.2014.01.010 |
| [2] | 刘继福. 恢复冬水田势在必行. 四川水利, 2005, 26(5): 44–45. |
| [3] | 马秀梅, 朱波, 杜泽林, 郑循华. 冬水田休闲期温室气体排放通量的研究. 农业环境科学学报, 2005, 24(6): 1199–1202. DOI:10.3321/j.issn:1672-2043.2005.06.031 |
| [4] | 魏朝富, 高明, 黄勤, 车福才, 杨剑虹, 谢德体, 蔡祖聪, 徐华. 耕种制度对西南地区冬水田甲烷排放的影响. 土壤学报, 2000, 37(2): 157–165. DOI:10.3321/j.issn:0564-3929.2000.02.002 |
| [5] | 庞帅, 王超, 王光华, 王雪芹, 祝增荣, 白耀宇. 利用18S rDNA分析冬水田蜘蛛对弹尾虫的捕食作用. 植物保护学报, 2017, 44(6): 1011–1016. |
| [6] | Neher D A, Weicht T R, Barbercheck M E. Linking invertebrate communities to decomposition rate and nitrogen availability in pine forest soils. Applied Soil Ecology, 2012, 54: 14–23. DOI:10.1016/j.apsoil.2011.12.001 |
| [7] | Chang L, Wu H T, Wu D, Sun X. Effect of tillage and farming management on Collembola in marsh soils. Applied Soil Ecology, 2013, 64: 112–117. DOI:10.1016/j.apsoil.2012.11.007 |
| [8] | 张文庆, 古德祥, 张古忍. 论短期农作物生境中节肢动物群落的重建Ⅰ.群落重建的概念及特性. 生态学报, 2000, 20(6): 1107–1112. DOI:10.3321/j.issn:1000-0933.2000.06.032 |
| [9] | 张文庆, 古德祥, 张古忍. 论短期农作物生境中节肢动物群落的重建Ⅱ.群落重建的分析和调控. 生态学报, 2001, 21(6): 1020–1024. DOI:10.3321/j.issn:1000-0933.2001.06.028 |
| [10] | Bjökman C, Bommarco R, Eklund K, Höglund S. Harvesting disrupts biological control of herbivores in a short-rotation coppice system. Ecological Applications, 2004, 14(6): 1624–1633. DOI:10.1890/03-5341 |
| [11] | 尤民生, 陶方玲, 庞雄飞. 稻田节肢动物群落相似性的研究. 华南农业大学学报, 1991, 12(2): 23–28. |
| [12] | 陶方玲, 梁广文, 庞雄飞. 不同生境区稻田节肢动物群落动态分析. 华南农业大学学报, 1996, 17(1): 25–30. |
| [13] | 俞晓平, 胡萃, Heong K L. 非作物生境对农业害虫及其天敌的影响. 中国生物防治, 1996, 12(3): 130–l33. |
| [14] | 刘雨芳, 古德祥, 张古忍. 广东双季稻区杂草地和稻田中捕食性节肢动物的群落动态. 昆虫学报, 2003, 46(5): 591–597. DOI:10.3321/j.issn:0454-6296.2003.05.009 |
| [15] | 张古忍, 古德祥, 张文庆. 稻田捕食性节肢动物群落的种库与群落的重建. 中国生物防治, 1997, 13(2): 65–68. |
| [16] | 张洁, 胡良雄, 刘杰, 凌善峰, 陈海东, 张润杰. 稻田与非稻田生境越冬节肢动物群落调查与分析. 中山大学学报:自然科学版, 2010, 49(5): 118–121. |
| [17] | 刘其全, 姚凤銮, 郑云开, 林胜, 尤民生. 武夷山稻田生境越冬节肢动物群落的物种组成. 华东昆虫学报, 2007, 16(1): 30–35. |
| [18] | 张娟, 曾玲, 梁广文, 陆永跃. 稻田越冬害虫及其捕食性节肢动物类群的空间分布动态. 环境昆虫学报, 2010, 32(3): 299–306. DOI:10.3969/j.issn.1674-0858.2010.03.002 |
| [19] | 王智. 洞庭湖平原稻田蜘蛛群落越冬生态分析. 河北师范大学学报:自然科学版, 2002, 26(3): 295–296, 308-308. |
| [20] | 李剑泉, 赵志模, 朱文炳, 侯丽娜, 周彦, 李雪燕. 重庆市稻田动物群落及农田蜘蛛资源考察. 西南农业大学学报, 2001, 23(4): 312–316. DOI:10.3969/j.issn.1673-9868.2001.04.008 |
| [21] | 闫瑞红, 白耀宇, 程家安, 叶恭银. Bt稻Cry1Ab蛋白的表达和降解及对采后季节土表灰橄榄长角跳虫发生的影响. 植物保护学报, 2009, 36(5): 431–436. DOI:10.3321/j.issn:0577-7518.2009.05.009 |
| [22] | Bai Y Y, Yan R H, Ye G Y, Huang F N, Cheng J A. Effects of transgenic rice expressing Bacillus thuringiensis Cry1Ab protein on ground-dwelling collembolan community in postharvest seasons. Environmental Entomology, 2010, 39(1): 243–251. DOI:10.1603/EN09149 |
| [23] | 徐富贤, 张林, 熊洪, 周兴兵, 蒋鹏, 朱永川, 郭晓艺, 刘茂. 不同栽培方式对杂交中稻产量及冬水田肥力的影响. 中国稻米, 2017, 23(2): 27–31. DOI:10.3969/j.issn.1006-8082.2017.02.007 |
| [24] | Drapela T, Frank T, Heer X, Moser D, Zaller J G. Landscape structure affects activity density, body size and fecundity of Pardosa wolf spiders (Araneae:Lycosidae) in winter oilseed rape. European Journal of Entomology, 2011, 108(4): 609–614. DOI:10.14411/eje.2011.079 |
| [25] | 俞晓平, 吕仲贤, 陈建明, 郑许松, 徐红星, 陶林勇, Heong K L. 水稻害虫及其天敌在生境间迁移的监测方法和原理. 浙江农业学报, 1999, 11(6): 325–332. DOI:10.3969/j.issn.1004-1524.1999.06.011 |
| [26] | 施波, 姚凤銮, 陈少波, 尤民生. 陷阱法的改进及其在稻田蜘蛛取样中的应用. 应用昆虫学报, 2011, 48(3): 782–786. |
| [27] | 徐国良, 莫江明, 周国逸. 氮沉降对三种林型土壤动物群落生物量的影响. 动物学研究, 2005, 26(6): 609–615. DOI:10.3321/j.issn:0254-5853.2005.06.006 |
| [28] | 钱迎倩, 马克平. 生物多样性研究的原理与方法. 北京: 中国科学技术出版社, 1994. |
| [29] | Widyastuti R. Population dynamics of microarthropods (Oribatida and Collembola) in rainfed paddy field ecosystem in Pati, central Java. Jurnal Ilmu Tanah & Lingkungan, 2005, 7(1): 11–14. |
| [30] | 祝向钰, 李志毅, 常亮, 袁一扬, 戈峰, 吴刚, 陈法军. 转Bt水稻土壤跳虫群落组成及其数量变化. 生态学报, 2012, 32(11): 3546–3554. |
| [31] | 师光禄, 席银宝, 王海香, 李捷, 赵飞. 枣园节肢动物群落的数量与生物量多样性特征分析. 林业科学, 2004, 40(2): 107–112. |
| [32] | 戈峰, 李典谟, 谢宝瑜, 丁岩钦, 刘向辉. 棉田节肢动物群落的数量与能流量多样性特征分析. 生态学报, 2000, 20(6): 971–976. DOI:10.3321/j.issn:1000-0933.2000.06.011 |
| [33] | 刘志龙, 王连生, 杜一新, 俞晓平, 吕仲贤, 郑许松, 梅晓青, 潘慧萍. 捕食性天敌在单季稻田与非稻田生境间的迁移规律及其保护利用. 浙江农业学报, 1999, 11(6): 344–348. DOI:10.3969/j.issn.1004-1524.1999.06.014 |
| [34] | Carpenter S R, Kitchell J F, Hodgson J R. Cascading trophic interactions and lake productivity:fish predation and herbivory can regulate lake ecosystems. BioScience, 1985, 35(10): 634–639. DOI:10.2307/1309989 |
| [35] | McQueen D J, Post J R, Mills E L. Trophic relationships in freshwater pelagic ecosystems. Canadian Journal of Fisheries and Aquatic Sciences, 1986, 43(8): 1571–1581. DOI:10.1139/f86-195 |
| [36] | Patrick L B, Kershner M W, Fraser L H. Epigeal spider responses to fertilization and plant litter:testing biodiversity theory at the ground level. Journal of Arachnology, 2012, 40(3): 309–324. DOI:10.1636/P11-92.1 |
| [37] | Chen B R, Wise D H. Bottom-up limitation of predaceous arthropods in a detritus-based terrestrial food web. Ecology, 1999, 80(3): 761–772. DOI:10.1890/0012-9658(1999)080[0761:BULOPA]2.0.CO;2 |
| [38] | Saitoh S, Fujii S, Takeda H. Evaluation of the bottom-up force of accumulated organic matter on microarthropods in a temperate forest floor. European Journal of Soil Biology, 2011, 47(6): 409–413. DOI:10.1016/j.ejsobi.2011.07.013 |
| [39] | Lawrence K L, Wise D H. Spider predation on forest-floor Collembola and evidence for indirect effects on decomposition. Pedobiologia, 2000, 44(1): 33–39. DOI:10.1078/S0031-4056(04)70026-8 |
| [40] | 吴进才, 胡国文, 唐健, 束兆林, 杨金生, 万志农, 任正才. 稻田中性昆虫对群落食物网的调控作用. 生态学报, 1994, 14(4): 381–386. DOI:10.3321/j.issn:1000-0933.1994.04.015 |
| [41] | 郭玉杰, 王念英, 蒋金炜, 陈俊炜, 唐建. 中性昆虫在稻田节肢动物群落中作为捕食者营养桥梁作用的研究. 中国生物防治, 1995, 11(1): 5–9. |
| [42] | 刘雨芳, 古德祥. 稻田中性昆虫多样性及其生态功能分析. 中国生物防治, 2002, 18(4): 149–152. |
| [43] | 尤民生, 侯有明, 刘雨芳, 杨广, 李志胜, 蔡鸿娇. 农田非作物生境调控与害虫综合治理. 昆虫学报, 2004, 47(2): 260–268. DOI:10.3321/j.issn:0454-6296.2004.02.021 |
| [44] | 庞保平, 程家安, 王启法. 稻田蜘蛛对弹尾虫的功能反应及选择作用. 植物保护学报, 1998, 25(3): 193–196. DOI:10.3321/j.issn:0577-7518.1998.03.001 |
| [45] | 赵伟春, 王光华, 程家安, 陈正贤. 弹尾虫单克隆抗体的制备及其在捕食研究中的应用. 生态学报, 2007, 27(9): 3694–3700. DOI:10.3321/j.issn:1000-0933.2007.09.018 |
| [46] | Park H H, Lee J H. Arthropod trophic relationships in a temperate rice ecosystem:a stable isotope analysis with δ13C and δ15N. Environmental Entomology, 2006, 35(3): 684–693. DOI:10.1603/0046-225X-35.3.684 |
| [47] | Agustí N, Shayler S P, Harwood J D, Vaughan I P, Sunderland K D, Symondson W O C. Collembola as alternative prey sustaining spiders in arable ecosystems:prey detection within predators using molecular markers. Molecular Ecology, 2003, 12(12): 3467–3475. DOI:10.1046/j.1365-294X.2003.02014.x |
| [48] | Kuusk A K, Agustí N. Group-specific primers for DNA-based detection of springtails (Hexapoda:Collembola) within predator gut contents. Molecular Ecology Resources, 2008, 8(3): 678–681. DOI:10.1111/j.1471-8286.2007.02046.x |
| [49] | Yang X D, Yang Z, Warren M W, Chen J. Mechanical fragmentation enhances the contribution of Collembola to leaf litter decomposition. European Journal of Soil Biology, 2012, 53: 23–31. DOI:10.1016/j.ejsobi.2012.07.006 |
| [50] | Chamberlain P M, McNamara N P, Chaplow J, Stott A W, Black H I J. Translocation of surface litter carbon into soil by Collembola. Soil Biology and Biochemistry, 2006, 38(9): 2655–2664. DOI:10.1016/j.soilbio.2006.03.021 |
| [51] | 张文庆, 张古忍, 古德祥. 论短期农作物生境中节肢动物群落的重建Ⅲ.群落重建与天敌保护利用. 生态学报, 2001, 21(11): 1927–1931. DOI:10.3321/j.issn:1000-0933.2001.11.024 |
| [52] | 王洪全. 中国稻区蜘蛛群落结构和功能的研究. 长沙: 湖南师范大学出版社, 2006. |
| [53] | 杨国庆, 吴进才, 王洪全, 姜永厚, 邱慧敏. 水对稻田蜘蛛捕食功能的影响. 生态学报, 2003, 23(4): 681–687. DOI:10.3321/j.issn:1000-0933.2003.04.008 |
| [54] | Settle W H, Ariawan H, Astuti E T, Cahyana W, Hakim A L, Hindayana D, Lestari A S. Managing tropical rice pests through conservation of generalist natural enemies and alternative prey. Ecology, 1996, 77(7): 1975–1988. DOI:10.2307/2265694 |
| [55] | Dittmer S, Schrader S. Longterm effects of soil compaction and tillage on Collembola and straw decomposition in arable soil. Pedobiologia, 2000, 44(3/4): 527–538. |