 2018, Vol. 38
2018, Vol. 38文章信息
- 田俊霞, 魏丽萍, 何念鹏, 徐丽, 陈智, 侯继华.
- TIAN Junxia, WEI Liping, HE Nianpeng, XU Li, CHEN Zhi, HOU Jihua.
- 温带针阔混交林叶片性状随树冠垂直高度的变化规律
- Vertical variation of leaf functional traits in temperate forest canopies in China
- 生态学报. 2018, 38(23): 8383-8391
- Acta Ecologica Sinica. 2018, 38(23): 8383-8391
- http://dx.doi.org/10.5846/stxb201801020006
-
文章历史
- 收稿日期: 2018-01-02
- 网络出版日期: 2018-09-26





2. 中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室, 北京 100101
2. Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China
植物功能性状(Plant functional trait or plant trait)简称为植物性状, 是植物在进化和发展过程中对外界环境的响应和适应的特定参数, 它与植物资源利用策略、生长策略和适应策略密切相关[1-3]。各种植物功能性状间的权衡和组合, 较大程度决定着植物的生活史对策[4]。因此, 开展植物功能性状的研究不仅有助于更好地理解生态系统功能, 还可为更准确预测生态系统对环境变化的响应和适应提供重要的理论基础[5]。
叶片是植物进行光合作用的主要器官, 可为植物生长提供能量。叶片性状对外界环境变化较为敏感, 能够反映植物捕获光资源的能力以及应对环境变化所采取的生存对策[6]。表征叶片性状的指标较多, 叶片比叶面积(SLA)、叶片干物质含量(LDMC)、叶片氮含量(N)、叶片磷含量(P)、氮磷比(N : P)和叶绿素含量(Chl)是常用的性状指标。其中, SLA可表征植物对光、热和水资源的利用效率, 是反映植物碳收获策略的主要性状[7-8];LDMC为叶片的干重与鲜重之比, 能够反映植物利用资源的能力以及适应环境的生存对策[9];叶片N、P在植物的生长和生理机能调节方面发挥着重要作用[10], 二者既相互影响又相互独立, 可以反映森林生态系统的养分利用和循环效率;N : P可以反映森林群落树种的营养限制状况, 表征特定森林立地条件的N或P限制状况[11];叶绿素是植物光合作用的主要光合色素, Chl可一定程度的反映植物叶片的光合能力[12-13]。
目前, 科研人员针对植物叶片性状开展了大量研究工作, 主要包括个体尺度上叶片生理和结构特性研究[14-16];区域尺度上环境因子如海拔、纬度、温度、水分和土壤等对植物叶片性状影响的研究[17-19]。现有的研究主要是分析或比较了多个物种叶片性状之间的差异, 或是探讨叶片性状在不同环境梯度、大尺度范围内的差异[20-23]。在自然界中, 森林植物叶片性状随树冠高度常呈现明显的垂直分布现象, 然而目前对于叶片性状随着树冠垂直高度增加的变化规律仍不清楚, 且相关研究极少。叶片性状随树冠垂直高度的变化被认为是光照在树冠垂直高度的变化引起的, 水分和温度等因素也可能对其产生影响[24]。自树冠顶部到树冠底层, 光照、温度和水分等生态因子存在着明显差异。不同层植物叶片在光照、水分吸收和元素利用等方面均存在差异, 而植物对资源的高效分配和利用存在着权衡关系, 投资策略也不同。树冠上层叶片处于高辐射环境, 易于缺水, 会影响植物叶片对于N和P元素的吸收利用和再分配[10], 下层叶片受光率普遍较低, 此外不同层叶片受到风力、温度和水分的影响较大, 叶片的生理生化特征也会发生相应变化。在已有的研究中, 叶片往往采取随机取样, 未考虑树冠垂直分层现象, 那么这种随机采集植物叶片是否会影响叶片性状或叶片相关试验的研究结果?是否会影响叶片水分和养分利用效率之间的关系以及群落中树种共存的机制?
本文以中国东北分布最广泛的红松针阔混交林为基础, 选取群落内8种优势植物为研究对象, 定量研究了不同植物沿9个冠层垂直高度的6个常规叶片性状(SLA、LDMC、N、P、N : P和Chl等)随树冠垂直高度的变化规律, 为拓展植物性状的研究提供更多的理论依据和技术支持。本研究的主要目的:(1)分析叶片性状在树冠不同层之间的差异;(2)揭示叶片性状随树冠垂直高度的变化规律;(3)探讨叶片性状间的相互关系。
1 材料与方法 1.1 研究区概况实验样地位于吉林省蛟河市林业实验区管理局林场内(43°57′N, 127°44′E), 地势东南较为平坦西北较高。实验区域地处于北温带的中纬度, 受季风影响较大, 冬寒冷, 夏温暖, 气温年较差和日较差较大;降水集中在6—8月, 平均降水量为700—800 mm。土壤类型主要为暗棕色森林土, 适合各种植物的生存。研究区域植被属于温带典型针阔混交林, 除针叶树种红松外(Pinus koraiensis Sieb. et Zucc), 落叶阔叶树种主要包括核桃楸(Juglans mandshurica)、水曲柳(Fraxinus mandschurica Rupr.)、蒙古栎(Quercus mongolica Fisch. ex Ledeb.)、紫椴(Tilia amurensis Rupr.)、白桦(Betula platyphylla Suk.)、色木槭(Acer mono Maxim.)、大青杨(Populus ussuriensis)和千金榆(Carpinus cordata Bl.)等。灌木主要有毛榛(Corylus mandshurica Maxim.)和东北鼠李(Rhamnus schneideri Levl. et Vaniot var. manshurica Nakai)等;主要草本植物有东北蹄盖蕨(Athyrium brevifrons Nakai ex Kitagawa)、山茄子(Brachybotrys paridiformis Maxim. ex Oliv.)等, 还有山葡萄(Vitis amurensis Rupr.)和狗枣猕猴桃(Actinidia kolomikta(Maxim. & Rupr.))等藤本植物。
1.2 实验样地设置和样品采集2012年7—8月, 依托林场开展的森林碳含量估算项目;我们首先在选择的典型天然红松针阔混交林里建立4块100 m×100 m的样地, 并开展群落调查工作。根据群落调查结果统计样地内各树种的重要值和相对生物量, 选取重要值大于0.05的8个优势树种开展相关研究, 8个优势物种分别为红松、水曲柳、核桃楸、色木槭、蒙古栎、大青杨、千金榆和白桦。这些树种约占群落生物量的80.6%。在实际操作过程中, 每个物种选择树高相对一致、树干生长良好、无严重变形和病害的成树5—7株, 共计48株;挖掘前分别测量每株植物的胸径和冠幅等指标(表 1)。
| 树种 Tree species |
胸径 Mean diameter at breast height/cm |
树高 Mean treeheight/m |
树冠高度 Mean crown height/m |
株数 Number of plants |
| 红松Pinus koraiensis Sieb. Et Zucc. | 34.2±2.6 | 18.9±1.4 | 12.1±1.3 | 6 |
| 水曲柳Fraxinus mandschurica Rupr. | 29.9±7.9 | 21.9±1.6 | 11.6±1.3 | 7 |
| 核桃楸Juglans mandshurica Maxim. | 30.4±2.9 | 21.2±1.1 | 11.5±1.8 | 6 |
| 色木槭Acer mono Maxim. | 33.9±2.9 | 19.2±1.1 | 12.4±1.6 | 6 |
| 蒙古栎Quercus mongolica Fisch. ex Ledeb. | 30.3±2.7 | 21.3±1.0 | 12.6±1.5 | 6 |
| 大青杨Populus ussuriensis | 32.2±3.1 | 22.2±1.4 | 11.8±1.3 | 6 |
| 白桦Betula platyphylla Suk. | 29.6±2.5 | 22.6±1.2 | 13.2±1.7 | 5 |
| 千金榆Carpinus cordata Bl. | 28.5±2.2 | 20.1±1.3 | 12.2±1.1 | 6 |
目标树整株伐倒后, 用米尺准确测量树高(H)和树冠高度(CH), 记录第一活枝下高位置, 作为取样的起点, 至树冠顶端将树冠等距离分为九等份, 并分为上、中、下三层(图 1)。每一层取样时, 在靠近主干和树冠边缘的各个方向随机采取若干成熟叶片, 再用SPAD-502plus便携式叶绿素仪测定5次叶绿素含量, 取平均值。然后采集20片左右完全展开、无病虫害、成熟健康叶片冷藏保存。回到实验室后尽快对样品进行测量。其中, 红松叶片样品的采集是在每一层上选取标准枝上不少于5个不再分枝的枝条。

|
| 图 1 植物冠层叶片性状的垂直取样图 Fig. 1 The picture of vertical sampling for leaf traits in forest canopy |
本研究选取了SLA、LDMC、N、P、N : P和叶绿素值6个常规的叶片性状指标(表 2)。将所采集的叶片样品表面擦洗干净, 每种叶片5个为一组, 避开主叶脉, 均匀选取3个点, 用精度为0.01 mm的数显游标卡尺测定叶厚度, 取其平均值。用电子天平测定叶片鲜重(精度0.01 g), 用HP Scanjet G3110扫描仪(Hewlett-Packard Development Company, PaloAlto, USA)扫描所有叶片面积, 应用ImageJ软件(National Institutes of Health, Bethesda, USA)计算叶面积。用电子天平称取每物种20片叶片的饱和鲜重(精度0.01 g), 然后在85℃下烘干48 h至恒重, 分别称量干重。采用凯氏定氮法测定叶片全N含量, 采用钼锑抗比色法测定叶片全P含量。测定方法见表 2, 部分指标的计算公式如下:

|
(1) |
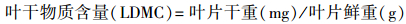
|
(2) |
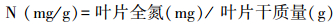
|
(3) |

|
(4) |
| 指标Indicators | 测定方法Measurement methods | 指示意义Denotative meaning |
| 比叶面积SLA Specific leaf area | 应用HP Scanjet G3110扫描仪获得叶片面积, 应用ImageJ软件计算叶面积 | 反映植物对营养的获取与利用、相对生长速率、光合效率和资源利用等 |
| 叶干物质含量LDMC Leaf dry matter content | 叶片干重与鲜重的比值 | 反映植物对养分的保有能力, 是资源获取轴上比较稳定的预测指标 |
| 叶片氮含量N N content | 叶片全氮的测定采用凯氏定氮法 | 反映生态系统植物的生长状况、养分循环及环境状况 |
| 叶片磷含量P P content | 叶片全磷的测定采用钼锑抗比色法 | 反映生态系统的植物生长状况、养分循环及环境状况 |
| 叶绿素含量Chl Chlorophyll content | 用Minolta SPAD502叶绿素仪直接测定植株叶片叶绿素含量 | 叶绿素是绿色植物光合作用的重要结构体, 可以反映植物的光合能力 |
研究计算了8个树种所有叶片性状指标的算数平均值和标准误差;采用单变量双因素(树种和高度)方差分析检验叶片性状在树种和冠层高度之间的差异。采用Fisher最小显著差数(LSD)法检验各性状在树冠不同垂直高度间的显著性差异, 对叶片性状间进行Pearson相关检验和回归分析。不同树种叶片性状与树冠相对位置关系分析采用Pearson相关性分析, 并对与树冠位置显著相关的性状进行线性拟合。统计分析使用SPSS 17.0(SPSS, Chicago, USA), 作图在Sigmaplot 12.0(SystatSoftware, San Jose, USA)软件中完成。显著性检验水平为P =0.05。
2 结果与分析 2.1 不同冠层高度下各物种植物叶片性状的差异表 3显示树种对叶片性状(SLA、LDMC、N、P、N : P和Chl)具有显著影响(P < 0.001), 冠层高度对叶片的LDMC和SLA影响显著(P < 0.05)。对于各个植物物种, 都至少有一个叶片性状在不同树冠高度表现出差异, 且不同树种表现出差异的性状不一致(表 4)。其中, 在不同树冠层次, 白桦6个叶片性状均差异显著(P < 0.05);除Chl外, 水曲柳叶片的LDMC、SLA、N、P和N : P差异显著(P < 0.05);色木槭叶片的SLA、N、P和N : P和千金榆叶片的LDMC、N、P和N : P差异显著(P < 0.05);大青杨只有P和Chl差异显著(P < 0.05);核桃楸叶片的LDMC和N差异显著(P < 0.05);红松叶片的N和P差异显著(P < 0.05)。对于蒙古栎, 其Chl的SPAD变化范围为40.90—44.17, 在不同冠层高度间差异显著, 但其他叶片性状没有显著差异(表 4)。
| 指标Indicators | LDMC | SLA | N | P | N:P | Chl | |||||||||||
| F | P | F | P | F | P | F | P | F | P | F | P | ||||||
| 树种Tree species | 2.795 | < 0.001 | 22.825 | < 0.001 | 38.677 | < 0.001 | 12.623 | < 0.001 | 6.022 | < 0.001 | 4.764 | < 0.001 | |||||
| 高度Height | 3.280 | 0.039 | 3.764 | 0.025 | 2.398 | 0.315 | 2.476 | 0.086 | 1.162 | 0.315 | 0.373 | 0.689 | |||||
| 树种×高度 Tree species×Height |
0.748 | 0.724 | 2.031 | 0.017 | 0.720 | 0.133 | 2.100 | 0.013 | 1.446 | 0.133 | 0.923 | 0.525 | |||||
| 性状Traits | 分层 | 白桦 | 水曲柳 | 蒙古栎 | 色木槭 | 大青杨 | 千金榆 | 核桃楸 | 红松 |
| 比叶面积 | 上层 | 16.75±2.40b | 80.80±23.83b | 11.35±9.24a | 76.12±2.48b | 4.81±1.35a | 7.69±0.97a | 187.63±26.40a | 31.78±11.39a |
| SLA/cm2 | 中层 | 20.81±3.41ab | 96.0±10.40ab | 12.90±2.66a | 83.17±2.96ab | 5.33±0.44a | 10.11±1.99a | 217.35±20.04a | 34.27±8.89a |
| 下层 | 21.95±2.02a | 110.4±26.24a | 13.79±1.93a | 98.31±1.70a | 6.69±0.63a | 11.61±1.87a | 223.39±19.71a | 25.32±4.64a | |
| 叶干物质含量 | 上层 | 411.79±14.14a | 433.21±3.07a | 418.2±31.70a | 440.90±22.67a | 451.78±11.72a | 434.34±6.71a | 482.24±6.54a | 496.28±17.74a |
| LDMC/(g/kg) | 中层 | 405.93±8.09b | 419.64±4.20b | 416.38±9.11a | 440.42±12.63a | 448.44±5.89a | 395.23±10.93b | 428.13±12.16b | 492.12±11.58a |
| 下层 | 385.55±7.51c | 381.73±4.56c | 388.60±4.61a | 417.78±13.71a | 416.96±18.15a | 371.55±5.73b | 421.37±13.78b | 490.0±14.63a | |
| 氮含量N / (mg/g) | 上层 | 21.14±0.89b | 23.44±0.45c | 23.35±0.31a | 25.06±0.65a | 13.62±0.73a | 21.11±0.14b | 31.15±0.53b | 14.68±0.36b |
| 中层 | 24.20±1.57ab | 26.99±0.43b | 22.81±0.33a | 23.06±0.40b | 14.41±0.43a | 23.18±0.16b | 32.39±0.34ab | 14.13±0.27b | |
| 下层 | 27.25±1.22a | 29.67±0.59a | 22.97±0.25a | 23.39±0.19b | 14.55±0.49a | 24.87±0.56a | 33.23±0.42a | 15.56±0.20a | |
| 磷含量 | 上层 | 1.45±0.05b | 1.55±0.04b | 1.44±0.09a | 1.68±0.22a | 1.23±0.05b | 1.60±0.03b | 1.85±0.04a | 1.10±0.06b |
| P/(mg/g) | 中层 | 1.53±0.05a | 1.70±0.04a | 1.40±0.03a | 1.32±0.03b | 1.30±0.02ab | 1.71±0.01b | 1.89±0.03a | 1.11±0.04b |
| 下层 | 1.46±0.03ab | 1.76±0.03a | 1.48±0.03a | 1.44±0.03b | 1.37±0.02a | 2.04±0.04a | 1.92±0.02a | 1.31±0.04a | |
| N:P | 上层 | 14.51±0.31b | 15.21±0.43b | 16.57±0.98a | 16.21±1.26b | 11.45±0.73a | 13.19±0.07a | 16.94±0.38a | 13.79±0.58a |
| 中层 | 16.05±0.93a | 16.12±0.34ab | 16.46±0.40a | 17.62±0.30a | 11.50±0.40a | 13.54±0.12a | 17.16±0.22a | 13.42±0.51a | |
| 下层 | 18.84±0.89a | 16.88±0.37a | 15.59±0.24a | 16.49±0.20ab | 10.82±0.37a | 12.20±0.20b | 17.37±0.23a | 12.61±0.35a | |
| 叶绿素含量 | 上层 | 40.73±0.66b | 42.61±4.49a | 44.17±2.04a | 45.00±1.14a | 41.14±1.07b | 36.21±1.43a | 41.63±3.10a | — |
| (Chl, SPAD) | 中层 | 39.25±0.48ab | 41.79±1.25a | 41.49±0.81ab | 43.85±0.72a | 46.09±0.47a | 38.03±1.23a | 43.76±1.32a | — |
| 下层 | 38.01±0.46a | 39.09±1.52a | 40.90±0.59b | 43.63±0.67a | 45.21±0.96a | 37.18±1.44a | 44.69±1.66a | — | |
| 同列数据后具有相同字母表示数据间差异不显著(P < 0.05) | |||||||||
白桦、蒙古栎、色木槭、大青杨、千金榆和核桃楸的SLA随树冠垂直高度的增加呈显著减小趋势。所有树种的LDMC随树冠垂直高度的增加呈增加趋势, 其中白桦、大青杨、千金榆和红松的LDMC与树冠垂直高度呈显著正相关。白桦叶片的Chl和水曲柳叶片N含量与树冠垂直高度呈显著线性相关关系, 随着树冠垂直高度的增加, 白桦叶片的Chl呈线性减少而水曲柳叶片的N含量呈线性增长。千金榆、水曲柳和大青杨的叶片P随树冠垂直高度的增加呈减小的趋势。然而, 所有树种的N : P与随树冠垂直高度关系不显著(图 2)。

|
| 图 2 叶功能性状随树冠垂直高度的变化 Fig. 2 Changes in leaf functional traits along the vertical height of forest canopy 其中, *P < 0.05; * * P < 0.01 |
对于不同植物功能群, 其叶片性状随树冠高度的变化呈现的变化趋势不同。其中, 随着树冠垂直高度的增加, 阔叶树种的SLA呈线性减少, 而针叶树种和阔叶树种的LDMC均呈线性增长。Chl、N、P和N : P等叶片性状与树冠垂直高度无显著的相关关系(图 2)。
2.3 叶片性状间相互关系Pearson相关性分析表明, 叶性状之间普遍存在着显著或极显著相关关系(表 5)。其中, SLA与N、P和N : P间呈极显著正相关(P < 0.01), 与Chl显著正相关(P < 0.05);N与P和N : P极显著正相关(P < 0.01);LDMC与其他性状无显著相关关系(表 5)。不区分物种差异, 将所有树种的SLA与N和P含量进行最优拟合, 发现叶片性状SLA和N、P含量均呈显著的线性相关关系, 随着N和P含量的增加, 植物叶片的SLA呈线性增长(图 3)。
| LDMC | SLA | N | P | N:P | Chl | |
| LDMC | 1 | |||||
| SLA | -0.155 | 1 | ||||
| N | -0.080 | 0.662** | 1 | |||
| P | -0.185 | 0.467** | 0.782** | 1 | ||
| N:P | 0.089 | 0.329** | 0.405** | -0.126 | 1 | |
| Chl | -0.038 | 0.259* | 0.089 | 0.031 | -0.044 | 1 |
| * P < 0.05;* * P < 0.01 | ||||||

|
| 图 3 比叶面积与氮磷含量的关系 Fig. 3 The relationship between specific leaf area and nitrogen (N) and phosphorus (P) content |
叶片性状在树冠不同高度之间存在显著差异, 尤其是白桦、水曲柳、色木槭和千金榆等树种。不同树种不同层间叶片性状指标差别较大, 如核桃楸和水曲柳的SLA明显大于其他树种、白桦N含量最大值出现在下层、而P含量最大值却出现在中层, 且P含量上下两层差异较小、色木槭的N和P含量的最大值均出现在上层(表 3)。光照强度和水分等因素可能是造成上述差异的重要原因。一般地, 树冠上层叶片接受到的光照强度较大, 水分易于损失, 而下层叶片光照强度相对较弱, 植物会通过调整自身组成和形态结构来适应外界环境。处于上层的叶片会通过减小SLA来减少水分的散失, 并且会增加叶片厚度来适应强光, 下层叶片则会通过增加SLA来提高吸收阳光、无机盐和CO2的能力[25], 因此上层叶片相对较小、较厚, 而下层叶片较薄、较大。树种对光环境和水环境的适应策略使得树冠不同层次间叶片性状产生差异, 充分体现出植物随着高度的变化采取不同的资源利用策略来适应外界环境的变化。
从物种水平来看, 植物自身的特性和群落树种的演替进程也可能是树种在不同树冠高度间存在差异的原因。从植物自身特性来看, 白桦、蒙古栎、大青杨和核桃楸为喜光树种, 红松、千金榆和水曲柳为中性树种, 喜光树种和中性树种叶片的形态和解剖结构不同, 其中喜光树种的叶片排列稀疏, 角质层比较发达, 单位面积上气孔较多、叶脉密、机械组织发达, 光合速率和代谢速率较高;而中性树种角质层较薄, 气孔和叶绿体数量较阳性树种较少, 光合速率和呼吸速率都较低, 对光的需求也低于阳性树种[26]。此外, 由于植物利用资源的总量有限, 若一方面资源投入大则其他方面的资源投入就会相应减少, 树种自身特性引起的叶片物质投资和分配格局的变化也会造成不同冠层间叶片性状的差异[27]。从群落树种演替进程来看, 群落不同演替时期的先锋树种和优势种不同, 并且随着演替的推进, 林冠郁闭度逐渐增加, 生存环境趋于复杂, 植物种间竞争越发激烈, 群落中的先锋树种例如白桦相对于中后期的优势树种如蒙古栎逐渐失去竞争力, 也可能会引起不同树种的叶片性状在不同冠层之间的差异[28]。
3.2 叶片性状随树冠垂直高度增加具有明显的变化趋势随着树冠高度的增加, 植物叶片性状呈现出不同的变化趋势。其中, 阔叶树种的SLA呈显著下降趋势, 针叶树种无明显变化趋势, 但所有树种的LDMC均呈上升趋势(图 2), 这与已有的研究结论基本一致[23]。之所以呈现这种变化趋势是因为光在冠层中的垂直分布变化明显, 植物叶片为适应这种变化采取增减叶片厚度或密度的方法来适应不同强度的光, 树冠上层光照辐射较强, 水分蒸发较强, 叶片处于较为缺水的环境, 因此阔叶树种的叶片会通过减少受光面积、关闭气孔和增加角质层厚度来减少水分流失, 避免失水, 同时为了将水分运回到叶肉细胞当中, 会将大量的生物量投资于微管和厚壁组织, 用于同化和支持结构的建设, 使得分配到叶肉细胞的生物量相对较少[29];而针叶树种各个层次的叶片角质层均较厚, 能有效减少水分的蒸发, 故各个层次叶片的SLA差异较小。本研究发现色木槭和水曲柳的LDMC在达到最高点后均出现了部分下降(图 2B), 说明叶片对资源的需求在一定程度上存在非线性的响应, 这是因为在树冠顶部存在一定的水分胁迫现象, 水分和光可能是制约叶片LDMC的主要因素[30]。叶片N和P含量随树冠垂直高度的增加没有表现出明显一致的变化规律, 这可能是因为叶片N和P含量受光照强度、水分、风力大小、养分利用方式、自身遗传因素[7]、土壤中N和P元素含量以及演替的进程等因素的综合影响。本研究中8种优势树种的Chl变化幅度较小, 其SPAD值均在40附近上下波动, 说明冠层高度对植物Chl的影响较小[31]。
3.3 叶片性状间的相关性本研究发现叶片的SLA与N、P呈显著的正相关关系, 这与前人的研究结论一致[32-33]。SLA、N和P作为植物叶片关键功能性状, 分别表征植物对光的捕获能力和同化CO2的能力[34]。其中, SLA是叶片对光环境的一种适应, SLA的大小会直接或间接影响植物的光合速率, 通常情况下, SLA越大则植物叶片可获得更多的光源, 提高其光合作用能力。植物叶片约75%的氮存储于叶绿体中, 叶片N含量直接影响叶绿素含量, 同时对叶片内参与光合作用的酶含量和酶活性产生影响, 进而影响光合速率[35]。叶片N是影响植物光合作用的主要限制因子[36], 叶片N与SLA呈显著的正相关关系, SLA增大有助于提高叶片N含量, 增强光合作用能力。叶片P含量也会通过影响叶绿素和蛋白质的含量来影响植物叶片的光合速率。叶片P与SLA呈显著正相关关系, 说明SLA对叶片获取光源有很大的贡献, 有助于树木叶片光合能力的增强[37-38]。本研究中树木叶片的SLA与N : P呈正相关关系, 不仅说明植物叶片通过SLA、N和P功能性状之间的协同作用, 提高光合作用效率, 同时也说明在植物叶片光合作用过程中, N对其的影响更大[39]。而叶片N和P之间显著的正相关关系, 进一步说明N和P之间协同作用来提高植物的光合能力。
通常情况下, 植物叶片LDMC与SLA呈负相关关系, 这与Vile等[40-41]的研究结果一致, 叶片的这种关系可作为植物资源分类轴上划分植物种类的最佳变量[30]。LDMC与SLA和N、P在树冠垂直方向上的负相关关系, 表明了植物随着树高的变化对自然资源分配和利用上的差异。其中, 低SLA的树种其叶和根的寿命较长, 有利于植物在干旱条件下构建防卫机构, 适应干旱恶劣的环境。
综上所述, 树冠垂直高度对温带针阔混交林中8种优势树种叶片性状具有显著影响, 且不同树种的部分叶片性状在不同冠层高度存在差异。阔叶树种SLA随着树冠垂直高度的增加而减小, 而针叶树种则呈现相反的趋势;所有树种的LDMC随着树冠垂直高度的增加而增加, 但N、P、N : P和Chl随着树冠垂直高度的增加没有显著的变化规律;在温带针阔混交林冠层中, SLA与N、P和N : P均存在显著的正相关关系。温带针阔混交林内优势树种通过改变各自叶片性状来适应外界环境的改变, 反映了森林群落树种的共生策略。叶片性状随树冠高度的变化而变化反映了植物对光环境的适应, 同时也体现植物对水资源的利用策略, 是植物与环境综合作用的结果。对于本研究地区的树种而言, 光照和水分可能是主要影响因素, 这对未来利用激光雷达技术探测树高, 并建立模型来估算区域生物生产力具有重要意义, 同时也对后续开展叶片性状的相关研究时是否需要考虑叶片垂直分层所带来的影响具有一定的指导意义。但目前针对叶片性状随树冠垂直高度变化规律的研究还非常少, 本文的许多现象和推测有待进一步证实。
| [1] | Blonder B, Baldwin B G, Enquist B J, Robichaux R H. Variation and macroevolution in leaf functional traits in the Hawaiian silversword alliance (Asteraceae). Journal of Ecology, 2016, 104(1): 219–228. DOI:10.1111/1365-2745.12497 |
| [2] | Santiago L S, Wright S J. Leaf functional traits of tropical forest plants in relation to growth form. Functional Ecology, 2007, 21(1): 19–27. |
| [3] | 刘晓娟, 马克平. 植物功能性状研究进展. 中国科学:生命科学, 2015, 45(4): 325–339. |
| [4] | Johnston R F, Frank P W, Michener C D. Annual review of ecology and systematics. Taxon, 1971, 20(4): 626–626. DOI:10.2307/1218271 |
| [5] | Cadotte M W, Arnillas C A, Livingstone S W, Yasui S E. Predicting communities from functional traits. Trends in Ecology & Evolution, 2015, 30(9): 510–511. |
| [6] | 毛伟, 李玉霖, 张铜会, 赵学勇, 黄迎新, 宋琳琳. 不同尺度生态学中植物叶性状研究概述. 中国沙漠, 2012, 32(1): 33–41. |
| [7] | Song L Y, Peng C L, Peng S L. Comparison of leaf construction costs between three invasive species and three native species in South China. Biodiversity Science, 2009, 17(4): 378–384. DOI:10.3724/SP.J.1003.2009.09077 |
| [8] | 宝乐, 刘艳红. 东灵山地区不同森林群落叶功能性状比较. 生态学报, 2009, 29(7): 3692–3703. DOI:10.3321/j.issn:1000-0933.2009.07.030 |
| [9] | Liu J H, Zeng D H, Lee D K, Fan Z P, Zhong L. Leaf traits and their interrelationship of 23 plant species in southeast of Keerqin Sandy Lands, China. Frontiers of Biology in China, 2008, 3(3): 332–337. DOI:10.1007/s11515-008-0050-x |
| [10] | Ryther J H, Dunstan W M. Nitrogen, phosphorus, and eutrophication in the coastal marine environment. Science, 1971, 171(3975): 1008–1013. DOI:10.1126/science.171.3975.1008 |
| [11] | Gusewell S. N:P ratios in terrestrial plants:variation and functional significance. New Phytologist, 2004, 164(2): 243–266. DOI:10.1111/j.1469-8137.2004.01192.x |
| [12] | McGill B J, Enquist B J, Weiher E, Westoby M. Rebuilding community ecology from functional traits. Trends in Ecology & Evolution, 2006, 21(4): 178–185. |
| [13] | Croft H, Chen J M, Luo X Z, Bartlett P, Chen B, Staebler R M. Leaf chlorophyll content as a proxy for leaf photosynthetic capacity. Global Chang Biology, 2017, 23(9): 3513–3524. DOI:10.1111/gcb.2017.23.issue-9 |
| [14] | Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets V, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821–827. DOI:10.1038/nature02403 |
| [15] | 张育新, 马克明, 祁建, 冯云, 张洁瑜. 北京东灵山海拔梯度上辽东栎种群结构和空间分布. 生态学报, 2009, 29(6): 2789–2796. DOI:10.3321/j.issn:1000-0933.2009.06.004 |
| [16] | Li N N, He N P, Yu G R, Wang Q F, Sun J. Leaf non-structural carbohydrates regulated by plant functional groups and climate:Evidences from a tropical to cold-temperate forest transect. Ecological Indicators, 2016, 62: 22–31. DOI:10.1016/j.ecolind.2015.11.017 |
| [17] | Wood S A, Karp D S, DeClerck F, Kremen C, Naeem S, Palm C A. Functional traits in agriculture:agrobiodiversity and ecosystem services. Trends in Ecology & Evolution, 2015, 30(9): 531–539. |
| [18] | 罗璐, 申国珍, 谢宗强, 喻杰. 神农架海拔梯度上4种典型森林的乔木叶片功能性状特征. 生态学报, 2011, 31(21): 6420–6428. |
| [19] | Wang R L, Yu G R, He N P, Wang Q F, Zhao N, Xu Z W. Latitudinal variation of leaf morphological traits from species to communities along a forest transect in eastern China. Journal of Geographical Sciences, 2016, 26(1): 15–26. DOI:10.1007/s11442-016-1251-x |
| [20] | da Silveira T I, Boeger M R T, Maranho L T, De Melo Jr J C F, Soffiatti P. Functional leaf traits of 57 woody species of the Araucaria forest, Southern Brazil. Brazilian Journal of Botany, 2015, 38(2): 357–366. DOI:10.1007/s40415-014-0111-0 |
| [21] | Osnas J L D, Lichstein J W, Reich P B, Pacala S W. Global leaf trait relationships:mass, area, and the leaf economics spectrum. Science, 2013, 340(6133): 741–744. DOI:10.1126/science.1231574 |
| [22] | 杨冬梅, 章佳佳, 周丹, 钱敏杰, 郑瑶, 金灵妙. 木本植物茎叶功能性状及其关系随环境变化的研究进展. 生态学杂志, 2012, 31(3): 702–713. |
| [23] | Kunstler G, Falster D, Coomes D A, Hui F, Kooyman R M, Laughlin D C, Poorter L, Vanderwel M, Vieilledent G, Wright S J, Aiba M, Baraloto C, Caspersen J, Cornelissen J H C, Gourlet-Fleury S, Hanewinkel M, Herault B, Kattge J, Kurokawa H, Onoda Y, Peñuelas J, Poorter H, Uriarte M, Richardson S, Ruiz-Benito P, Sun I F, Ståhl G, Swenson N G, Thompson J, Westerlund B, Wirth C, Zavala M A, Zeng H C, Zimmerman J K, Zimmermann N E, Westoby M. Plant functional traits have globally consistent effects on competition. Nature, 2015, 529(7585): 204–207. |
| [24] | Rozendaal D M A, Hurtado V H, Poorter L. Plasticity in leaf traits of 38 tropical tree species in response to light; relationships with light demand and adult stature. Functional Ecology, 2006, 20(2): 207–216. DOI:10.1111/fec.2006.20.issue-2 |
| [25] | Ruger N, Wirth C, Wright S J, Condit R. Functional traits explain light and size response of growth rates in tropical tree species. Ecology, 2012, 93(12): 2626–2636. DOI:10.1890/12-0622.1 |
| [26] | Liu C C, He N P, Zhang J H, Li Y, Wang Q F, Sack L, Yu G R, Niu S L. Variation of stomatal traits from cold temperate to tropical forests and association with water use efficiency. Functional Ecology, 2017, 32(1): 20–28. |
| [27] | Niklas K J, Owens T, Reich P B, Cobb E D. Nitrogen/phosphorus leaf stoichiometry and the scaling of plant growth. Ecology Letters, 2005, 8(6): 636–642. DOI:10.1111/ele.2008.8.issue-6 |
| [28] | Lienin P, Kleyer M. Plant trait responses to the environment and effects on ecosystem properties. Basic and Applied Ecology, 2012, 13(4): 301–311. DOI:10.1016/j.baae.2012.05.002 |
| [29] | Kröber W, Plath I, Heklau H, Bruelheide H. Relating stomatal conductance to leaf functional traits. Journal of Visualized Experiments, 2015(104): 52738. |
| [30] | 李玉霖, 崔建垣, 苏永中. 不同沙丘生境主要植物比叶面积和叶干物质含量的比较. 生态学报, 2005, 25(2): 304–311. |
| [31] | Bullock D G, Anderson D S. Evaluation of the Minolta SPAD-502 chlorophyll meter for nitrogen management in corn. Journal of Plant Nutrition, 1998, 21(4): 741–755. DOI:10.1080/01904169809365439 |
| [32] | Li Y, Liu C C, Zhang J H, Yang H, Xu L, Wang Q F, Sack L, Wu X Q, Hou J H, He N P. Variation in leaf chlorophyll concentration from tropical to cold-temperate forests:association with gross primary productivity. Ecological Indicators, 2018, 85: 383–389. DOI:10.1016/j.ecolind.2017.10.025 |
| [33] | Reich P B, Walters M B, Ellsworth D S, Uhl C. Photosynthesis-nitrogen relations in Amazonian tree species. Oecologia, 1994, 97(1): 62–72. DOI:10.1007/BF00317909 |
| [34] | 黄菊莹, 袁志友, 李凌浩. 羊草绿叶氮、磷浓度和比叶面积沿氮、磷和水分梯度的变化. 植物生态学报, 2009, 33(3): 442–448. DOI:10.3773/j.issn.1005-264x.2009.03.003 |
| [35] | Chen Z H, Zha T S, Jia X, Wu Y J, Wu B, Zhang Y Q, Guo J B, Qin S G, Chen G P, Peltola H. Leaf nitrogen is closely coupled to phenophases in a desert shrub ecosystem in China. Journal of Arid Environments, 2015, 122: 124–131. DOI:10.1016/j.jaridenv.2015.07.002 |
| [36] | Reich P B, Ellsworth D S, Walters M B. Leaf structure (specific leaf area) modulates photosynthesis-nitrogen relations:evidence from within and across species and functional groups. Functional Ecology, 1998, 12(6): 948–958. DOI:10.1046/j.1365-2435.1998.00274.x |
| [37] | 李轩然, 刘琪璟, 蔡哲, 马泽清. 千烟洲针叶林的比叶面积及叶面积指数. 植物生态学报, 2007, 31(1): 93–101. DOI:10.3321/j.issn:1005-264X.2007.01.012 |
| [38] | Goedhart C M, Pataki D E, Billings S A. Seasonal variations in plant nitrogen relations and photosynthesis along a grassland to shrubland gradient in Owens Valley, California. Plant and Soil, 2010, 327(1/2): 213–223. |
| [39] | Wilson P J, Thompson K, Hodgson J G. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytologist, 1999, 143(1): 155–162. DOI:10.1046/j.1469-8137.1999.00427.x |
| [40] | Lima J D, Mosquim P R, da Matta F M. Leaf gas exchange and chlorophyll fluorescence parameters in Phaseolus vulgaris as affected by nitrogen and phosphorus deficiency. Photosynthetica, 1999, 37(1): 113–121. DOI:10.1023/A:1007079215683 |
| [41] | Vile D, Garnier É, Shipley B, Laurent G, Navas M L, Roumet C, Lavorel S, Diaz S, Hodgson J G, Lloret F, Midgley G F, Poorter H, Rutherford M C, Wilson P J, Wright I J. Specific leaf area and dry matter content estimate thickness in laminar leaves. Annals of Botany, 2005, 96(6): 1129–1136. DOI:10.1093/aob/mci264 |