 2018, Vol. 38
2018, Vol. 38文章信息
- 马天光, 李向义, 林丽莎, 李磊, 李世民, 王波.
- MA Tianguang, LI Xiangyi, LIN Lisha, LI Lei, LI Shimin, WANG Bo.
- 遮阴对骆驼刺叶性状和水分生理的影响
- The effects of shade on leaf traits and water physiological characteristics in Alhagi sparsifolia
- 生态学报. 2018, 38(23): 8466-8474
- Acta Ecologica Sinica. 2018, 38(23): 8466-8474
- http://dx.doi.org/10.5846/stxb201711252105
-
文章历史
- 收稿日期: 2017-11-25
- 网络出版日期: 2018-09-26





2. 新疆策勒荒漠草地生态系统国家野外科学观测研究站, 策勒 848300;
3. 中国科学院干旱区地理与生物资源重点实验室, 乌鲁木齐 830011;
4. 中国科学院大学, 北京 100049
2. Cele National Station of Observation and Research for Desert-Grassland Ecosystems, Cele 848300, China;
3. Key Laboratory of Biogeography and Bioresearch in Arid Zone, Chinese Academy of Sciences, Urumqi 830011, China;
4. University of Chinese Academy of Sciences, Beijing 100049, China
光是影响植物生存和生长的重要环境因子之一, 是植物生长的能量来源[1]。遮阴环境主要减少了植物光合有效辐射, 进而会明显影响植物的光合作用和光形态建成[2]。一般认为, 随着遮阴程度的加强, 植物通过增加单叶面积和比叶面积, 减小叶片厚度和比叶质量等来适应光环境的变化[3];植物叶片光合生理通过增加光合色素含量, 尤其增加叶绿素b的含量[4], 降低光饱和点、光补偿点和暗呼吸速率, 增大光能利用效率等来适应光环境的减弱[5-6];植物的生物量分配也会通过增加叶干重, 减小根冠比等来适应光环境的减弱[7]。然而, 目前遮阴环境对干旱区荒漠植物生理生态影响的研究多见于叶形态特征、生物量分配和光合生理等方面, 对植物水分生理响应以及叶性状变化与水分生理关系的研究少见报道。干旱区荒漠植物的水分生理(组织相对含水量、失水速率、水势、蒸腾速率、水分利用效率、PV水分参数等)对环境的响应的研究多集中于干旱和水分胁迫环境下植物的生理生态响应以及植物抗旱性评价等方面[8-10]。然而, 不同光强环境也会影响干旱区荒漠植物的水分生理。如弱光环境影响植株叶性状适应弱光的同时也增大了植株叶片的失水速率, 降低了蒸腾速率和水分利用效率等水分生理参数, 进而影响植物的抗旱性[11-12]。这说明光环境的改变在影响植物光合作用和光形态建成的同时也影响了植物的水分生理。本研究集中探讨干旱区荒漠植物水分生理对遮阴环境的响应特征以及遮阴条件下叶性状变化与水分生理的关系。
干旱区荒漠植物通常遭受环境的多重胁迫(干旱、高温、强光、风沙等), 研究极端环境条件下植物对生境的适应机制和策略, 对于退化植被修复具有重要意义。骆驼刺(Alhagi sparsifolia Shap.)是塔克拉玛干沙漠南缘沙漠绿洲过渡带地区重要的植被群落, 对防风固沙、维护绿洲脆弱的生态环境具有重要作用[13]。因此有关骆驼刺在塔克拉玛干沙漠南缘极端强光、干旱等气候下生理生态特征的研究历来受到人们的关注[14]。目前认为疏叶骆驼刺为阳生植物, 但调查骆驼刺生境发现, 当其他条件一致, 随着生境遮阴程度的增加(100%自然光—20%自然光), 骆驼刺的形态和生理发生显著变化, 当遮阴增加到一定程度时骆驼刺植株就极少出现, 这说明遮阴环境对骆驼刺的形态和生理生态产生了显著的影响并且随着遮阴程度的增加会影响骆驼刺植被群落的分布与演替。前人对骆驼刺在不同遮阴环境下生理生态响应的研究多集中于叶形态、生物量分配、光合生理等方面[15-16];对骆驼刺水分生理特征的研究多关注于不同地下水埋深、土壤水分和灌溉等环境以及自然环境中多种荒漠植物水分关系的比较等[17-19];然而, 不同遮阴环境下骆驼刺叶性状和水分生理的响应特征以及它们之间的相关性少有研究[20]。因此本试验以极端干旱区自然环境中生长的骆驼刺为研究对象, 研究了遮阴环境对骆驼刺叶性状和水分生理的影响, 以阐明骆驼刺在叶性状和水分生理上对不同光环境的响应特征, 为干旱区荒漠植物响应光环境胁迫的内在机理提供基础理论研究; 探讨骆驼刺在何种光环境下的叶性状和水分生理特征更适合干旱环境, 并试图确定更适合骆驼刺生存的遮阴强度, 进而为骆驼刺植被修复和生态保护以及种群扩大提供科学依据。
1 材料与方法 1.1 试验区域概况研究区位于塔克拉玛干沙漠南缘策勒绿洲外围的沙漠绿洲过渡带间(经纬度:35°17′55″—39°30′00″N, 80°03′24″—82°10′34″E, 海拔1360 m)。试验样地设置在中国科学院策勒荒漠草地生态系统国家野外研究站自然植被观测场内。该研究区属典型内陆暖温带荒漠气候。夏季炎热, 干旱少雨, 光热充足, 日照时间长, 昼夜温差大; 年均日照数2697 h, 年太阳总辐射能604.2 kJ/cm2, 仅次于青藏高原。多年平均降水量35.1 mm, 5月和7月的降雨次数相对较多, 但降雨量很少, 年平均蒸发量高达2595.3 mm, 水分亏值较大。研究区地下水埋深为13.5 m, 植物的水分来源主要是地下水[17]。骆驼刺在沙漠绿洲过渡带广泛分布, 是当地的主要建群种和优势种。
1.2 试验设计试验材料为沙漠绿洲过渡带自然生长的骆驼刺植株。于2017年4月底, 在中国科学院策勒荒漠草地生态系统国家野外研究站自然植被观测场内进行试验。对生长良好的骆驼刺植株进行遮光处理。遮光处理前骆驼刺平均株高在20 cm左右, 并且呈现一定簇状。试验利用黑色尼龙遮阳网进行遮光处理, 根据实验地附近骆驼刺生境光照强度的变化范围(100%—20%自然光)设置中度遮阴(40%自然光)、重度遮阴(20%自然光)和自然光照3个处理。每个遮阴棚面积为3 m×6 m, 高2 m, 每个处理4个重复。遮阴棚东、南、西三面遮网, 四周垂下部分距离地面80 cm高度不进行遮阴(确保遮阴棚内外空气流动自由), 遮阴棚内边缘0.5 m内生长的骆驼刺为缓冲植株。遮阴处理60 d, 于7月初对骆驼刺叶性状、相对含水量、失水速率、水势、光合参数和水分利用效率等进行测定。
1.3 测定项目及方法 1.3.1 叶性状参数的测定于2017年7月初, 在每个处理随机选取足够量骆驼刺的健康无损坏叶片并进行均匀混合, 在低温、遮光条件下带回实验室立即进行各叶性状参数测定。从采集的不同处理的叶片中各随机选取10枚叶片用精度为0.01 mm的游标卡尺进行叶厚测量。同时从采集的叶片中随机选取10枚叶片进行叶面积扫描, 通过Image-Pro Plus软件计算叶面积, 然后放入烘箱105℃杀青30 min, 最后80℃烘干至恒重。根据以下公式计算叶片的比叶面积(SLA)和比叶质量(LMA):SLA =叶片面积(cm2)/叶片干重(g); LMA=叶片干重(g)/叶片面积(m2)。每个指标每个处理都设10个重复。
1.3.2 叶相对含水量和失水速率的测定于2017年7月初, 在每个处理随机选取足够量骆驼刺的健康无损坏叶片并进行均匀混合, 在低温、遮光条件下带回实验室立即进行叶相对含水量和失水速率测定。
从采集的不同处理的叶片中各随机选取10枚叶片用精度为0.0001 g的分析天平进行称重得鲜重W1, 然后将叶片浸泡在装有蒸馏水的烧杯里用自封袋密封放置在黑暗处使叶片进行饱和吸水24 h, 然后取出饱和吸水的叶片, 用滤纸吸掉叶片表面多余的水分后立即进行称重得饱和鲜重W2, 然后将叶片放入烘箱105 ℃杀青30 min, 最后80 ℃烘干至恒重得叶片干重W3。每个处理都设10个重复。根据以下公式计算叶片相对量(RWC):
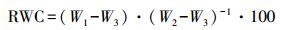
|
同时从采集的不同处理的叶片中各随机选取10枚叶片用精度为0.0001 g的分析天平进行称重得鲜重W1(记此时称重时间为t0), 然后将叶片放在25 ℃的实验室内让其进行自然失水12 h, 每1 h称重一次得实时鲜重Wi (记每次称重时间为ti), 然后将叶片放入烘箱105 ℃杀青30 min, 最后80 ℃烘干至恒重得叶片干重W2。根据以下公式计算每个失水时间点的叶片失水速率(RWL):
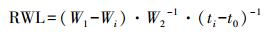
|
然后用每个失水时间点和对应的失水速率做叶片自然失水12 h的失水率速变化曲线。选取不同处理叶片失水速率差异最大时的失水速率作为各处理的叶片失水速率。每个处理都设10个重复。
1.3.3 水势的测定于2017年7月初, 选择晴朗无云的天气, 在每块测定样区内选择6株生长状况良好的样株。在样株的中上部分选择1个二级分枝, 用PMS压力室(Corvallis, Oregon, USA)进行带叶小枝的水势测定。于6:30(北京时间, 下同)测定其清晨水势, 14:00测定其正午水势。每个处理选取3个重复样区。
1.3.4 光合参数和水分利用效率的测定于2017年7月初, 选择晴朗无云天气, 在每块测定样区内选择2株生长状况良好的样株。在样株的中上部的二级分枝上选择1片健康无损坏叶片, 于12:00和14:00用LI-6400光合仪(LI-COR, Lincoln, USA)进行光合参数测定。测定完之后将叶片低温带回实验室进行叶面积扫描, 用Image-Pro Plus软件计算出叶面积后替换光合仪默认叶室面积得到叶片的净光合速率(Net photosynthetic rate, Pn)、气孔导度(Stomatal conductance, Gs)、胞间二氧化碳浓度(Intercellular CO2 concentration, Ci)和蒸腾速率(Transpiration rate, Tr)。然后根据以下公式计算出叶片水分利用效率WUE。WUE=净光合速率Pn/蒸腾速率Tr。每个处理选取3个重复样区。
1.4 数据处理前期利用Microsoft Excel 2007对数据进行整理计算。然后采用SPSS 20.0统计软件分析各指标参数不同处理之间的差异显著性, 不同变量之间的相关性分析采用Pearson相关法进行。P < 0.05为差异显著; 参数以平均值±均值标准误表示。最后采用Origin 9.0软件对各参数数据进行绘图。
2 结果与分析 2.1 遮阴对骆驼刺叶性状的影响如表 1所示, 与正常光照相比, 中度遮阴条件下骆驼刺叶厚和比叶质量分别降低8.45%和14.37%, 单叶面积和比叶面积分别增加17.80%和16.84%(P < 0.05);重度遮阴条件下骆驼刺叶厚和比叶质量分别降低14.16%和31.67%, 单叶面积和比叶面积分别增加32.40%和46.33%(P < 0.05), 其中重度遮阴变化更显著。
| 参数 Parameter |
处理Treatment | ||
| NL | MS | SS | |
| 叶厚LT/mm | 0.438 ± 0.003a | 0.401 ± 0.003b | 0.376 ± 0.002c |
| 单叶面积LA/cm2 | 1.534 ± 0.069a | 1.807 ± 0.057b | 2.031 ± 0.096c |
| 比叶面积SLA/(cm2/g) | 92.278 ± 1.812a | 107.819 ± 2.450b | 135.033 ± 2.582c |
| 比叶质量LMA/(g/m2) | 108.543 ± 2.224a | 92.942 ± 2.137b | 74.166 ± 1.435c |
| NL, 正常光照Normal light; MS, 中度遮阴Moderate shade; SS, 重度遮阴Severe shade; LT, Leaf thickness; LA, Single leaf area; SLA, Specific leaf area; LMA, Leaf mass per unit area; 不同字母表示不同处理之间差异显著(P < 0.05) | |||
由图 1可见, 遮阴条件下骆驼刺清晨水势、正午水势和叶片相对含水量都明显高于正常光照。与正常光照相比, 中度遮阴条件下骆驼刺清晨水势、正午水势和叶片相对含水量分别升高28.3%、11.43%和15.69%(P < 0.05), 重度遮阴条件下清晨水势、正午水势和叶片相对含水量分别升高43.4%、20.48%和14.92%(P < 0.05)。其次, 对于骆驼刺水势和叶片相对含水量, 不同遮阴条件下清晨水势和叶片相对含水量的变化具有较高的一致性, 正午水势变化更显著。

|
| 图 1 不同遮阴条件下骆驼刺水势和叶片相对含水量的变化(均值±标准误) Fig. 1 Changes of water potential and relative water content in A. sparsifolia under different shade treatment (mean±SE) NL, 正常光照Normal light; MS, 中度遮阴Moderate shade; SS, 重度遮阴Severe shade; 不同字母表示不同处理之间差异显著(P < 0.05) |
由图 2可见, 正常光照下, 骆驼刺失水速率较低, 失水过程变化趋势平稳; 遮阴条件下失水速率较高, 变化幅度较大, 其中重度遮阴表现更明显。与正常光照相比, 中度遮阴和重度遮阴条件下骆驼刺失水率速分别增加85.21%和124.38%(P < 0.05)。

|
| 图 2 不同遮阴条件下骆驼刺叶片失水速率失水过程的变化特征(均值±标准误) Fig. 2 Change characteristics of leaf water loss rate in A. sparsifolia under different shade treatment (mean±SE) |
由图 3可见, 不同遮阴条件下骆驼刺净光合速率Pn、气孔导度Gs、蒸腾速率Tr和水分利用效率WUE都显著低于自然光照, 12:00差异更明显(P < 0.05), 变化幅度较大。以12:00为例, 与正常光照相比, 中度遮阴条件下骆驼刺Pn、Gs、Tr和WUE分别增大68.89%、45.83%、33.94%和49.41%(P < 0.05), 重度遮阴条件下Pn、Gs、Tr和WUE分别增大90.03%、66.67%、58.82%和74.12%(P < 0.05)。其次, 12:00时, 不同遮阴条件下骆驼刺Pn、Gs、Tr和WUE的变化均具有较好的一致性; 而14:00时, Pn、Gs和Tr变化一致, 但与WUE变化呈负相关(图 3)。比较12:00和14:00各处理骆驼刺的WUE, 14:00均低于12:00, 并且自然光照下变化幅度相对最大, 中度遮阴次之, 重度遮阴最小(图 3)。

|
| 图 3 不同遮阴条件下骆驼刺蒸腾速率和水分利用效率的变化(均值±标准误) Fig. 3 Changes of transpiration rate and water use efficiency in A. sparsifolia under different shade treatment (mean±SE) 不同字母表示同一时间不同处理之间差异显著, *表示同一处理在不同时间差异显著(P < 0.05) |
由表 2可见, 遮阴条件下叶片各结构型性状参数的变化都与各水分生理参数的变化有着极显著的相关性(P < 0.01)。叶厚、比叶质量与叶片相对含水量、失水速率呈极显著负相关, 与清晨水势、正午水势、蒸腾速率、水分利用效率呈极显著正相关; 而单叶面积、比叶面积与叶片相对含水量、失水速率呈极显著正相关, 与清晨水势、正午水势、蒸腾速率、水分利用效率呈极显著负相关。比较相关系数, 叶厚、比叶面积、比叶质量与失水速率、正午水势、蒸腾速率、水分利用效率相关性更好, 单叶面积与失水速率、清晨水势相关性更好。
| 参数 Parameter |
叶厚 Leaf thickness |
单叶面积 Single leaf area |
比叶面积 Specific leaf area |
比叶质量 Leaf mass per unit area |
| 相对含水量Relative water content | -0.699** | 0.592** | 0.609** | -0.662** |
| 失水速率Water loss rate | -0.899** | 0.801** | 0.890** | -0.907** |
| 清晨水势Predawn water potential | 0.784** | -0.788** | -0.817** | 0.821** |
| 正午水势Noon water potential | 0.946** | -0.740** | -0.944** | 0.953** |
| 蒸腾速率Transpiration rate | 0.962** | -0.768** | -0.930** | 0.951** |
| 水分利用效率Water use efficiency | 0.966** | -0.767** | -0.912** | 0.938** |
| **极显著性相关(P < 0.01) | ||||
植物叶性状与植物的生长对策和植物利用资源的能力密切相关, 而且直接影响到植物的基本行为和功能[21]。因此研究叶性状的变化对深入了解植物适应环境具有重要意义。叶结构型性状是植物叶片的生物化学结构特征的基础, 能够客观表达植物对外部环境的适应性[22]。叶片是植物光合作用和散失水分的主要器官, 生活在干旱区的荒漠植物通过叶厚、单叶面积、比叶面积等叶性状变异减小受光面积和水分散失, 这是植物适应强光及干旱环境的重要特征。有研究表明, 叶片厚度与光照强度成正相关, 单叶面积与光照强度成负相关。强光环境下叶片会通过增大叶片厚度减小单叶面积以减少光辐射的穿透量, 从而避免强光对叶片的伤害[23-24]。弱光环境下植物叶片倾向于增加共质体的投入减少叶片保护性结构投入, 减小叶片厚度增大单叶面积, 这一方面有利于叶片尽可能多的接收有限的光辐射, 另一方面有利于有限的光辐射穿透叶表皮到达叶肉组织或直接在叶表皮中进行光化学反应, 从而增强叶片细胞对光的捕获能力[25]。本试验结果显示, 生活在不同遮阴环境下的骆驼刺, 其叶性状在变化性质上表现为叶片厚度和比叶质量显著变小, 单叶面积和比叶面积显著变大, 在变化数量上表现为单叶面积、比叶面积和比叶质量变化较大, 叶厚变化较小(表 1)。结果表明, 与正常光照相比, 遮阴环境下骆驼刺叶厚和比叶质量变小单叶面积和比叶面积变大, 明显增大受光面积减小光辐射穿透距离以便增强叶片细胞对弱光辐射的捕获能力从而提高光能利用效率尽可能保证一定的光合能力。不同遮阴环境下骆驼刺单叶面积、比叶面积和比叶质量变化较大, 叶厚变化较小, 这说明骆驼刺叶性状主要通过单叶面积和单叶干物质积累的变化响应光环境的变化。其次, 遮阴环境下骆驼刺叶性状的变化也明显增大了叶片的失水面积和失水阻力降低了其叶片的保水力。
3.2 遮阴对骆驼刺水分生理的影响植物叶片的保水力是反映植物抗旱能力的一个重要指标, 保水力强的荒漠植物能够在极端干旱区维持正常的生命活动。叶片相对含水量和失水速率是衡量植物叶片保水力的常用指标。水势是反映植物水分状况或水分亏缺程度的一个直接判定指标。其中, 清晨水势可以反映植物水分平衡的恢复状况; 正午水势则表示在当日所受最大环境胁迫时的程度状况[26]。有研究表明, 随着干旱胁迫的加剧, 植物叶片相对含水量和水势下降幅度加快[27];植物拥有较低的叶片失水速率是一种抗旱生理机制类型[12]。也有研究表明, 植物叶片水势和遮阴程度有着较好的相关性[28]。本试验结果显示, 遮阴环境下骆驼刺叶片相对含水量、水势和失水速率都显著高于自然光照(图 1和图 2), 其中, 叶片失水速率和水势变化相对较大, 清晨水势与叶片相对含水量变化一致性较好, 正午水势与叶片失水速率变化一致性较好。结果表明, 与正常光照相比, 遮阴环境下骆驼刺叶片相对含水量显著变大说明遮阴环境明显降低叶温和蒸腾速率(图 3)进而减小叶片水分消耗改善骆驼刺的干旱胁迫程度。遮阴环境下骆驼刺清晨水势和正午水势显著变大说明遮阴环境明显缓解了骆驼刺正午所受的强光和干旱胁迫程度, 提升了植株的水分亏缺恢复状况; 也说明自然光照下的骆驼刺确实存在着强光胁迫, 其植被修复需要一定程度的遮阴; 其清晨水势与叶片相对含水量变化一致性较好也说明高叶片含水量和低蒸腾速率使得骆驼刺不需要通过维持较低的渗透压以提升吸水能力。遮阴环境下骆驼刺叶片失水速率及其变化幅度相对较大说明其水分生理中叶片失水速率受生境光强变化影响较大, 遮阴环境明显降低了骆驼刺叶片的保水力。正午水势与叶片失水速率变化一致性较好一方面说明自然光照下骆驼刺存在强光胁迫生境, 其叶片表现低且平稳的失水速率适应强光胁迫; 另一方面说明遮阴环境虽然缓解了强光胁迫增大了骆驼刺叶片含水量和水势减小了蒸腾速率, 但同时也使其高的叶片保水能力出现大幅度退化。
另外, 从植物的水分消耗的变化也可以进一步阐释遮阴环境对骆驼刺水分生理的影响。蒸腾速率作为一个重要的水分指标, 能够反映植物潜在的耗水能力。水分利用效率WUE是衡量叶片碳固定与水分消耗比例的良好指标, 是植物自身的水分利用状况和和抗旱性的一个客观综合评价参数[9]。本实验结果显示, 遮阴环境下骆驼刺Pn、Gs、Tr和WUE都显著低于正常光照。其中, 12:00时, Pn、Gs、Tr和WUE变化均有较好一致性; 正午光强最大时, Pn、Gs和Tr变化一致性较好, 但它们与WUE变化呈负相关(图 3)。结果表明, 与正常光照相比, 遮阴环境下骆驼刺明显受有限光强的限制, 其主要靠气孔调节和更多的水分消耗用于提高光能利用效率维持其一定的光合速率进而呈现较低的WUE[16]。本试验结果中, Pn和Gs的变化特征表明自然光照下骆驼刺存在着强光胁迫, 同时中度遮阴也存在着弱光胁迫; 叶片失水速率和WUE的变化特征表明大程度遮阴环境可能会使骆驼刺的抗旱能力出现一定程度的退化[29]。
3.3 遮阴环境下骆驼刺叶性状变化与水分生理变化的关系有研究表明, 植物水分生理状况与叶片性状有着密切的联系[30]。本实验对遮阴条件下骆驼刺叶片各结构型性状参数与水分生理参数做了相关性分析, 结果发现遮阴环境下骆驼刺叶片结构型性状的变化对其水分生理状况的变化有着极显著的相关性(表 2)。研究表明, 强光和干旱环境下植物叶片厚度增加单叶面积减小, 增加水分在叶片组织中的传导距离减小水分散失面积, 从而减少组织内水分损失; 比叶质量的增加是提高叶片水分利用效率的重要途径[9]。从本试验结果看, 遮阴环境下骆驼刺叶厚和比叶质量变小单叶面积和比叶面积变大, 明显增大水分散失面积减小失水阻力进而显著影响其失水速率和水分利用效率。本实验结果中, 不同遮阴环境下骆驼刺叶性状的变化特征以及与水分生理参数极显著的相关性说明自然光照其叶性状表现为阳生和旱生特点, 使其水分生理参数更多呈现出对由强光胁迫带来的水分胁迫的响应特征; 遮阴环境下骆驼刺的叶性状特征使其水分生理参数更多呈现出对弱光胁迫的响应特征甚至牺牲对水分胁迫有较好抗逆能力的水分生理特征。比较叶性状参数与水分生理参数的相关系数的结果说明遮阴环境下骆驼刺叶性状的变化主要通过影响失水速率、水势、蒸腾速率以及水分利用效率进而影响其水分生理的变化特征。结果表明不同遮阴环境下骆驼刺叶片结构型性状与水分生理各参数之间相互制约、相互影响, 是影响其水分生理参数变化的主要因素。
综上所述, 遮阴环境显著影响了骆驼刺叶性状和水分生理状况。遮阴环境下:骆驼刺叶性状变化特征表现为叶厚和比叶质量显著变小, 单叶面积和比叶面积显著变大; 与其他干旱区荒漠植物相比, 骆驼刺主要通过单叶面积和单叶干物质积累的变化来响应遮阴环境。骆驼刺水分生理变化特征表现为叶片相对含水量、水势、失水速率变大, 气孔导度、蒸腾速率和水分利用效率变小; 但其变化特征主要是牺牲对水分胁迫有较好抗逆性的水分生理特征并通过气孔调节和更多的水分消耗用于维持一定的光合能力来响应遮阴环境。从叶性状与水分生理参数的相关性来看, 遮阴环境下骆驼刺叶性状变化与水分生理参数之间相互制约、相互影响, 是影响其水分生理参数变化的主要因素; 但主要是通过影响失水速率、水势、蒸腾速率以及水分利用效率进而影响其水分生理的变化特征。综合分析本试验结果可知, 自然光照下骆驼刺确实存在强光胁迫, 中度遮阴环境下骆驼刺也确实存在明显的弱光胁迫, 重度遮阴环境下弱光胁迫更严重, 其环境下的骆驼刺甚至开始出现植株死亡现象。因此, 本研究建议可利用遮阴措施对极端干旱区荒漠植物骆驼刺进行植被恢复, 但其遮光度应设置在70%—80%自然光之间(自然光照下骆驼刺光饱和点时的光辐射强度约为最大光辐射强度的70%—80%), 此遮光度环境可能实现既缓解了极端干旱区骆驼刺的强光和干旱胁迫, 又能保持其较好的保水能力以及抗旱能力。
致谢: 感谢李成道和岳泽伟等对试验的帮助。| [1] | 刘建锋, 杨文娟, 江泽平, 郭泉水, 金江群, 薛亮. 遮荫对濒危植物崖柏光合作用和叶绿素荧光参数的影响. 生态学报, 2011, 31(20): 5999–6004. |
| [2] | Sofo A, Dichio B, Montanaro G, Xiloyannis C. Shade effect on photosynthesis and photoinhibition in olive during drought and rewatering. Agricultural Water Management, 2009, 96(8): 1201–1206. DOI:10.1016/j.agwat.2009.03.004 |
| [3] | Dai Y J, Shen Z G, Ying L, Wang L L, Hannaway D, Lu H F. Effects of shade treatments on the photosynthetic capacity, chlorophyll fluorescence, and chlorophyll content of Tetrastigma hemsleyanum Diels et Gilg. Environmental and Experimental Botany, 2009, 65(2/3): 177–182. |
| [4] | Sarijeva G, Knapp M, Lichtenthaler H K. Differences in photosynthetic activity, chlorophyll and carotenoid levels, and in chlorophyll fluorescence parameters in green sun and shade leaves of Ginkgo and Fagus. Journal of Plant Physiology, 2007, 164(7): 950–955. DOI:10.1016/j.jplph.2006.09.002 |
| [5] | 张林, 罗天祥. 植物叶寿命及其相关叶性状的生态学研究进展. 植物生态学报, 2004, 28(6): 844–852. DOI:10.3321/j.issn:1005-264X.2004.06.014 |
| [6] | 秦舒浩, 李玲玲. 遮光处理对西葫芦幼苗形态特征及光合生理特性的影响. 应用生态学报, 2006, 17(4): 653–656. DOI:10.3321/j.issn:1001-9332.2006.04.019 |
| [7] | 王蕊, 孙备, 李建东, 王国骄, 孙佳楠, 王欣若, 钟日亭. 不同光强对入侵种三裂叶豚草表型可塑性的影响. 应用生态学报, 2012, 23(7): 1797–1802. |
| [8] | 涂璟, 王克勤. 干旱地区造林树种的水分生理生态的研究进展. 西北林学院学报, 2003, 18(3): 26–30. DOI:10.3969/j.issn.1001-7461.2003.03.008 |
| [9] | 张硕, 刘勇, 李国雷, 陈晓, 孙巧玉, 许飞. 稳定碳同位素在森林植物水分利用效率研究中的应用. 世界林业研究, 2013, 26(3): 39–45. |
| [10] | 刘建平, 李志军, 韩路, 龚卫江, 周正立. 胡杨、灰叶胡杨P-V曲线水分参数的初步研究. 西北植物学报, 2004, 24(7): 1255–1259. DOI:10.3321/j.issn:1000-4025.2004.07.019 |
| [11] | 蔡海霞, 吴福忠, 杨万勤. 干旱胁迫对高山柳和沙棘幼苗光合生理特征的影响. 生态学报, 2011, 31(9): 2430–2436. |
| [12] | Liu F, Stützel H. Biomass partitioning, specific leaf area, and water use efficiency of vegetable amaranth (Amaranthus spp.) in response to drought stress. Scientia Horticulturae, 2004, 102(1): 15–27. DOI:10.1016/j.scienta.2003.11.014 |
| [13] | 张晓蕾, 曾凡江, 刘波, 张慧, 刘镇, 安桂香. 不同地下水埋深下骆驼刺幼苗叶片生理参数光响应特性. 干旱区地理, 2011, 34(2): 229–235. |
| [14] | 罗维成, 曾凡江, 刘波, 张利刚, 宋聪, 彭守兰, Arndt S K. 疏叶骆驼刺根系对土壤异质性和种间竞争的响应. 植物生态学报, 2012, 36(10): 1015–1023. |
| [15] | 李磊.温度及光强对疏叶骆驼刺表型可塑性及光系统Ⅱ活性的影响[D].乌鲁木齐: 中国科学院新疆生态与地理研究所, 2015. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y2958631 |
| [16] | 薛伟, 李向义, 朱军涛, 林丽莎, 王迎菊. 遮阴对疏叶骆驼刺叶形态和光合参数的影响. 植物生态学报, 2011, 35(1): 82–90. |
| [17] | 张晓蕾, 曾凡江, 刘波, 刘镇, 安桂香, 孙旭伟. 不同土壤水分处理对疏叶骆驼刺幼苗光合特性及干物质积累的影响. 干旱区研究, 2010, 27(4): 649–655. |
| [18] | 朱军涛, 李向义, 张希明, 曾凡江, 闫海龙, 杨尚功. 灌溉对疏叶骆驼刺(Alhagi sparsifolia)幼苗光合生理指标及渗透物质的影响. 中国沙漠, 2009, 29(4): 697–702. |
| [19] | 李向义, 张希明, 何兴元, 曾凡江, Foetzki A, Thomas F M. 沙漠-绿洲过渡带四种多年生植物水分关系特征. 生态学报, 2004, 24(6): 1164–1171. DOI:10.3321/j.issn:1000-0933.2004.06.011 |
| [20] | 唐钢梁, 李向义, 林丽莎, 李磊, 鲁建荣. 骆驼刺在不同遮阴下的水分状况变化及其生理响应. 植物生态学报, 2013, 37(4): 354–364. |
| [21] | Reich P B, Buschena C, Tjoelker M G, Wrage K, Knops J, Tilman D, Machado J L. Variation in growth rate and ecophysiology among 34 grassland and savanna species under contrasting N supply:a test of functional group differences. New Phytologist, 2010, 157(3): 617–631. |
| [22] | Ackerly D, Knight C, Weiss S, Barton K, Starmer K. Leaf size, specific leaf area and microhabitat distribution of chaparral woody plants:contrasting patterns in species level and community level analyses. Oecologia, 2002, 130(3): 449–457. DOI:10.1007/s004420100805 |
| [23] | Davi H, Barbaroux C, Dufrêne E, François C, Montpied P, Bréda N, Badeck F. Modelling leaf mass per area in forest canopy as affected by prevailing radiation conditions. Ecological Modelling, 2008, 211(3/4): 339–349. |
| [24] | 李芳兰, 包维楷. 植物叶片形态解剖结构对环境变化的响应与适应. 植物学通报, 2005, 22(S1): 118–127. |
| [25] | Lusk C H, Reich P B, Montgomery R A, Ackerly D D, Cavender-Bares J. Why are evergreen leaves so contrary about shade?. Trends in Ecology & Evolution, 2008, 23(6): 299–303. |
| [26] | 付爱红, 陈亚宁, 李卫红, 张宏锋. 干旱、盐胁迫下的植物水势研究与进展. 中国沙漠, 2005, 25(5): 744–749. DOI:10.3321/j.issn:1000-694X.2005.05.021 |
| [27] | González-Rodríguez A M, Martín-Olivera A, Morales D, Jiménez M S. Physiological responses of tagasaste to a progressive drought in its native environment on the Canary Islands. Environmental and Experimental Botany, 2005, 53(2): 195–204. DOI:10.1016/j.envexpbot.2004.03.013 |
| [28] | 魏晓霞, 呼和牧仁, 周梅, 翟洪波, 李良, 邵仁旭, 宋吉祥. 不同年龄华北落叶松叶水势及其影响因素的研究. 干旱区资源与环境, 2010, 24(7): 144–148. |
| [29] | 罗永忠, 成自勇. 水分胁迫对紫花苜蓿叶水势、蒸腾速率和气孔导度的影响. 草地学报, 2011, 19(2): 215–221. |
| [30] | Sack L, Cowan P D, Jaikumar N, Holbrook N M. The 'hydrology' of leaves:co-ordination of structure and function in temperate woody species. Plant, Cell & Environment, 2003, 26(8): 1343–1356. |