 2018, Vol. 38
2018, Vol. 38文章信息
- 龙嘉翼, 赵宇萌, 孔祥琦, 陈治羊, 王秀松, 赵凯, 曹然, 黄丽莎, 吕娇, 崔义, 余玉磊, 徐程扬.
- LONG Jiayi, ZHAO Yumeng, KONG Xiangqi, CHEN Zhiyang, WANG Xiusong, ZHAO Kai, CAO Ran, HUANG Lisha, LÜ Jiao, CUI Yi, YU Yulei, XU Chengyang.
- 观赏灌木小枝和叶性状在林下庇荫环境中的权衡关系
- Trade-offs between twig and leaf traits of ornamental shrubs grown in shade
- 生态学报. 2018, 38(22): 8022-8030
- Acta Ecologica Sinica. 2018, 38(22): 8022-8030
- http://dx.doi.org/10.5846/stxb201801140102
-
文章历史
- 收稿日期: 2018-01-14
- 网络出版日期: 2018-08-24

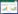




2. 干旱半干旱地区森林培育和生态系统国家林业局重点实验室, 北京 100083
2. Key Laboratory for Silviculture and Forest Ecosystem in Arid and Semi-arid Areas of State Forestry Administration, Beijing 100083, China
耐阴树种的选择是多层次城市森林构建的关键, 在林下配置耐阴树种是实现人工城市森林树种多样性的有效手段。然而枝条上着生的花、果、叶等构件的观赏价值是配置在林下树种不可忽视的因素。如何在不牺牲树木观赏价值的前提下, 构建多层次、多树种混交的城市森林是构建近自然城市森林的重要目标, 也是生产中亟待解决的关键问题。
顶端小枝是植物分枝系统最具有活力的部分, 小枝内部构件的资源分配与形态构建是植物生活史对策研究的重要内容之一[1]。针对小枝性状和叶片性状间的关系, 20世纪40年代末, Corner提出了两种模式:一是枝条越粗, 着生在枝条上面的构件(叶片、花序、果实)就越大;二是分枝越密集, 小枝越纤细, 着生在小枝上的构件越小。这两种模式后被称为Corner法则(Corner's Rules)[2]。之后的学者们对Corner法则中阐述的枝叶性状间的关系在不同环境梯度[3-4]、植物种间和种内的个体发育差异[5-7], 及植物采取的生态学生存策略[8-9]等方面进行过大量研究。这些研究表明, Corner法则在空间的大尺度和小尺度上都普遍适用[3, 10], 小枝大小和小枝上总叶面积也普遍存在正相关的异速生长关系[11], 小枝上叶数量和叶大小则普遍存在显著的权衡关系[12-13], 而小枝内生物量分配的变化则较为复杂[11]。由于植物枝叶性状间的关系对其生存能力和生境的选择都有着重要的影响, 所以小枝与叶片间的关系能够很好地解释植物对环境的适应策略[2]。目前, 关于树木耐阴性或者树木对光受限制环境的适应领域的研究多集中在器官形态与解剖结构[14-15]、光合作用[16-17]等角度, 从枝叶功能性状响应机制与权衡关系相结合的角度研究树木对遮阴环境适应的生态学策略的报道尚少[9, 18]。其中, 关于植物在庇荫环境中的权衡策略存在两种假说:一是“生长-生长”权衡的假说, 其认为耐阴植物会通过提高光截获能力, 降低呼吸强度避免碳消耗来适应遮阴环境[9];二是“生长-生存”权衡的假说, 其认为耐阴植物会通过降低比叶面积(specific leaf area)、叶面积比率(leaf area ratio)、潜在相对生长率[12], 以及形成粗壮、密度高和抗机械干扰能力强枝条[19], 提高养分储存和防御能力来适应遮阴环境[9]。那么, 在庇荫环境中, 灌木为了提高光照资源的利用能力或是生存和防御能力, 是否在小枝和叶性状上采取权衡策略?
因此, 本研究以生长在城市绿地中不同直射光照强度环境中且没有人工修枝、整形等经营措施干扰的金银木(Lonicera maackii)、小花溲疏(Deutzia parviflora)和连翘(Forsythia suspensa)等11种、79株林下灌木作为研究对象, 探讨当年生小枝和叶性状间的关系在林下不同光照强度中的变化规律, 以期揭示灌木在当年生小枝和叶片的构建上会采取何种策略来适应荫庇环境。
1 材料与方法 1.1 光环境测定和样品采集在北京林业大学校园绿地内, 分别于林隙和林冠下两种环境中选择了生长状态良好、生长环境稳定, 且没有人工修枝、整形等经营措施的11种、79株林下落叶灌木(表 1)作为研究对象, 其中, 林隙环境下11种、49株;林冠下7种, 30株。于2017年8月下旬至9月上旬, 选择晴朗无云的天气, 从8:30至17:30, 每隔1小时, 使用SpectroSense2多通道冠层光谱测量仪(Skye, Inc., UK), 参考徐程扬[20]的方法, 分别在每株样木的树冠顶层中央, 以及树冠东、西、南、北方向的1/2冠厚度处设置观测点, 测量光合有效辐射(photosynthetically active radiation, PAR, 400—700 nm, μmol m-2 s-1)、红光(600—700 nm)和远红光(700—800 nm)的光强, 并以某时刻5个观测点的平均光强表示样木该时刻的光环境;另外, 在周围没有高大乔、灌木和其他遮挡物遮挡的旷地上, 设置全光对照。最后, 对灌木不同时刻的PAR平均光强积分后求和得到该样木所处光环境8:30—17:30的PAR累积光量, 来表示灌木的一天中的PAR累积光量。本研究以PAR相对累积光量和红光与远红光的比值代表光环境, 分别依据公式(1)、(2)计算。
| 树种 Species |
株数 Numbers |
树高 Height/m |
冠厚度 Crown thickness/m |
冠幅Crown breadth/m | 光环境 Light environment |
|
| 东-西 East-West |
南-北 South-North |
|||||
| 棣棠Kerria japonica | 2 | 1.90±0.07 | 1.85±0.07 | 2.75±0.49 | 2.60±0.01 | A |
| 风箱果Physocarpus amurensis | 5 | 1.36±0.41 | 1.31±0.41 | 1.92±0.36 | 1.70±0.48 | A, B |
| 金银木Lonicera maackii | 17 | 3.94±0.82 | 2.70±0.62 | 4.96±1.95 | 5.12±2.20 | A, B |
| 锦带Weigela florida | 4 | 2.78±0.71 | 2.72±0.72 | 2.53±1.25 | 2.28±0.62 | A |
| 连翘Forsythia suspensa | 10 | 3.04±0.42 | 2.95±0.43 | 2.89±0.80 | 2.84±1.17 | A, B |
| 太平花Philadelphus pekinensis | 3 | 2.09±0.35 | 1.86±0.17 | 2.17±0.35 | 2.37±0.50 | A |
| 天目琼花Viburnum opulus var.calvescens | 15 | 1.93±0.26 | 1.73±0.20 | 2.20±0.76 | 2.14±0.53 | A, B |
| 猬实Kolkwitzia amabilis | 7 | 2.53±0.51 | 2.00±0.39 | 3.06±0.78 | 2.59±0.66 | A, B |
| 小花溲疏Deutzia parviflora | 6 | 3.05±0.71 | 2.55±0.74 | 3.24±0.93 | 3.71±0.56 | A, B |
| 小叶女贞Ligustrum quihoui | 4 | 3.40±0.59 | 2.21±0.41 | 5.55±1.15 | 5.08±0.82 | A |
| 紫丁香Syringa oblata | 6 | 3.62±2.46 | 2.77±2.17 | 2.86±1.84 | 3.20±1.96 | A, B |
| 总计Total | 79 | 2.87±1.16 | 2.32±0.87 | 3.28±1.66 | 3.27±1.75 | |
| A:林隙, gap;B:林冠下, understory | ||||||

|
(1) |
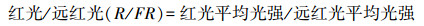
|
(2) |
3种环境的光强均存在极显著(P<0.001)差异:全光>林隙>林冠下。全光对照组的PAR相对累积光量和R/FR均为1.0, 林隙中的PAR相对累积光量是林冠下的7.64倍(0.191和0.025), 林隙中的R/FR是林冠下的1.64倍(0.772和0.470)(图 1)。

|
| 图 1 林冠下和林隙中PAR相对累积光量和R/FR Fig. 1 Mean of PAR relative light summation and R/FR under the tree canopy and in the gap ***表示P<0.001;**表示P<0.01;*表示P<0.05。PAR:光合有效辐射, photosynthetically active radiation;R/FR:红光/远红光, spectral ratio of red light and far-red light |
2017年9月上旬, 参考Pérez-Harguindeguy等的标准测量手册上的方法[21], 从待测灌木树冠的东、西、南、北, 以及对应的树冠上、中、下层分别采集3至5个具有代表性的当年生, 不包含分枝的小枝, 并进行性状指标的测定。
1.2 性状指标测定对于某一样木, 分别测定其每根小枝的小枝长度和每根小枝上的叶片数量, 求平均后得到该样木的平均小枝长度和单位小枝叶数量。从每根样枝上采集分布在枝条中部的具代表性的若干健康成熟叶片, 去除叶柄之后, 通过扫描仪扫描获取叶片图像, 并用Image J软件分析图像数据, 计算得到具代表性叶片的总叶面积, 并除以叶片采集量得到该样木的单叶面积(individual leaf area;ILA), 再用单叶面积乘以单位小枝叶数量, 估算得到该样木的单位小枝叶面积。把小枝和具代表性的叶片放在105℃烘箱中杀青30 min后, 转移到80℃的烘箱中, 烘至恒重, 测定得到小枝干重(twig dry weight;TDW)和具代表性叶片的总干重。具代表性叶片的总干重除以叶片采集量得到其平均叶干重后, 再乘以单位枝条平均叶数量, 估算得到该样木的单位小枝叶干重(leaf dry weight unit twig;LDW);叶面积支持效率(leaf area supporting efficiency)[11]、出叶强度(leafing intensity)[7]和比枝长(specific twig length)[21]分别依据公式(3)、(4)、(5)计算。
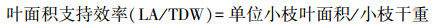
|
(3) |
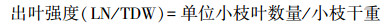
|
(4) |
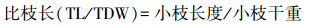
|
(5) |
用植株个体性状的均值除以该种植株性状的均值, 来对该植株个体的性状进行标准化[20], 以消除由于物种间遗传因素差异和量纲不同造成的干扰。然后对标准化后的个体性状指标和光环境指标进行以10为底的对数函数数据转换;运用Pearson相关检验分析光环境指标和灌木性状之间的相关性;灌木枝叶性状之间关系采用log(y)=log(a) + b log(x)进行描述, 式中x和y表示两个不同的性状, a为性状关系的截距, b为斜率, 即相对生长的指数, 当b = 1时, 表示两者是等速变化关系;当b ≠ 1时, 表示两者是异速变化关系。异速生长方程的参数估计采用标准化主轴估计(Standardized major axis estimation, SMA)方法[11], SMA和回归斜率与1或-1显著性检验的计算通过R的“smatr”程序包完成(https://cran.r-project.org/web/packages/smatr/);运用多元回归分析不同光强下, 各性状指标的相互关系[22]。本文使用R.3.3.1和Sigmaplot 12.5进行数据分析、统计和制图。
2 结果 2.1 灌木小枝性状和叶性状间的关系TDW与LDW呈极显著正相关关系(r2=0.79, P<0.001), SMA斜率为0.724(95%的置信区间=0.648—0.809), 极显著小于1.0(P<0.001), 表明TDW与LDW存在极显著(P<0.001)的异速生长关系, 且TDW的增长速率大于LDW的增长速率;LA/TDW与TL/TDW呈极显著正相关关系(r2=0.56, P<0.001), SMA斜率为0.947(95%的置信区间=0.778—1.152), 与1.0无显著差异(P=0.581), 表明LA/TDW与TL/TDW存在极显著(P<0.001)等速生长关系, 表明随LA/TDW的增长, TL/TDW近等速地增长;ILA与LN/TDW呈极显著负相关关系(r2=0.27, P<0.001), SMA斜率为-1.302(95%的置信区间=-1.483—-1.142), 极显著小于-1.0(P<0.001), 表明ILA与LN/TDW存在极显著(P<0.001)异速生长关系, 且LN/TDW的变化速率大于ILA的变化速率(图 2, 图 3)。

|
| 图 2 性状间的异速生长关系 Fig. 2 Allometric relationship between traits ***表示P<0.001;**表示P<0.01;*表示P<0.05;作图数据为原始数据 |

|
| 图 3 性状间的相关关系 Fig. 3 Correlational relationship between traits ***表示P<0.001;**表示P<0.01;*表示P<0.05;作图数据为标准化数据 |
PAR相对累积光量与TDW、LDW都呈极显著正相关(r=0.320, P<0.01;r=0.483, P<0.001), 与TL/TDW、LA/TDW、LN/TDW则都呈极显著负相关(r=-0.480, P<0.001;r=-0.386, P<0.001;r=-0.371, P<0.01), 与ILA相关性不显著(P=0.329);R/FR与TDW、LDW也都呈极显著正相关(r=0.427, P<0.001;r=0.540, P<0.001;), 与TL/TDW、LA/TDW、LN/TDW则都呈极显著负相关(r=-0.594, P<0.001;r=-0.477, P<0.001;r=-467, P<0.001;r=-0.331, P<0.01), 与ILA相关性不显著(P=0.536)。另外, R/FR与各灌木性状(除了ILA)的相关性都大于PAR相对累积光量与各性状的相关性(表 2)。
| TDW | LDW | TL/TDW | LA/TDW | LN/TDW | ILA | |
| PAR相对累积光量 | 0.320** | 0.483*** | -0.480*** | -0.386*** | -0.371** | 0.111(P=0.329) |
| R/FR | 0.427*** | 0.540*** | -0.594*** | -0.477*** | -0.467*** | 0.071(P=0.536) |
| ***表示P<0.001;**表示P<0.01;*表示P<0.05; PAR:光合有效辐射, photosynthetically active radiation;R/FR:红光/远红光, spectral ratio of red light and far-red light;TDW:小枝干重, twig dry weight;LDW:单位小枝叶干重, leaf dry weight unit twig;TL/TDW:比枝长, specific twig length;LA/TDW:叶面积支持效率, leaf area supporting efficiency;LN/TDW:出叶强度, leafing intensity;ILA:单叶面积, individual leaf area | ||||||
R/FR与TDW-LDW、LA/TDW-TL/TDW和ILA-LN/TDW的相关关系均极显著(P<0.001), 决定系数为0.46—0.83。在TDW-R/FR-LDW三维关系中, LDW与TDW、R/FR均呈极显著(P<0.001)正相关关系, TDW维度的斜率更陡峭, 因为TDW的偏回归系数大于R/FR的(0.496和0.256);在LA/TDW-R/FR-TL/TDW三维关系中, TL/TDW与LA/TDW呈极显著(P<0.001)正相关关系, 与R/FR呈极显著(P<0.001)负相关关系, R/FR维度的斜率更加陡峭, 因为LA/TDW的偏回归系数的绝对值小于R/FR的(0.256和-0.281);在ILA-R/FR-LN/TDW三维关系中, LN/TDW与ILA、R/FR呈极显著(P<0.001)负相关关系, ILA维度的斜率更加陡峭, 因为ILA的偏回归系数的绝对值大于R/FR的(-0.886和-0.467)(图 4)。在考虑光质的影响后, 3组性状间关系的斜率, 即相对生长的指数均有所下降(0.543和0.496;0.477和0.256;-0.966和-0.886)(图 3、图 4), 意味着随着R/FR的减弱, LDW随TDW减小而减小的速率减小, TL/TDW随LA/TDW增加而增加的速率减小, ILA随LN/TDW增加而减小的速率减小。

|
| 图 4 R/FR与性状的共变关系 Fig. 4 Covariation of R/FR and traits ***表示P<0.001;**表示P<0.01;*表示P<0.05; R/FR:红光/远红光, spectral ratio of red light and far-red light;TDW:小枝干重, twig dry weight;LA/TDW:叶面积支持效率, leaf area supporting efficiency;ILA:单叶面积, individual leaf area;作图数据为标准化数据 |
灌木枝叶间生物量分配的偏好在植物生态学策略中是普遍存在的现象[23-24]。本研究显示, 随着光强的减小, 灌木当年生小枝和叶片的生物量都趋于减小(表 2), 但是叶干重随小枝干重减小而减小的速率减小(图 3、4), 表现出倾向于叶片生物量的投资偏好。说明植物通常会依据对受限资源的需求进行分配来适应不利光环境[25], 把光合产物更多分配到叶片用于强化叶片构建, 从而达到提高对光照资源的利用能力。不过, 本研究无法针对光以外的其他因素, 如不同树种间遗传差异等进行探讨, 故不能说明光对这种现象有多大的解释能力。但是, 本研究还发现, 灌木当年生小枝和叶片的生物量存在极显著(P<0.001)异速生长关系, 且小枝生物量的增长速率大于叶生物量(图 2), 这与一些研究结果存在差异。这些研究认为, 枝条生物量和叶生物量间是等速生长关系[11, 23], 产生差异的原因可能是研究对象所处环境的不同造成的。本研究重点考虑荫庇环境下的灌木, 研究对象所处环境较为典型, 而其他学者的研究对象所处环境较为复杂多变。不过这样的结果差异也进一步说明了光强的减弱能够使得灌木在枝叶间的生物量分配做出权衡来适应环境。
3.2 灌木对不同光获取构件存在投资偏好根据最优化分配理论, 遮阴环境下的植物会倾向于增加光获取构件的投资[26]。不仅如此, 本研究显示, 在光资源紧张的情况下, 植物会在光获取构件的投资策略上存在偏好。虽然, 基于小枝干重的当年生小枝长度和总叶面积呈等速生长关系(图 2), 说明单位小枝干重上, 枝条总叶面积对枝条长度的变化并没有显著影响。随着光强的减小, 单位小枝干重下, 灌木的当年生小枝长度和枝条总叶面积都趋于增加(表 2), 但比枝长随叶面积支持效率增加而增加的速率却减小(图 3、4), 灌木表现出了相对偏向于增加叶面积的现象。说明灌木在庇荫条件下, 倾向于采取相对忍耐型的光资源利用策略, 通过提高现有树冠区域的总叶片面积来提高对光资源的利用能力, 而非采取通过小枝快速地伸长生长以拓展光资源截获空间的“掠光”策略, 这与Meng等的研究结果相似[27]。原因可能是与树种的遗传特征有关。和灌木相比, 遮阴环境下的乔木幼苗, 尤其是喜光树种幼苗的枝叶生长通常会优先促进枝条生长以占领较高的采光位置[28], 后完成展叶, 表现出显著的不同步性;而耐阴植物则表现出较高的枝叶生长同步性[29]。说明对于通常生长在林冠层之下的灌木来说, 忍耐型的光资源就地利用策略更符合其生活型特征。
3.3 灌木出叶强度和单叶面积的权衡小枝出叶强度和单叶面积的权衡在植物中普遍存在[7, 11, 13]。本研究显示, 随着光强的减小, 灌木当年生小枝的叶数量趋于增加, 而光强对单叶面积的影响不显著(表 2)。这与很多研究结果不符, 这些研究认为随着光强的减小, 植物单叶面积会显著增大, 而出叶强度则趋于减小[13, 30-31]。本研究结果与这些研究结果存在差异的原因可能是观赏灌木的单叶面积对庇荫环境的响应较差。这也说明观赏灌木的耐阴性较差, 弱光环境下灌木对光资源的利用能力不强, 合成的光合产物无法满足个体旺盛生长的需求, 使其偏向于采取忍耐型策略, 与本文3.2中得出的结论相符;另外, 也有可能是因为本研究的研究对象均为落叶灌木, 且叶片通常生长在当年生小枝上, 这意味着这类植物在春天可能会利用去年累积的资源, 在冠层树种树冠郁闭前生长叶片, 增加叶数量, 形成较圆满的树冠, 以此增加光合作用时间, 再利用今年的光合产物维持小枝的进一步生长[18]。但在冠层树种树冠郁闭之后, 又会因为不再有充足的光合产物为单叶面积的增大提供有利条件, 使单叶面积对弱光环境的响应较差。所以, 观赏灌木在庇荫环境下倾向于增大出叶强度, 而其单叶面积却响应较差的现象可能是植物多年生活在庇荫环境下的一种适应行为。
另外, 本研究中灌木小枝的出叶强度和单叶面积呈极显著(P<0.001)异速生长负相关关系, 出叶强度的变化速率大于单叶面积(图 2), 所以当出叶强度随着光强的减弱而增加时, 单叶面积也会趋于减小, 不过减小的速率会降低(图 3、4)。这也与一些研究结果不符, 这些研究认为单叶面积和出叶强度呈显著的等速生长负相关关系[11, 32]。造成结果不同的原因可能是植物叶特性和生活型存在差异。有研究发现, 在针叶、复叶和冠层阔叶树种的叶大小和出叶强度是等速生长关系, 而落叶阔叶林、常绿和落叶阔叶树种、单叶以及亚冠层阔叶树种的则为异速生长关系[33]。这说明了植物叶特性和生活型的差异可能会影响到叶大小和出叶强度间的关系。本研究的研究对象均为单叶的落叶灌木, 意味着它们叶大小和出叶强度间的关系更倾向于异速生长关系, 符合本研究的结果。因此, 总的来说, 在庇荫环境下, 灌木的叶大小和出叶强度间存在一定的权衡关系, 但是观赏灌木整体的耐阴性较差, 其性状对环境的响应能力不强。
综上所述, 小枝和叶生物量、出叶强度和单叶面积存在极显著的异速生长关系, 而比枝长和叶面积支持效率则呈极显著的等速生长关系。林下灌木枝叶的大多数性状(除了单叶面积)都与光环境有极显著的相关性。其中光质对性状的影响整体上大于有效光合辐射的影响, 但是与性状间相互作用相比, 光环境指标与性状间的相关性较低, 说明光环境对性状变化的解释能力较弱, 原因可能是灌木性状的变化还受许多其他因素的影响, 比如遗传差异[34]、水分条件[35]等。不过在不同程度的庇荫环境中, 灌木在当年生小枝和叶片的生物量分配、获光构件的投资策略上存在偏好, 而出叶强度和单叶面积则存在权衡, 具体表现为:随着光强的减弱, 灌木倾向于增加叶生物量的分配, 采取忍耐型光资源利用策略, 通过增加树冠现有区域的总叶面积来提高光能的利用能力。另外在当年生小枝上则倾向于以着生大量小叶的方式来增加单位枝条上的总叶面积, 提高光能截获能力。所以总体上, 林下灌木倾向于采取提高光截获能力的耐阴策略, 与“生长-生长”的权衡假说相符。但根据本研究的结果推断, 观赏灌木的耐阴性较差, 枝叶性状对庇荫环境的响应能力不强, 对弱光资源的利用能力也较差。若种植在林冠下很难保证观赏灌木的旺盛生长和观赏价值的高效发挥, 故建议种植在林缘或旷地等光照充足的环境中。
| [1] | Osada N. Crown development in a pioneer tree, Rhus trichocarpa, in relation to the structure and growth of individual branches. New Phytologist, 2006, 172(4): 667–678. DOI:10.1111/nph.2006.172.issue-4 |
| [2] | Westoby M, Falster D S, Moles A T, Vesk P A, Wright I J. Plant ecological strategies:some leading dimensions of variation between species. Annual Review of Ecology and Systematics, 2002, 33(1): 125–159. DOI:10.1146/annurev.ecolsys.33.010802.150452 |
| [3] | Sun S C, Jin D M, Shi P L. The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient:an invariant allometric scaling relationship. Annals of Botany, 2006, 97(1): 97–107. DOI:10.1093/aob/mcj004 |
| [4] | 张艳茹, 陈红, 王海洋. 低海拔常绿杜鹃小枝繁殖分配与异速生长关系研究. 西南大学学报:自然科学版, 2016, 38(3): 77–82. |
| [5] | Maiti R, Rodriguez H G, Sarkar N C, Sarkar A. Branching pattern and leaf crown architecture of some tree and shrubs in northeast Mexico. International Journal of Bio-resource and Stress Management, 2015, 6(1): 41–50. DOI:10.5958/0976-4038.2015.00009.3 |
| [6] | Cai H Y, Di X Y, Jin G Z. Allometric models for leaf area and leaf mass predictions across different growing seasons of elm tree (Ulmus japonica). Journal of Forestry Research, 2017, 28(5): 975–982. DOI:10.1007/s11676-017-0377-8 |
| [7] | 李曼, 郑媛, 郭英荣, 程林, 卢宏典, 郭炳桥, 钟全林, 程栋梁. 武夷山不同海拔黄山松枝叶大小关系. 应用生态学报, 2017, 28(2): 537–544. |
| [8] | Urbas P, Zobel K. Adaptive and inevitable morphological plasticity of three herbaceous species in a multi-species community:field experiment with manipulated nutrients and light. Acta Oecologica, 2000, 21(2): 139–147. DOI:10.1016/S1146-609X(00)00115-6 |
| [9] | Poorter L. Leaf traits show different relationships with shade tolerance in moist versus dry tropical forests. New Phytologist, 2009, 181(4): 890–900. DOI:10.1111/j.1469-8137.2008.02715.x |
| [10] | 史青茹, 许洺山, 赵延涛, 周刘丽, 张晴晴, 马文济, 赵绮, 阎恩荣. 浙江天童木本植物Corner法则的检验:微地形的影响. 植物生态学报, 2014, 38(7): 665–674. |
| [11] | 李亚男, 杨冬梅, 孙书存, 高贤明. 杜鹃花属植物小枝大小对小枝生物量分配及叶面积支持效率的影响:异速生长分析. 植物生态学报, 2008, 32(5): 1175–1183. DOI:10.3773/j.issn.1005-264x.2008.05.022 |
| [12] | Kleiman D, Aarssen L W. The leaf size/number trade-off in trees. Journal of Ecology, 2007, 95(2): 376–382. DOI:10.1111/jec.2007.95.issue-2 |
| [13] | 史元春, 赵成章, 宋清华, 杜晶, 陈静, 王继伟. 兰州北山刺槐枝叶性状的坡向差异性. 植物生态学报, 2015, 39(4): 362–370. |
| [14] | 王雁.北京市主要园林植物耐荫性及其应用的研究[D].北京: 北京林业大学, 1996. |
| [15] | 殷东生, 沈海龙. 森林植物耐荫性及其形态和生理适应性研究进展. 应用生态学报, 2016, 27(8): 2687–2698. |
| [16] | 张斌斌, 姜卫兵, 翁忙玲, 韩健. 遮阴对园艺园林树种光合特性的影响. 经济林研究, 2009, 27(3): 115–119. DOI:10.3969/j.issn.1003-8981.2009.03.025 |
| [17] | 陆銮眉, 阎光宇, 杜晓娜, 卞阿娜. 8种园林植物耐荫性与光合特性的研究. 热带作物学报, 2011, 32(7): 1249–1254. DOI:10.3969/j.issn.1000-2561.2011.07.013 |
| [18] | Dlugos D M, Collins H, Bartelme E M, Drenovsky R E. The non-native plant Rosa multiflora expresses shade avoidance traits under low light availability. American Journal of Botany, 2015, 102(8): 1323–1331. DOI:10.3732/ajb.1500115 |
| [19] | 冯秋红, 史作民, 董莉莉. 植物功能性状对环境的响应及其应用. 林业科学, 2008, 44(4): 125–131. DOI:10.3321/j.issn:1001-7488.2008.04.023 |
| [20] | 徐程扬.紫椴幼苗、幼树对光的响应与适应研究[D].北京: 北京林业大学, 1999. |
| [21] | Pérez-Harguindeguy N, Díaz S, Garnier E, Lavorel S, Poorter H, Jaureguiberry P, Bret-Harte M S, Cornwell W K, Craine J M, Gurvich D E, Urcelay C, Veneklaas E J, Reich P B, Poorter L, Wright I J, Ray P, Enrico L, Pausas J G, de Vos A C, Buchmann N, Funes G, Quétier F, Hodgson J G, Thompson K, Morgan H D, ter Steege H, van der Heijden M G A, Sack L, Blonder B, Poschlod P, Vaieretti M V, Conti G, Staver A C, Aquino S, Cornelissen J H C. New handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany, 2013, 61(3): 167–234. DOI:10.1071/BT12225 |
| [22] | Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets V, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821–827. DOI:10.1038/nature02403 |
| [23] | 杨冬梅, 毛林灿, 彭国全. 常绿和落叶阔叶木本植物小枝内生物量分配关系研究:异速生长分析. 植物研究, 2011, 31(4): 472–477. |
| [24] | 李肇锋, 潘军, 王金盾, 周东雄, 欧建德. 光环境对闽楠幼树生长及其表型可塑性的影响. 西南林业大学学报, 2014, 34(6): 65–69. |
| [25] | 张志浩, 杨晓东, 孙宝伟, 黄海侠, 马文济, 史青茹, 阎恩荣. 浙江天童太白山不同群落植物构型比较. 生态学报, 2015, 35(3): 761–769. |
| [26] | 李钰, 赵成章, 侯兆疆, 马小丽, 张茜. 高寒退化草地狼毒种群个体大小与茎、叶的异速生长. 生态学杂志, 2013, 32(2): 241–246. |
| [27] | Meng F Q, Zhang G F, Li X C, Niklas K J, Sun S C. Growth synchrony between leaves and stems during twig development differs among plant functional types of subtropical rainforest woody species. Tree Physiology, 2015, 35(6): 621–631. DOI:10.1093/treephys/tpv021 |
| [28] | 徐程扬. 不同光环境下紫椴幼树树冠结构的可塑性响应. 应用生态学报, 2001, 12(3): 339–343. DOI:10.3321/j.issn:1001-9332.2001.03.005 |
| [29] | 蒙凤群.浙江天童亚热带常绿阔叶林木本植物小枝构型的功能生态学研究[D].南京: 南京大学, 2014. |
| [30] | 王祥宁, 熊丽, 陈敏, 杨永福, 余明荣. 不同光照条件下东方百合生长状态及生物量的分配. 西南农业学报, 2007, 20(5): 1091–1096. DOI:10.3969/j.issn.1001-4829.2007.05.048 |
| [31] | 蒋运生, 柴胜丰, 唐辉, 李虹, 黄夕洋, 李锋. 光照强度对广西地不容光合特性和生长的影响. 广西植物, 2009, 29(6): 792–796, 723-723. DOI:10.3969/j.issn.1000-3142.2009.06.016 |
| [32] | 杨冬梅, 占峰, 张宏伟. 清凉峰不同海拔木本植物小枝内叶大小-数量权衡关系. 植物生态学报, 2012, 36(4): 281–291. |
| [33] | 刘长柱, 郭强, 池秀莲. 我国温带山地森林48种常见树种叶片重量-出叶强度的关系. 植物学报, 2015, 50(2): 234–240. |
| [34] | 周刘丽.浙江天童木本植物功能性状种内变异的大小、来源和空间结构[D].上海: 华东师范大学, 2016. |
| [35] | Sánchez-Gómez D, Zavala M A, Valladares F. Functional traits and plasticity linked to seedlings' performance under shade and drought in Mediterranean woody species. Annals of Forest Science, 2011, 65(3): 311–311. |