 2018, Vol. 38
2018, Vol. 38文章信息
- 陈法霖, 张凯, 王芸, 吴爱平, 李有志, 邹冬生, 郑华.
- CHEN Falin, ZHANG Kai, WANG Yun, WU Aiping, LI Youzhi, ZOU Dongsheng, ZHENG Hua.
- 引进种桉树人工林取代天然次生林对土壤微生物群落结构和功能的影响
- Impacts of converting natural secondary forests to exotic Eucalyptus plantations on structure and function of soil microbial communities
- 生态学报. 2018, 38(22): 8070-8079
- Acta Ecologica Sinica. 2018, 38(22): 8070-8079
- http://dx.doi.org/10.5846/stxb201801060036
-
文章历史
- 收稿日期: 2018-01-06
- 网络出版日期: 2018-08-24





2. 中国科学院生态环境研究中心城市与区域生态国家重点实验室, 北京 100085;
3. 新疆农业大学草业与环境科学学院, 乌鲁木齐 830052;
4. 山东省水土保持与环境保育研究所, 临沂大学资源与环境学院, 临沂 276005;
5. 中国科学院大学, 北京 100049
2. State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China;
3. College of Grassland and Environment Sciences, Xinjiang Agricultural University, Urumqi 830052, China;
4. Shandong Provincial Key Laboratory of Water and Soil Conservation and Environmental Protection, College of Resources and Environment, Linyi University, Linyi 276005, China;
5. University of Chinese Academy of Sciences, Beijing 100049, China
砍伐天然林后用外来速生树种造林(如:桉树)是中国乃至全球面临的重大生态变化之一[1-2]。由于外来种对原生生态系统存在潜在的破坏性影响, 人类活动导致的外来种的广泛扩散已受到广泛关注[3]。已有研究表明, 与原生森林相比, 外来物种人工林不仅生长速率快, 导致土壤水分和养分资源的大量消耗[4], 其生态系统碳输入的数量和质量也发生改变[5], 并且林下植被的生长还常受到林业作业的物理干扰[6]以及外来物种的化学干扰[7]。
为了强调人工林的经济效益, 我国在植树造林过程中引进了一些速生且能适应多种环境的外来物种。桉树(Eucalyptus), 桃金娘科(Myrtaceae), 桉树属(Eucalyptus), 原产于澳大利亚, 已成为全球以及我国南方发展速生丰产林的重要造林树种之一。现全球桉树人工林面积超过2000万hm2, 占世界人工林面积的15%, 而我国桉树造林面积在2015年已经超过450万hm2, 位列世界第三[8]。然而, 种植桉树可能会对引入地的环境产生有害影响, 例如:生物多样性下降, 土壤侵蚀, 土壤肥力下降, 从而削弱其益处[9-10]。已有研究表明引进种桉树造林显著影响了我国华南地区森林土壤的物理和化学性质以及植物群落的多样性[6, 11], 但有关桉树人工林对华南地区土壤微生物群落结构和功能影响的研究较少。
本研究以引进种桉树造林对土壤微生物群落的影响为主线, 应用成对试验设计(天然次生林-桉树人工林), 采用磷脂脂肪酸图谱(PLFA)和BIOLOG微平板培养等技术手段, 探讨引进种桉树人工林取代天然次生林对土壤微生物群落生物量、结构和功能的影响, 揭示导致土壤微生物群落变化的主要因素, 为明确大面积桉树造林对华南地区森林土壤微生物多样性的影响, 以及桉树人工林的可持续经营提供理论依据。
1 材料与方法 1.1 试验点概况为了全面揭示桉树种植对土壤微生物群落的影响, 本研究选择海南省白沙县细水乡龙村小流域开展研究。白沙东临琼中县, 西接昌江县, 北抵儋州市, 南临乐东县。面积约为2118 km2, 境内41.9%为山地。白沙属于热带湿润季风性气候, 年平均降雨量约1725 mm, 并且70%—80%的降水集中在5—10月。年平均气温21.9—23.4℃, 最高温度35—37℃, 最低温度5—6℃。土壤母质属于花岗岩风化物, 土壤类型为砖红壤。地带性森林群落为热带季雨林。
本实验区域天然次生林中物种丰富, 乔木物种主要有厚皮树, 对叶榕, 银柴, 木棉, 倒吊笔, 菲律宾合欢, 海南红豆, 海南蒲桃, 枫香, 黄牛木, 毛果扁担杆, 水锦树以及越南山矾, 林下灌木物种主要有红腺紫珠, 假苹婆, 假鹰爪, 九节, 牛筋果, 破布叶等, 主要的草本物种则有竹叶草, 弓果黍, 细花百部, 求米草, 华南毛蕨, 海南老鸭嘴, 大叶竹叶草等。
桉树人工林在种植前会进行炼山、翻耕整地、施基肥, 种植后的前三年每年追肥并除草一次, 林下灌草以马缨丹和飞机草最为常见。轮伐期较短, 一般5—6年。第一代桉树人工林造林方式为植苗, 第二代则为萌芽, 第三代植苗, 第四代萌芽, 依次类推。
1.2 试验设计及采样以天然次生林为对照, 为了比较桉树人工林取代天然次生林造林后土壤微生物群落结构和功能的变化, 确保研究结果的可比性, 本研究在海南白沙县细水乡龙村小流域随机选取了10对(样点坐标范围为109°34'—109°35'E, 19°8'—19°10'N)相邻的天然次生林和桉树林作为研究对象。野外样方调查和采样时间为2011年10月。采样点桉树人工林造林时间为2002—2008年, 造林之前均为与对照一样的天然次生林, 调查样地均为一代或二代桉树林, 林龄为3—5年。
每块供试林地选取3个独立的面积为20 m×20 m乔木样方, 调查样方乔木层盖度及样方内所有乔木物种的株高、胸径和株数。每个乔木样方内调查2个面积为5 m×5 m的灌木样方以及3个面积为1 m×1 m的草本样方, 分别调查灌木和草本层盖度及样方内所有灌木和草本的株数。
每块供试林地的3个乔木样方中, 分别随机选取10处用直径为3.6 cm的土钻取0—10 cm土壤, 混为一个土壤样品, 每块林地3个土壤样品。采集的土壤样品放在冰盒中(4℃)保存, 并在3天内运回实验室。采回的土壤样品过2 mm孔径的土壤筛进行匀质化并去除较大的石块、土壤动物及植物残体。过筛后的土壤一部分放在4℃保存用于土壤微生物生物量、碳源代谢功能等的测定; 一部分冷冻干燥后, 磨碎并过100目土壤筛, 保存于-20℃用于土壤PLFA的提取; 其余的土样风干用于土壤理化性质分析。不同森林类型的植被及土壤基本特征见表 1。
| 植被及土壤特征 Plant and soil parameters |
天然次生林 Natural secondary forest |
桉树林 Eucalyptus plantation |
| 乔木层盖度Tree coverage/% | 78±4 | 41±3 |
| 灌木层盖度Shrub coverage/% | 47±4 | 43±6 |
| 草本层盖度Herb coverage/% | 61±7 | 86±5 |
| 乔木层物种丰富度Species richness of tree layer | 11±1 | 1 |
| 灌木层物种丰富度Species richness of shrub layer | 17±1 | 9±1 |
| 草本层物种丰富度Species richness of herb layer | 13±1 | 13±1 |
| 含水量Water content/% | 21.82±0.44 | 19.90±0.49 |
| pH | 5.55±0.14 | 5.11±0.13 |
| 总碳Total C/(mg/g) | 24.18±1.14 | 18.77±1.11 |
| 总氮Total N/(mg/g) | 2.20±0.09 | 1.86±0.10 |
| 总磷Total P/(μg/g) | 436.01±27.16 | 496.62±44.14 |
| 总钾Total K/(mg/g) | 16.00±0.73 | 19.07±0.65 |
| 有机碳Total organic C/(mg/g) | 23.14±0.97 | 17.52±1.15 |
| 碱解氮Alkaline hydrolyzable-nitrogen/(μg/g) | 156.59±3.20 | 126.27±7.00 |
| 铵态氮NH4+/(μg N/g) | 12.31±0.65 | 10.66±0.29 |
| 硝态氮NO3-/(μg N/g) | 3.72±0.61 | 2.44±0.18 |
| 有效磷Available P/(μg/g) | 5.47±0.61 | 11.71±2.17 |
| 速效钾Available K/(μg/g) | 93.87±15.75 | 98.55±5.87 |
土壤微生物生物量碳、氮采用氯仿熏蒸-K2SO4浸提法[12]进行前处理, 实验在样品取回后24 h内进行。测定生物量碳所用仪器为德国Elementar公司生产的Liqui TOC Ⅱ分析仪。生物量氮的测定采用碱性过硫酸钾消解-紫外分光光度法。
1.4 土壤微生物群落的磷脂脂肪酸组成分析土壤微生物磷脂脂肪酸的测定方法在Buyer[13]的方法上加以改进。所用试验用品和器皿均为玻璃或Teflon(特氟龙)材料, 不使用清洗剂清洗容器。提取过程避光, 远离热源。样品冷冻保存, 避光、水和氧气。主要步骤为:提取:(1)4 g冻干土(100目)至40 mL玻璃离心管, 依次加入3.2 mL磷酸缓冲液、8 mL甲醇、4 mL氯仿; 于室温超声浸提10 min后避光水平震荡1 h, 2500 rpm/min离心10 min, 收集上清至100 mL分液漏斗; (2)依次加入3.2 mL磷酸缓冲液(pH 7.4)、4 mL氯仿, 混匀, 避光过夜分层; (3)收集下层氯仿相至40 mL玻璃离心管, N2吹干。分离:(4)过硅胶柱(500 mg silica gel column, Part No.5982-2265, Agilent Technologies, Wilmington, DE, USA):样品过柱前先加入5 mL氯仿润湿硅胶柱; 再用10 mL氯仿分2次洗涤转移N2吹干的样品至硅胶柱内; (5)待氯仿滴干后加入10 mL丙酮过硅胶柱; (6)待丙酮完全滴干后, 加入5 mL甲醇(色谱纯)过硅胶柱, 收集甲醇相至10 mL Teflon离心管, N2吹干。甲酯化:(7)用1 mL甲醇-甲苯(1: 1, v/v)溶液溶解吹干的脂类物质; (8)加入0.2 mol/L KOH(现用现配, 甲醇做溶剂)1 mL, 混匀, 35℃温育15 min; (9)待样品冷却至室温后, 依次加入2 mL氯仿-正己烷(1: 4, v/v)、1 mL 1 mol/L醋酸、2 mL超纯水, 混匀; (10)2000 rpm/min离心5 min, 收集上层正己烷相至干净的10 mL样品瓶; (11)加入2 mL氯仿-正己烷(1: 4, v/v)重复提取一次, 合并两次提取的正己烷相, N2吹干, -20℃保存。上机前准备:(12)过柱(NH2 SPE column, Part No. 8B-S009-EAK, Phenomenex, Torrance, CA):过柱前先加入1 mL氯仿润湿柱子; 再用1 mL氯仿分2次洗涤转移吹干的样品至柱子内; (13)收集氯仿相至2 mL棕色样品瓶, N2吹干; (14)加入1 mL含有10 ppm内标物(十九烷酸甲酯)的正己烷(农残级)溶液。
磷脂脂肪酸的定量采用气质联用仪进行测定。GC-MS条件:HP6890/MSD5793(Agilent Technologies, Bracknell, UK), HP-5毛细管柱(30 m×0.25 mm×0.25 μm), 不分流进样。进样口温度230℃; 检测器温度270℃。升温程序:50℃持续1 min, 以15℃/min增加至150℃, 保持2 min, 再以3℃/min增加至250℃, 保持15 min。He作载气, 流量为1 mL/min。
脂肪酸定量用峰面积和内标(19:0)曲线法。PLFA含量用nmol/g表示。
本研究共有24种PLFA被检出。这些脂肪酸主要由饱和脂肪酸、不饱和脂肪酸、带甲基支链的脂肪酸和带环丙基的脂肪酸组成。主要微生物类群的生物量通过以下PLFA的总量来估算:细菌(15:0、17:0、20:0、革兰氏阳性细菌:i15:0、a15:0、i16:0、i17:0、a17:0、i18:0;革兰氏阴性细菌:16:1ω9t、16:1ω7c、cy17:0、18:1ω9t、18:1ω7c、cy19:0);用16:1ω5c(丛枝菌根真菌)、18:2ω6, 9c、18:1ω9c的和来估算真菌的生物量; 用10Me17:0、10Me18:0与10Me19:0的和来估算放线菌的生物量, 14:0、16:0、18:0为通用脂肪酸[14-17]。
1.5 碳源代谢功能分析微生物群落碳源功能应用BIOLOG微生物自动分析系统进行测定[18]。BIOLOG实验在取样后一周内进行。称取10 g鲜土, 加入90 mL无菌的0.145 mol/L的NaCl溶液, 在摇床上180 rpm/min往复式振荡30 min, 然后将土壤悬浊液再稀释100倍, 用8通道移液器吸取150 μL稀释过后的土壤悬浊液接种至生态板(BIOLOG-ECO PLATE)的每一个孔中, 将接种好的板放置于25℃恒温培养10 d, 每12 h用BIOLOG微生物自动分析系统测定590 nm波长处的光密度值。
孔的平均颜色变化率(Average well colour development, AWCD)计算方法如下[19]:
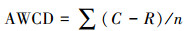
|
式中, C为每个有培养基孔的光密度值, R为对照孔的光密度值, n为培养基碳源种类, Eco板n值为31。C-R < 0的孔在计算时全部替换为0。
采用曲线整合方法[20]估计碳源代谢活性:
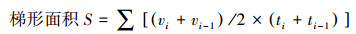
|
式中, vi为i时刻的AWCD值。
本研究用培养72 h的数据来表征BIOLOG板中的微生物代谢功能多样性特征:包括利用碳源的丰富度(Richness, S)、香农多样性指数(Shannon index, H')。计算丰富度时以(C-R) > 0.25的数据为阳性值。
香农多样性指数的计算公式为:
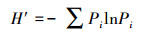
|
式中, Pi=ni/N, ni是第i种培养基的光密度值, N为样品中所有培养基光密度值的总和。
1.6 数据处理本研究以样点为彼此独立的重复, 所以统计分析时取每个样点3个土样的均值进行分析。用成对T检验(Paired-t-test)分析引进种桉树取代天然次生林造林对微生物群落特征是否有显著影响, 所用软件为SPSS 16.0(SPSS Inc., Chicago, IL, USA)。用R中vegan包的bioenv程序筛选出显著影响土壤微生物群落的土壤因子和植物因子, 再用典型对应分析(Canonical Correspondence Analysis, CCA)分析筛选出的环境因子与土壤微生物落结构及功能之间的关系, 统计分析通过CANOCO 4.5实现。用SigmaPlot 11.0(Systat Software Inc., Chicago, IL, USA)绘图。
2 结果 2.1 土壤微生物量碳和氮桉树取代天然次生林造林导致土壤微生物量碳、氮显著降低。天然次生林土壤微生物量碳为631.86 mg/kg, 生物量氮为94.72 mg/kg, 而桉树人工林土壤微生物量碳为376.65 mg/kg, 生物量氮则为61.91 mg/kg, 分别比天然次生林低40%(P < 0.001)和35%(P < 0.01)(图 1)。

|
| 图 1 不同森林土壤微生物生物量碳和氮 Fig. 1 Soil microbial biomass carbon and nitrogen in different forests |
两种林型土壤微生物群落的磷脂脂肪酸总量间差异显著并达到极显著水平(表 2, P < 0.001), 桉树人工林土壤微生物群落的磷脂脂肪酸总量为67.6 nmol/g, 而天然次生林则达到105.9 nmol/g, 是桉树林的1.6倍。不管是革兰氏阳性菌, 还是革兰氏阴性菌的特征脂肪酸含量, 桉树人工林都显著低于天然次生林; 天然次生林土壤中细菌的磷脂脂肪酸含量比桉树人工林高58%。天然次生林土壤中真菌的磷脂脂肪酸含量也显著高于桉树人工林, 特别是丛枝菌根真菌特征脂肪酸的含量是桉树人工林的1.8倍。另外, 桉树人工林土壤放线菌特征脂肪酸的含量也比天然次生林低39%。
| 特征磷脂脂肪酸 Characteristic PLFA |
天然次生林 Natural secondary forest |
桉树林 Eucalyptus plantation |
| 革兰式阳性细菌Gram-positive bacteria/(nmol/g) | 40.20±2.7*** | 26.70±2.3 |
| 革兰式阴性细菌Gram-negative bacteria/(nmol/g) | 18.20±1.4*** | 10.50±0.9 |
| 细菌Bacteria/(nmol/g) | 59.90±4.1*** | 38.00±3.3 |
| 丛枝菌根真菌Arbuscular mycorrhizal fungi/(nmol/g) | 4.20±0.2*** | 2.30±0.2 |
| 真菌Fungi/(nmol/g) | 12.10±0.8*** | 7.70±0.6 |
| 放线菌Actinomycetes/(nmol/g) | 9.30±0.7*** | 5.70±0.6 |
| 磷脂脂肪酸总量Total PLFA/(nmol/g) | 105.90±7.3*** | 67.60±5.5 |
| 饱和直链/单不饱和脂肪酸(SAT/MONO)Saturated/Monounsaturated PLFA | 0.90±0.02 | 1.02±0.03** |
| 革兰氏阳性/阴性菌(GP/GN)Gram+/Gram- bacteria | 2.24±0.08 | 2.56±0.05** |
| cy19:0/18:1ω7c | 6.10±0.58 | 9.35±1.10* |
| 异构/反异构脂肪酸(Ⅰ/A)Iso-/Anteiso-branched PLFA | 1.83±0.10 | 2.16±0.09* |
| *表示P < 0.05, **表示P < 0.01, ***表示P < 0.001 | ||
饱和直链脂肪酸/单不饱和脂肪酸(SAT/MONO)、革兰氏阳性菌/革兰氏阴性菌(GP/GN)、异构/反异构脂肪酸(Ⅰ/A)以及环丙烷脂肪酸/前体(cyc/pre)的比值常用来指示土壤微生物受生理胁迫的程度, 随受胁迫程度的增加而升高。与天然次生林相比, 桉树造林显著改变了土壤微生物群落的磷脂脂肪酸结构:桉树人工林土壤微生物群落的SAT/MONO、GP/GN、Ⅰ/A以及cy19:0/18:1ω7c的比值上升, 并且都达到显著水平(表 2), 表明与天然次生林相比, 桉树人工林土壤微生物群落受生理胁迫增强。
2.3 土壤微生物群落碳代谢功能与天然次生林相比, 桉树的引种显著降低了土壤微生物群落的碳源代谢活性(表 3, P < 0.001)。培养72 h后, 桉树林地土壤微生物群落利用碳源的丰富度及香农多样性指数也都显著低于天然次生林(表 3, P < 0.001)。桉树林土壤微生物群落利用碳源的种类为17.7种, 而天然次生林土壤微生物群落利用碳源的种类达到23.4种。桉树林土壤微生物群落利用碳源的多样性指数只有2.89, 而天然次生林土壤微生物群落利用碳源的多样性指数为3.10。
| 森林类型 Forest type |
代谢活性 Metabolic intensity |
丰富度 Richness |
多样性指数 Shannon diversity |
| 天然次生林Natural secondary forest | 296.74±6.26*** | 23.4±0.8*** | 3.10±0.02*** |
| 桉树林Eucalyptus plantation | 253.89±7.62 | 17.7±0.6 | 2.89±0.02 |
| ***表示P < 0.001 | |||
桉树人工林和天然次生林土壤微生物群落磷脂脂肪酸组成的差异与桉树林的乔木盖度、灌木丰富度、土壤含水量、pH、碱解氮、有机碳低于天然次生林有关。第一轴代表了土壤微生物群落磷脂脂肪酸组成38.7%的变异, 第二轴代表了土壤微生物群落磷脂脂肪酸组成11.3%的变异。乔木盖度及灌木的物种丰富度是影响土壤微生物群落磷脂脂肪酸组成的主要植物因子, 累积可解释20.0%的变异; 土壤含水量、pH、碱解氮、有机碳是影响土壤磷脂脂肪酸组成的主要土壤因子, 累积可解释51.5%的变异; 由于植物因子与土壤因子间存在交互作用, 以上6种植物及土壤因子一共可解释62.5%的变异(图 2)。

|
| 图 2 土壤微生物群落PLFA组成和碳代谢功能与环境因子的典型对应分析及方差分解 Fig. 2 Canonical correspondence analysis and partition of variation showing relations between environmental factors and soil microbial PLFA composition, carbon utilization profiles TRC:乔木层盖度, Tree coverage; TRR:乔木层物种丰富度, Species richness of tree layer; SHR:灌木层物种丰富度, Species richness of shrub layer; WC:土壤含水量, Soil water content; pH:土壤酸碱度, Soil pH; SOC:土壤有机碳, Soil organic carbon; AN:土壤碱解氮, Soil alkaline hydrolyzable-nitrogen |
对土壤微生物群落的碳源代谢方式做CCA表明:桉树人工林和天然次生林在第一轴上差异显著, 主要与桉树人工林的乔木物种丰富度及盖度、灌木物种丰富度、土壤水分、pH及碳、氮资源可利用性低于天然次生林有关。第一轴代表了土壤微生物群落碳源代谢23.6%的变异, 第二轴代表了土壤微生物群落碳源代谢10.3%的变异。乔木的物种丰富度及盖度、灌木的物种丰富度是影响土壤微生物群落碳源代谢的主要植物因子, 累积可解释42.2%的变异; 土壤含水量、pH、碱解氮、有机碳是影响土壤微生物群落碳源代谢的主要土壤因子, 累积可解释44.2%的变异; 由于以上植物因子与土壤因子之间存在强烈的交互作用, 7种因子累积可解释土壤微生物群落碳代谢方式变异的48.6%(图 2)。
3 讨论森林土壤微生物群落的结构和功能受森林类型、气候、土壤条件以及人为管理活性等的影响[1, 21]。本研究以土壤微生物群落的结构和功能如何响应桉树人工林取代天然次生林这一变化为主线, 选取了10对相邻的天然次生林和桉树人工林, 不同的样地其土壤和小气候条件也各不相同, 使得结果更具有说服力及代表性。结果表明:桉树取代天然次生林造林后, 土壤微生物群落的结构和功能发生了显著的改变, 而这些变化与森林类型转变导致的环境因子的改变有关。
3.1 桉树人工林取代天然次生林对土壤微生物群落的影响土壤微生物是土壤生态系统变化的敏感指标之一, 能有效反映出土壤生态系统的质量, 微生物生物量越大, 土壤质量越高[22-23]。本研究中, 与天然次生林相比, 桉树人工林中无论是土壤微生物量碳、氮, 还是各类群微生物的特征磷脂脂肪酸的含量都显著下降, 表明桉树人工林土壤质量有所退化。土壤微生物群落结构的变化, 例如:饱和直链脂肪酸/单不饱和脂肪酸, 真菌/细菌, 革兰氏阳性/阴性菌, 异构/反异构支链磷脂脂肪酸以及环丙基脂肪酸/前体的比值, 与土壤微生物受生理胁迫的程度以及土壤营养胁迫条件显著正相关, 或与资源的可用性显著负相关[24-27]。以上磷脂脂肪酸比值, 随着森林类型的转变, 都表现出显著升高的趋势, 这些显著变化指示了森林土壤资源可利用性的下降以及土壤养分胁迫的增强[1, 9, 28-29]。
天然次生林转变为桉树人工林显著降低了土壤微生物群落的碳代谢功能。土壤微生物的碳代谢活性能很好的指示土壤微生物的活性, 土壤微生物群落利用单一碳源的丰富度和多样性常用来反映土壤微生物的功能多样性[23]。BIOLOG代谢多样性模式的变化与群落组成的变化相关[30]。本试验结果表明从天然次生林向桉树人工林的转变导致土壤微生物群落碳代谢活性和多样性的显著下降, 并且代谢模式也发生了显著改变。土壤微生物群落的功能与森林生态系统养分循环过程息息相关, 已有研究同样发现外来种造林导致森林或者土壤生态系统中土壤微生物群落功能的下降[5, 23, 31-33]。桉树人工林的土壤微生物群落相对于天然次生林具有较低的碳代谢功能, 也表明天然次生林向引进种桉树人工林的转变导致土壤质量、土壤微生物群落多样性甚至生态系统功能的下降。
3.2 桉树取代天然次生林造林影响土壤微生物群落的机制桉树人工林取代天然次生林后土壤微生物群落结构和功能发生了显著的改变。与天然次生林相比, 外来种桉树造林导致土壤微生物群落结构和功能退化的可能因素如下:
(1) 桉树本身的影响。桉树生长迅速, 生长过程中会利用大量的土壤水分以及养分。如此高的养分需求导致土壤水分和养分的大量损失[34], 从而导致土壤资源可利用性的下降。土壤理化性质在调节微生物群落结构和功能方面具有重要作用[5, 35]。Behera和Sahani[36]的研究发现由天然林转变为桉树人工林会导致土壤持水能力、有机碳、总氮以及微生物生物量及其代谢熵的下降。本研究中, 与天然次生林相比, 引进种桉树人工林的土壤水分含量、土壤有机碳以及碱解氮的含量较低, CCA分析结果也表明土壤微生物群落结构和功能的退化与上述因素有关。
桉树较小的冠层导致了桉树人工林较低的乔木层盖度, 本研究10片调查样地均值只有41%。另外, 桉树的生理(例如:较高的生长速率)和化学特征(例如:桉树叶片、树皮、和根内酚酸类和挥发油类的释放)有可能对林下层植被产生抑制作用[7, 37]。本研究结果发现, 天然次生林转变为桉树人工林后, 林下层植物的物种组成发生了显著的变化, 林下灌木层的丰富度也显著降低。森林植被对土壤微生物群落的影响巨大, 乔木层盖度以及林下灌木层丰富度的减少不仅导致土壤资源输入多样性和量的减少, 也导致微气候条件的恶化, 例如:导致土壤含水量的减少, 而土壤含水量的减少会影响有机质的分解速率[38-39]。本研究结果也发现, 地上部植物特征与土壤性质之间存在交互作用, 共同影响着土壤微生物群落结构和功能。总之, 森林植物群落的改变和各功能群对土壤养分及生境的影响显著的改变了土壤微生物群落结构和功能。
(2) 人工林管理, 例如大面积单一树种造林、炼山以及不合理的人工抚育会通过改变植物群落的结构和土壤资源的可利用性而影响土壤微生物群落的组成和功能。大面积单一树种造林导致乔木层物种单一, 直接影响土壤微生物可获得资源的多样性。炼山可以通过高温导致热敏感微生物种群的消亡而直接影响土壤微生物群落, 或者通过改变土壤的物理化学性质(例如:碳的质量、矿质营养的含量和土壤的容重[40-41]和土壤二氧化碳的排放[42]而间接的影响土壤微生物群落)。有研究表明炼山会导致土壤微生物生物量和丰度的显著下降[42-45]。桉树人工林的抚育和高强度的林业作业(例如:种植桉树前过度的样地翻耕整地, 除草、桉树木材收获过程中的伐木作用以及地上生物量的移除)也可能直接或间接的影响土壤微生物群落的结构和功能。本研究区域内的桉树人工林在造林及采伐前后, 为了方便人工进行林业作业, 会有目的地通过炼山来清理林地, 再加上种植过程中的各种人为干扰不仅导致林下层植被的破坏, 还破坏土壤结构, 即使桉树种植的头三年每年都对桉树林进行施肥, 这些人为干扰仍然可能导致土壤养分资源输入的减少和土壤养分、水分损失的增加[23], 这可能是桉树林土壤微生物功能退化的原因之一。
桉树人工林取代天然次生林导致土壤微生物群落的生物量以及碳代谢功能显著降低、致使土壤微生物群落生理胁迫显著增强, 导致土壤微生物群落结构变化和功能退化的主要环境因素为植物物种丰富度和覆盖度的降低以及土壤资源(碳、氮和水分)可获得性的降低。因此, 桉树种植和管理过程中, 建议减少对林下植被以及土壤干扰、增加土壤资源可获得性, 进而减缓桉树种植对土壤微生物群落的不利影响。
| [1] | Burton J, Chen C R, Xu Z H, Ghadiri H. Soil microbial biomass, activity and community composition in adjacent native and plantation forests of subtropical Australia. Journal of Soils and Sediments, 2010, 10(7): 1267–1277. DOI:10.1007/s11368-010-0238-y |
| [2] | Iovieno P, Alfani A, Bååth E. Soil microbial community structure and biomass as affected by Pinus pinea plantation in two mediterranean areas. Applied Soil Ecology, 2010, 45(1): 56–63. DOI:10.1016/j.apsoil.2010.02.001 |
| [3] | Wardle D A, Bellingham P J, Fukami T, Bonner K I. Soil-mediated indirect impacts of an invasive predator on plant growth. Biology Letters, 2012, 8(4): 574–577. DOI:10.1098/rsbl.2012.0201 |
| [4] | Tunlid A, Hoitink H A J, Low C, White D C. Characterization of bacteria that suppress Rhizoctonia damping-off in bark compost media by analysis of fatty acid biomarkers. Applied and Environmental Microbiology, 1989, 55(6): 1368–1374. |
| [5] | 张凯, 郑华, 陈法霖, 欧阳志云, 兰俊, 吴永富, 项学武. 桉树取代马尾松对土壤养分和酶活性的影响. 土壤学报, 2015, 52(3): 646–653. |
| [6] | 朱宏光, 温远光, 梁宏温, 徐海根, 杨瑶青, 李明臣, 黄志辉, 邓荣艳. 广西桉树林取代马尾松林对植物多样性的影响. 北京林业大学学报, 2009, 31(6): 149–153. |
| [7] | Zhang C L, Fu S L. Allelopathic effects of eucalyptus and the establishment of mixed stands of eucalyptus and native species. Forest Ecology and Management, 2009, 258(7): 1391–1396. DOI:10.1016/j.foreco.2009.06.045 |
| [8] | 谢耀坚. 论中国桉树发展的贡献和可持续经营策略. 桉树科技, 2016, 33(4): 26–31. DOI:10.3969/j.issn.1674-3172.2016.04.006 |
| [9] | Berthrong S T, Schadt C W, Pineiro G, Jackson R B. Afforestation alters the composition of functional genes in soil and biogeochemical processes in south american grasslands. Applied and Environmental Microbiology, 2009, 75(19): 6240–6248. DOI:10.1128/AEM.01126-09 |
| [10] | Lynch H B, Epps K Y, Fukami T, Vitousek P M. Introduced canopy tree species effect on the soil microbial community in a Montane tropical forest. Pacific Science, 2012, 66(2): 141–150. DOI:10.2984/66.2.4 |
| [11] | 温远光, 郑羡, 李明臣, 徐海根, 梁宏温, 黄承标, 朱宏光, 何斌. 广西桉树林取代马尾松林对土壤理化性质的影响. 北京林业大学学报, 2009, 31(6): 145–148. |
| [12] | Vance E D, Brookes P C, Jenkinson D S. An extraction method for measuring soil microbial biomass C. Soil Biology and Biochemistry, 1987, 19(6): 703–707. DOI:10.1016/0038-0717(87)90052-6 |
| [13] | Buyer J S, Teasdale J R, Roberts D P, Zasada I A, Maul J E. Factors affecting soil microbial community structure in tomato cropping systems. Soil Biology and Biochemistry, 2010, 42(5): 831–841. DOI:10.1016/j.soilbio.2010.01.020 |
| [14] | Olsson P A. Signature fatty acids provide tools for determination of the distribution and interactions of mycorrhizal fungi in soil. FEMS Microbiology Ecology, 1999, 29(4): 303–310. DOI:10.1111/fem.1999.29.issue-4 |
| [15] | Zelles L. Phospholipid fatty acid profiles in selected members of soil microbial communities. Chemosphere, 1997, 35(1/2): 275–294. |
| [16] | Zelles L. Fatty acid patterns of phospholipids and lipopolysaccharides in the characterisation of microbial communities in soil:a review. Biology and Fertility of Soils, 1999, 29(2): 111–129. DOI:10.1007/s003740050533 |
| [17] | Zelles L, Palojärvi A, Kandeler E, von Lützow M, Winter K, Bai Q Y. Changes in soil microbial properties and phospholipid fatty acid fractions after chloroform fumigation. Soil Biology and Biochemistry, 1997, 29(9/10): 1325–1336. |
| [18] | Winding A. Fingerprinting bacterial soil communities using biolog microtitre plates//Ritz K, Dighton J, Giller K E, eds. Beyond the Biomass. London: Wiley, 1994: 85-94. |
| [19] | Garland J L, Mills A L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Applied and Environmental Microbiology, 1991, 57(8): 2351–2359. |
| [20] | Hackett C A, Griffiths B S. Statistical analysis of the time-course of Biolog substrate utilization. Journal of Microbiological Methods, 1997, 30(1): 63–69. DOI:10.1016/S0167-7012(97)00045-6 |
| [21] | Ibell P T, Xu Z H, Blumfield T J. Effects of weed control and fertilization on soil carbon and nutrient pools in an exotic pine plantation of subtropical Australia. Journal of Soils and Sediments, 2010, 10(6): 1027–1038. DOI:10.1007/s11368-010-0222-6 |
| [22] | Powlson D S, Brookes P C, Christensen B T. Measurement of soil microbial biomass provides an early indication of changes in total soil organic matter due to straw incorporation. Soil Biology and Biochemistry, 1987, 19(2): 159–164. DOI:10.1016/0038-0717(87)90076-9 |
| [23] | Zheng H, Ouyang Z Y, Wang X K, Fang Z G, Zhao T Q, Miao H. Effects of regenerating forest cover on soil microbial communities:a case study in hilly red soil region, Southern China. Forest Ecology and Management, 2005, 217(2/3): 244–254. |
| [24] | Bossio D A, Scow K M. Impacts of carbon and flooding on soil microbial communities:phospholipid fatty acid profiles and substrate utilization patterns. Microbial Ecology, 1998, 35(3/4): 265–278. |
| [25] | Fierer N, Schimel J P, Holden P A. Variations in microbial community composition through two soil depth profiles. Soil Biology and Biochemistry, 2003, 35(1): 167–176. DOI:10.1016/S0038-0717(02)00251-1 |
| [26] | Mckinley V L, Peacock A D, White D C. Microbial community PLFA and PHB responses to ecosystem restoration in tallgrass prairie soils. Soil Biology and Biochemistry, 2005, 37(10): 1946–1958. DOI:10.1016/j.soilbio.2005.02.033 |
| [27] | Moore-Kucera J, Dick R P. PLFA profiling of microbial community structure and seasonal shifts in soils of a Douglas-fir chronosequence. Microbial Ecology, 2008, 55(3): 500–511. DOI:10.1007/s00248-007-9295-1 |
| [28] | Chen C R, Xu Z H, Mathers N J. Soil carbon pools in adjacent natural and plantation forests of subtropical Australia. Soil Science Society of America Journal, 2004, 68(1): 282–291. DOI:10.2136/sssaj2004.2820 |
| [29] | Macdonald C A, Thomas N, Robinson L, Tate K R, Ross D J, Dando J, Singh B K. Physiological, biochemical and molecular responses of the soil microbial community after afforestation of pastures with Pinus radiata. Soil Biology and Biochemistry, 2009, 41(8): 1642–1651. DOI:10.1016/j.soilbio.2009.05.003 |
| [30] | Haack S K, Garchow H, Klug M J, Forney L J. Analysis of factors affecting the accuracy, reproducibility, and interpretation of microbial community carbon source utilization patterns. Applied and Environmental Microbiology, 1995, 61(4): 1458–1468. |
| [31] | Wang Y, Ouyang Z Y, Zheng H, Wang X K, Chen F L, Zeng J. Carbon metabolism of soil microbial communities of restored forests in Southern China. Journal of Soils and Sediments, 2011, 11(5): 789–799. DOI:10.1007/s11368-011-0352-5 |
| [32] | Chen F L, Zheng H, Zhang K, Ouyang Z Y, Lan J, Li H L, Shi Q. Changes in soil microbial community structure and metabolic activity following conversion from native Pinus massoniana plantations to exotic Eucalyptus plantations. Forest Ecology and Management, 2013, 291: 65–72. DOI:10.1016/j.foreco.2012.11.016 |
| [33] | 谭宏伟, 杨尚东, 吴俊, 刘永贤, 熊柳梅, 周柳强, 谢如林, 黄国勤, 赵其国. 红壤区桉树人工林与不同林分土壤微生物活性及细菌多样性的比较. 土壤学报, 2014, 51(3): 575–584. |
| [34] | Laclau J P, Deleporte P, Ranger J, Bouilleti J P, Kazotti G. Nutrient dynamics throughout the rotation of Eucalyptus clonal stands in Congo. Annals of Botany, 2003, 91(7): 879–892. DOI:10.1093/aob/mcg093 |
| [35] | Fierer N, Jackson R B. The diversity and biogeography of soil bacterial communities. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(3): 626–631. DOI:10.1073/pnas.0507535103 |
| [36] | Behera N, Sahani U. Soil microbial biomass and activity in response to Eucalyptus plantation and natural regeneration on tropical soil. Forest Ecology and Management, 2003, 174(1/3): 1–11. |
| [37] | Florentine S K, Fox J E D. Allelopathic effects of Eucalyptus victrix L. on Eucalyptus species and grasses. Allelopathy Journal, 2003, 11(1): 77–83. |
| [38] | Kara Ö, Bolat İ, Çakıroğlu K, Öztürk M. Plant canopy effects on litter accumulation and soil microbial biomass in two temperate forests. Biology and Fertility of Soils, 2008, 45(2): 193–198. DOI:10.1007/s00374-008-0327-x |
| [39] | Zheng H, Chen F L, Ouyang Z Y, Tu N M, Xu W H, Wang X K, Miao H, Li X Q, Tian Y X. Impacts of reforestation approaches on runoff control in the hilly red soil region of Southern China. Journal of Hydrology, 2008, 356(1/2): 174–184. |
| [40] | De Marco A, Gentile A E, Arena C, De Santo A V. Organic matter, nutrient content and biological activity in burned and unburned soils of a Mediterranean maquis area of southern Italy. International Journal of Wildland Fire, 2005, 14(4): 365–377. DOI:10.1071/WF05030 |
| [41] | Neary D G, Klopatek C C, Debano L F, Ffolliott P F. Fire effects on belowground sustainability:a review and synthesis. Forest Ecology and Management, 1999, 122(1/2): 51–71. |
| [42] | Dooley S R, Treseder K K. The effect of fire on microbial biomass:a meta-analysis of field studies. Biogeochemistry, 2012, 109(1/3): 49–61. |
| [43] | Andersson M, Michelsen A, Jensen M, Kjøller A. Tropical savannah woodland:effects of experimental fire on soil microorganisms and soil emissions of carbon dioxide. Soil Biology and Biochemistry, 2004, 36(5): 849–858. DOI:10.1016/j.soilbio.2004.01.015 |
| [44] | Palese A, Giovannini G, Lucchesi S, Dumontet S, Perucci P. Effect of fire on soil C, N and microbial biomass. Agronomie, 2004, 24(1): 47–53. DOI:10.1051/agro:2003061 |
| [45] | 杨尚东, 吴俊, 谭宏伟, 刘永贤, 熊柳梅, 周柳强, 谢如林, 黄国勤, 赵其国. 红壤区桉树人工林炼山后土壤肥力变化及其生态评价. 生态学报, 2013, 33(24): 7788–7797. |