 2018, Vol. 38
2018, Vol. 38文章信息
- 刘从, 田甜, 李珊, 王芳, 梁宇.
- LIU Cong, TIAN Tian, LI Shan, WANG Fang, LIANG Yu.
- 中国木本植物幼苗生长对光照强度的响应
- Growth response of Chinese woody plant seedlings to different light intensities
- 生态学报. 2018, 38(2): 518-527
- Acta Ecologica Sinica. 2018, 38(2): 518-527
- http://dx.doi.org/10.5846/stxb201611012221
-
文章历史
- 收稿日期: 2016-11-01
- 网络出版日期: 2017-09-14





2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
光是影响植物存活、生长和分布的重要生态因子[1], 光照条件在植物整个生活史过程中对植物的生长发育和生理代谢都具有重要影响[2], 也是限制植物生存与生长的主要环境因素[3]。生活型是植物对相同环境长期趋同适应进化的结果[4], 它一方面与植物的生物学特性—遗传特性有关, 另一方面环境因素对植物生活型的影响也是不能低估的[5]。因此, 了解不同生活型植物在不同强度光照条件下的存活和生长能力, 对于预测生态系统对全球变化的响应具有重要作用[6]。
在树木生活史周期中, 幼苗阶段是个体生长最为脆弱、对环境变化最为敏感的时期, 也是个体数量变化最大的时期[7-8]。树木幼苗的存活状况间接响森林群落的物种组成、结构以及稳定性[9]。而幼苗的形态性状如株高和基径, 是植物适应环境变异最直接的外在表现[10]。Böhnke和Bruelheide[11]研究发现不同植物物种对光资源的利用方式是不同的。而江源等[12]对德国南部落叶阔叶林下常见植物的光适应性进行研究, 发现对于光照强度的改变, 植物体能够通过改变生物量分配和形态的变化, 即通常所说的形态可塑性来实现适应。
近年来, 国内外有不少关于植物耐阴性研究的报道, 研究多以森林木本植物作为主要研究对象[13-17]。Zhu等[18]和Adele Muscolo等[19]曾对不同林窗中的木本植物的更新生长做了meta分析, 发现林窗会促进木本植物的更新, 这种促进作用又会因森林类型, 林窗特点, 以及环境因素等的差异而有所不同。这些通过对比林窗内外植物生长来评估植物耐阴性的研究多是在野外自然条件下开展的观测实验, 其研究结果除了受光强影响之外, 还受到许多其他因素, 如森林类型、季节动态、林窗内外在温度、水分等因素的影响, 造成植物耐阴性评估的偏差。本研究通过系统搜集人工控制遮阴实验中光照对幼苗生长影响的研究文献, 将光照强度进行了分级, 采用Meta-analysis方法定量分析不同树种幼苗生长对不同光照强度的响应, 探究植物幼苗对光照的敏感度及其适应性, 以期为森林群落的演替和物种间共存机制研究提供参考, 并为人工管理和培育木本植物幼苗, 进行受损森林生态系统恢复提供科学依据。
1 材料与方法 1.1 文献检索及筛选方法利用国内外网络数据库CNKI、维普、万方、Web of Science、Google Scholar等, 设定关键词(遮阴(或光照)、幼苗、木本、生长等), 采用检索和回溯检索相结合的方法, 检索光照对幼苗形态生长影响研究的国内外文献, 检索年限至2016年, 在进行数据提取之前, 对所有收集到的文献资料进行再次评价, 符合以下条件者视为有效文献:1)文献研究对象为木本植物, 且仅限分布区主要在中国的物种;2)研究所进行的实验为人工控制遮阴实验, 并设有不同的光照梯度;3)数据资料中应包含不同光照梯度下的株高、基径或生物量这三者之一的指标;4)文献资料的数据应是具体的数值(包括图表), 指标包含平均值和样本量, 如有标准差(或标准误)也记录;5)具有重复测量的数据, 只选用最后一次的结果。经过筛选, 最终获得符合标准的有效文献71篇, 共涉及42科的106个树种(附录1)。
1.2 Meta分析统计方法 1.2.1 数据提取本研究通过对已有文献资料结果的归纳总结, 选取了不同光照梯度下幼苗株高、基径、生物量和株高基面积比四个指标来代表幼苗生长情况, 提取相应的实验样本量(N)、平均值(M)、标准误(SE)或标准差(SD)等。如果原文有表格且已给出样本数量、平均值和标准误(或标准差), 则直接抄录;如果为图形, 则通过Engauge digitalize 2.24软件进行数字化转换, 得到相应的数据。
1.2.2 光照处理为了分析不同光照强度的作用, 研究将各篇文献中全光照处理设为对照组, 其他光照强度处理设为实验组。同时为了便于将不同数据进行比较, 按如下标准对实验组的光照强度进行了分组, 共划分为4个等级, 如表 1所示。
| 光照等级 Light level |
光照强度范围 (透光率) Light transmittance |
样本数Sample size | |||
| 株高 Height |
基径 Basal diameter |
生物量 Biomass |
株高基面积比 Height: basal area ratio |
||
| 低光照:Ⅰ | [ 0, 20%) | 83 | 73 | 50 | 73 |
| 中低光照:Ⅱ | [ 20%, 40%) | 90 | 67 | 47 | 67 |
| 中高光照:Ⅲ | [ 40%, 60%) | 104 | 80 | 49 | 80 |
| 高光照:Ⅳ | [ 60%, 80%) | 42 | 37 | 19 | 37 |
利用MetaWin 2.1[20]软件中的随机效应模型对提取数据进行Meta分析。按照Hedges等[21]计算效应值(Effect Size, E)的方法, 将反应比(Responds ration, R)的自然对数作为计算的样本效应值, 其计算公式如下:
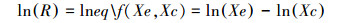
|
式中, Xe和Xc分别为一个独立研究中实验组和对照组的平均值, ln(R)>0表示与对照相比, 实验组光照对幼苗生长起促进作用, 小于0则抑制幼苗生长。
当研究文献中报道了对照组和实验组的平均值, 标准差或标准误, 和实验样本量时, 对实验数据进行meta分析是很方便的[22]。但是在整合分析中, 研究常常会遇到原始文献中测量统计值报道不全面的问题。最常见的是没有清楚地报道样本量及标准差。为了解决此问题, 研究采用Rosenberg等[20]提出的方法, 先用非权重平均法来计算出各指标的效应值[23], 效应值显著性由999次再取样进行检验, 95%置信区间通过MetaWin 2.1软件所整合的自助法(Bootstrap)非加权数据来产生, 然后通过随机检验法来进行异质性检验。如果不同组间的95%置信区间没有重叠, 那么不同组间光照的效应值具有显著差异。如果置信区间包含0, 那么与对照相比, 该处理不能使幼苗生长的变化达到显著水平;若置信区间全部>0, 说明该处理对幼苗生长起促进作用。反之, 则该处理抑制幼苗生长[24]。
2 结果 2.1 不同光照强度下幼苗生长随着光环境的变化, 植物会在形态和结构等方面产生可塑性响应, 来适应变化的光环境。不同光照强度对幼苗株高增长量(QB=33.86, P =0.001;表 2)、基径增长量(QB=25.88, P =0.005;表 2)以及株高基面积比(QB=11.88, P =0.010;表 2)的影响均表现出了显著差异(表 2)。Meta分析结果表明(图 1, 不同处理对应的点代表平均效应值, 横线代表 95%的置信区间), 随光照强度降低, 幼苗的株高、基径和生物量均明显下降, 而株高基面积比的效应值则在20%—40%全光照的情况下达到最大(图 1)。与全光照对照相比, 0—20%的光照强度对幼苗株高增长量有显著的抑制作用, 而60%—80%的光照强度能够显著促进幼苗长高(+0.17, 95% CI:0.107—0.228)。当光照强度小于40%全光照时, 与全光照下幼苗基径增长量(CK)相比, 幼苗基径的生长受到显著抑制。而对于幼苗生物量积累而言, 只有小于20%全光照的光照强度产生了显著抑制作用(-0.30, 95% CI:-0.507—-0.112)(图 1)。

|
| 图 1 不同光照强度下幼苗株高、基径、生物量、株高基面积比效应值分析 Fig. 1 Growth responses of woody plant′s height, basal diameter, biomass and height: basal area ratio Ⅰ、Ⅱ、Ⅲ、Ⅳ分别代表 0—20%, 20%—40%, 40%—60%和60%—80%四个不同的光照强度; 括号中数字为不同处理下样本数 |
| 形态指标 Morphological indicator |
分类变量 Categorical variables |
组间异质性QB Between-group heterogeneity |
P值 P-value |
| 株高Height | 所有树种 | 33.86 | 0.001 |
| 乔木 | 28.09 | 0.002 | |
| 灌木 | 4.76 | 0.128 | |
| 常绿阔叶树种 | 22.28 | 0.001 | |
| 落叶阔叶树种 | 8.68 | 0.090 | |
| 常绿针叶树种 | 5.13 | 0.004 | |
| 基径Basal diameter | 所有树种 | 25.88 | 0.005 |
| 乔木 | 17.60 | 0.014 | |
| 灌木 | 4.96 | 0.025 | |
| 常绿阔叶树种 | 8.82 | 0.037 | |
| 落叶阔叶树种 | 15.89 | 0.127 | |
| 常绿针叶树种 | 2.95 | 0.120 | |
| 生物量Biomass | 所有树种 | 4.89 | 0.169 |
| 乔木 | 4.68 | 0.200 | |
| 灌木 | 3.26 | 0.192 | |
| 常绿阔叶树种 | 14.74 | 0.001 | |
| 落叶阔叶树种 | 3.82 | 0.302 | |
| 常绿针叶树种 | 1.58 | 0.682 | |
| 株高基面积比 | 所有树种 | 11.88 | 0.010 |
| Height:basal area ratio | 乔木 | 8.66 | 0.037 |
| 灌木 | 4.29 | 0.243 | |
| 常绿阔叶树种 | 8.95 | 0.046 | |
| 落叶阔叶树种 | 11.88 | 0.016 | |
| 常绿针叶树种 | 2.31 | 0.592 |
按照休眠芽在不良季节的着生位置, 可将植物划分为乔木、灌木、草本等不同生活型[25]。Meta分析结果表明, 乔木和灌木树种幼苗各生长指标的生长效应均随光照强度降低而呈减小趋势, 而随着光照强度降低, 乔木树种幼苗在株高、基径和生物量累积上受到抑制的程度要大于灌木树种幼苗。对于乔木树种来说, 幼苗在株高增长量(QB=28.09, P =0.002)、基径增长量(QB=17.60, P =0.014)以及株高基面积比(QB=8.66, P=0.037)上在不同光照梯度之间均表现出显著差异。与全光照对照相比, 当光照强度为60%—80%全光照时, 幼苗株高增长量受到显著的促进作用(+0.17, 95%CI:0.109—0.242);而光照强度小于20%全光照时, 显著抑制了幼苗的株高和基径增长量和生物量积累, 并提高了幼苗的株高基面积比。对于灌木树种, 不同强度光照只对幼苗基径增长量产生了显著影响(QB=4.96, P =0.025;表 2), 与全光照对照相比, 当光照强度小于40%全光照时, 遮阴显著抑制幼苗基径增长量;而当光照强度为40%—60%全光照时, 幼苗株高增长量和生物量积累均显著增加(图 2)。

|
| 图 2 不同芽位生活型树种幼苗株高、基径、生物量、株高基面积比效应值分析 Fig. 2 Growth responses of woody plants with different dormant bud position (height, basal diameter, biomass and height: basal area ratio) |
依植物叶片的针叶阔叶和落叶常绿特征, 又可将植物分为落叶阔叶、常绿阔叶和常绿针叶等不同生活型[26]。Meta分析结果表明, 不同光照强度对常绿阔叶树种幼苗株高增长量、基径增长量、生物量累积和株高基面积比的影响均达到了显著水平(表 2)。同时, 落叶阔叶树种幼苗的株高基面积比和常绿针叶树种幼苗的株高增长量在不同光照强度组别之间也存在显著差异(表 2)。
由图 3可以看出, 60%—80%全光照处理能够显著促进幼苗株高增长(图 3)。阔叶树种在 < 40%全光照的条件下, 基径增长量受到显著抑制, 而常绿针叶树种基径增长量受到遮阴的影响较小。幼苗生物量累积对光强的响应与株高增长量基本一致。对于阔叶树种来说, 遮阴对幼苗的株高基面积比产生了显著促进作用, 而针叶树种的株高基面积比受到光强的影响较小(图 3)。

|
| 图 3 不同叶形态生活型树种幼苗株高、基径、生物量、株高基面积比效应值分析 Fig. 3 Growth responses of woody plants with different leaf traits (height, basal diameter, biomass and height: basal area ratio) |
本研究发现, 不同光照强度对幼苗株高和基径增长量以及株高基面积比的影响均表现出了显著差异(表 2, 图 1)。与全光照下幼苗生长相比, 遮阴处理下的幼苗基径增长量受到了相对较大的抑制作用, 而株高增长量则在一定程度上受到了促进。这一现象的产生, 有利于幼苗在林下更快的向上生长以获取足够的光资源, 充分体现了幼苗基径和株高对遮阴环境的适应策略[10]。有研究表明, 当植物受到遮阴时, 幼苗为了最大程度地获取光照, 会将同化的碳更多地用于垂直生长, 而减少用于基径增长的碳[10, 27-28]。
本研究结果表明, 60%—80%全光照条件下, 幼苗的株高增长量显著高于对照(图 1)。这与之前的许多研究结果是一致的:即中等强度的光照有利于木本植物幼苗的生长[29-30], 而过强的光照对幼苗生长具有抑制作用[30-34]。这是因为光是木本植物光合作用的能量来源, 光强增加有利于植物的生长和生物量积累[35-36]。但是木本植物幼苗的生物量积累存在着一个最佳光强范围[36], 处于幼年时期的树种对光能利用和保护的机制还未完善, 类似日常午间的高光强大多超出了幼苗所能利用的光能范围, 进而导致植物发生光抑制[37], 因此, 全光照并不是幼苗生长的最佳光照强度。
3.2 光照对乔木和灌木树种幼苗生长的影响由研究结果来看, 乔木和灌木树种幼苗生长受到遮阴处理的影响, 但相对而言乔木树种比灌木树种对光强更加敏感(图 2)。乔木常常处于森林的林冠层, 很多乔木种类也常常是喜阳植物。许多研究发现乔木对遮阴处理比较敏感, 如闫兴富等[38]对暴马丁香和臭椿幼苗的研究表明, 充分的光照条件是暴马丁香幼苗和臭椿幼苗得以良好成长的保证, 而徐飞等[3]也发现, 麻栎和刺槐在弱光条件下生长均受到光线不足的限制。灌木树种常常是生长于林下层, 并且具有分布广泛、萌生力强、适生范围广的特点[39], 可能是其具有较强的耐阴性的原因。Valladares和Niinemets[40]研究也发现, 很多灌木对遮阴并不敏感, 在中等光照强度下生长最好, 达到最大生物量。
3.3 光照对不同叶形态生活型树种幼苗生长的影响叶片是植物进化过程中对环境变化较敏感且可塑性较大的器官, 叶片可通过多种途径改变其外在或内在特征, 来适应变化的光环境[41]。本研究发现, 与针叶树种相比, 阔叶树种幼苗的基径增长量和株高基面积比更容易受到遮阴的影响(图 3)。通常认为, 松科植物等针叶树种往往具有很强的向阳性[42], 其生长和树形可塑性应该对遮阴处理更为敏感。而我们的结果表明, 阔叶树种的株高基面积比对遮阴处理比针叶树种更为敏感, 表现出对光资源变化更好的适应性。这一结果也可以在一定程度上解释在很多自然林中, 针叶树种的幼苗更新常常较阔叶树种更为困难, 在群落演替后期逐步被替代的现象。
本研究结果还发现, 常绿阔叶树种幼苗的株高增长量在重度遮阴条件下( < 20%全光照)受到抑制, 而落叶阔叶树种株高增长量并未受到明显抑制, 表明落叶树种较常绿树种有较强的耐阴性(图 3)。这一结果与一些前人的研究结果并不一致, Böhnke和Bruelheide[11]的研究发现, 在相同遮阴条件下, 落叶树种对光照的敏感性常常高于常绿树种。杨波[43]通过对亚热带森林树种遮阴抑制率的计算也发现, 与常绿树种相比, 落叶树种的遮阴抑制率较高, 常常表现出对光更强的依赖性。本研究表明, 落叶阔叶树种对遮阴的适应范围比我们通常认为的大得多, 而对光照条件的较强适应性也可以部分解释落叶树种在我国自然林中分布较广的原因。
| [1] | Rozendaal D M A, Hurtado V H, Poorter L. Plasticity in leaf traits of 38 tropical tree species in response to light; relationships with light demand and adult stature. Functional Ecology, 2006, 20(2): 207–216. DOI:10.1111/fec.2006.20.issue-2 |
| [2] | Callahan D M, Munday J N, Atwater H A. Solar cell light trapping beyond the ray optic limit. Nano Letters, 2012, 12(1): 214–218. DOI:10.1021/nl203351k |
| [3] | 徐飞, 郭卫华, 徐伟红, 王仁卿. 不同光环境对麻栎和刺槐幼苗生长和光合特征的影响. 生态学报, 2010, 30(12): 3098–3107. |
| [4] | 刘守江, 苏智先, 张璟霞, 胡进耀. 陆地植物群落生活型研究进展. 四川师范学院学报:自然科学版, 2003, 24(2): 155–159. |
| [5] | 徐廷志. 植物生活型及其分类系统的初步研究. 大自然探索, 1986(2): 77–81. |
| [6] | Valladares F, Niinemets V. Shade tolerance, a key plant feature of complex nature and consequences. Annual Review of Ecology, Evolution, and Systematics, 2008, 39: 237–257. DOI:10.1146/annurev.ecolsys.39.110707.173506 |
| [7] | 李俊清, 臧润国, 蒋有绪. 欧洲水青冈(Fagus sylvatical L.)构筑型与形态多样性研究. 生态学报, 2001, 21(1): 151–155. |
| [8] | 韩有志, 王政权. 森林更新与空间异质性. 应用生态学报, 2002, 13(5): 615–619. |
| [9] | Modrzyński J, Chmura D J, Tjoelker M G. Seedling growth and biomass allocation in relation to leaf habit and shade tolerance among 10 temperate tree species. Tree Physiology, 1983, 35(8): 879–893. |
| [10] | 洪明, 郭泉水, 聂必红, 康义, 裴顺祥, 金江群, 王祥福. 三峡库区消落带狗牙根种群对水陆生境变化的响应. 应用生态学报, 2011, 22(11): 2829–2835. |
| [11] | Böhnke M, Bruelheide H. How do evergreen and deciduous species respond to shade?-Tolerance and plasticity of subtropical tree and shrub species of South-East China. Environmental and Experimental Botany, 2013, 87: 179–190. DOI:10.1016/j.envexpbot.2012.09.010 |
| [12] | 江源. 德国南部落叶阔叶林下常见植物的光适应性研究——以德国Kraichtal地区落叶阔叶林为例. 植物学报, 2001, 43(9): 960–966. |
| [13] | Lusk C H. Long-lived light-demanding emergents in southern temperate forests:the case of Weinmannia trichosperma (Cunoniaceae) in Chile. Plant Ecology, 1999, 140(1): 111–115. DOI:10.1023/A:1009764705942 |
| [14] | Poorter L. Growth responses of 15 rain-forest tree species to a light gradient:the relative importance of morphological and physiological traits. Functional Ecology, 1999, 13(3): 396–410. DOI:10.1046/j.1365-2435.1999.00332.x |
| [15] | Malavasi U C, Malavasi M M. Leaf characteristics and chlorophyll concentration of Schyzolobium parahybum and Hymenaea stilbocarpa seedlings grown in different light regimes. Tree Physiology, 2001, 21(10): 701–703. DOI:10.1093/treephys/21.10.701 |
| [16] | Lusk C H, Del Pozo A. Survival and growth of seedlings of 12 Chilean rainforest trees in two light environments:gas exchange and biomass distribution correlates. Austral Ecology, 2002, 27(2): 173–182. DOI:10.1046/j.1442-9993.2002.01168.x |
| [17] | Bloor J M G, Grubb P J. Growth and mortality in high and low light:trends among 15 shade-tolerant tropical rain forest tree species. Journal of Ecology, 2003, 91(1): 77–85. DOI:10.1046/j.1365-2745.2003.00743.x |
| [18] | Zhu J J, Lu D L, Zhang W D. Effects of gaps on regeneration of woody plants:a meta-analysis. Journal of Forestry Research, 2014, 25(3): 501–510. DOI:10.1007/s11676-014-0489-3 |
| [19] | Muscolo A, Bagnato S, Sidari M, Mercurio R. A review of the roles of forest canopy gaps. Journal of Forestry Research, 2014, 25(4): 725–736. DOI:10.1007/s11676-014-0521-7 |
| [20] | Rosenberg M S, Adams D C, Gurevitch J. MetaWin:Statistical Software for Meta-Analysis. Version 2.0. Massachusetts, USA:Sinauer Associates Sunderland, 2000. |
| [21] | Hedges L V, Gurevitch J, Curtis P S. The Meta-Analysis of Response Ratios in Experimental Ecology. Ecology, 1999, 80(4): 1150–1156. DOI:10.1890/0012-9658(1999)080[1150:TMAORR]2.0.CO;2 |
| [22] | Gurevitch J, Hedges L V. Statistical issues in ecological meta-analyses. Ecology, 1999, 80(4): 1142–1149. DOI:10.1890/0012-9658(1999)080[1142:SIIEMA]2.0.CO;2 |
| [23] | Johnson D W, Curtis P S. Effects of forest management on soil C and N storage:meta analysis. Forest Ecology and Management, 2001, 140(2/3): 227–238. |
| [24] | Lin Y M, Dimitrakopoulos C, Jenkins K A, Farmer D B, Chiu H Y, Grill A. 100-GHz transistors from wafer-scale epitaxial graphene. Science, 2010, 327(5966): 662–662. DOI:10.1126/science.1184289 |
| [25] | 李博. 生态学. 北京: 高等教育出版社, 2000. |
| [26] | 高贤明, 陈灵芝. 植物生活型分类系统的修订及中国暖温带森林植物生活型谱分析. 植物学报, 1998, 40(6): 553–559. |
| [27] | 刘泽彬, 程瑞梅, 肖文发, 郭泉水, 王娜. 遮荫对中华蚊母树苗期生长及光合特性的影响. 林业科学, 2015, 51(2): 129–136. |
| [28] | 薛思雷, 王庆成, 孙欣欣, 张命军. 遮荫对水曲柳和蒙古栎光合、生长和生物量分配的影响. 植物研究, 2012, 32(3): 354–359. |
| [29] | Veenendaal E M, Swaine M D, Agyeman V K, Blay D, Abebrese I K, Mullins C E. Differences in plant and soil water relations in and around a forest gap in West Africa during the dry season may influence seedling establishment and survival. Journal of Ecology, 1996, 84(1): 83–90. DOI:10.2307/2261702 |
| [30] | 许邦锐, 张明生, 李祥栋, 曹然, 刘诗雅, 李立青. 光照强度对狐臭柴植株生长及叶片生理生化指标的影响. 中南林业科技大学学报, 2013, 33(12): 82-85, 95-95. |
| [31] | 李威. 东北红豆杉幼树和幼苗光合生理适应性研究[D]. 哈尔滨: 东北林业大学, 2012. |
| [32] | 刘鹏, 康华靖, 张志详, 徐根娣, 张争艳, 陈子林, 廖承川, 陈卫新. 香果树(Emmenopterys henryi)幼苗生长特性和叶绿素荧光对不同光强的响应. 生态学报, 2008, 28(11): 5656–5664. DOI:10.3321/j.issn:1000-0933.2008.11.051 |
| [33] | 徐燕, 张远彬, 乔匀周, 李青超, 王开运. 光照强度对川西亚高山红桦幼苗光合及叶绿素荧光特性的影响. 西北林学院学报, 2007, 22(4): 1-4, 12-12. |
| [34] | 李利, 张希明. 光照对胡杨幼苗定居初期生长状况和生物量分配的影响. 干旱区研究, 2002, 19(2): 31–34. |
| [35] | 蒋高明. 植物生理生态学. 北京: 高等教育出版, 2004. |
| [36] | 陈圣宾, 宋爱琴, 李振基. 森林幼苗更新对光环境异质性的响应研究进展. 应用生态学报, 2005, 16(2): 365–370. |
| [37] | Jiang C D, Li P M, Gao H Y, Zou Q, Jiang G M, Li M H. Enhanced photoprotection at the early stages of leaf expansion in field-grown soybean plants. Plant Science, 2005, 168(4): 911–919. DOI:10.1016/j.plantsci.2004.11.004 |
| [38] | 闫兴富, 方苏, 杜茜, 周立彪. 遮荫对暴马丁香种子萌发及幼苗生长的影响. 甘肃农业大学学报, 2010, 45(1): 104–110. |
| [39] | 侯学煜. 中华人民共和国植被图. 北京: 地图出版社, 1982. |
| [40] | Valladares F, Wright S J, Lasso E, Kitajima K, Pearcy R W. Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest. Ecology, 2000, 81(7): 1925–1936. DOI:10.1890/0012-9658(2000)081[1925:PPRTLO]2.0.CO;2 |
| [41] | Poorter L, Bongers F. Leaf traits are good predictors of plant performance across 53 rain forest species. Ecology, 2006, 87(7): 1733–1743. DOI:10.1890/0012-9658(2006)87[1733:LTAGPO]2.0.CO;2 |
| [42] | Landhäusser S M, Lieffers V J. Photosynthesis and carbon allocation of six boreal tree species grown in understory and open conditions. Tree Physiology, 2001, 21(4): 243–250. DOI:10.1093/treephys/21.4.243 |
| [43] | 杨波. 影响亚热带森林生物多样性与生态系统功能关系的关键因子研究[D]. 北京: 中国科学院植物研究所, 2014. |