 2018, Vol. 38
2018, Vol. 38文章信息
- 黄慧琴, 刘涛, 石金泽, 刘鹏, 张微微.
- HUANG Huiqin, LIU Tao, SHI Jinze, LIU Peng, ZHANG Weiwei.
- 蓝冠噪鹛繁殖期生境选择特征分析
- Habitat selection of the Blue-Crowned Laughingthrush during the breeding season
- 生态学报. 2018, 38(2): 493-501
- Acta Ecologica Sinica. 2018, 38(2): 493-501
- http://dx.doi.org/10.5846/stxb201609261942
-
文章历史
- 收稿日期: 2016-09-26
- 网络出版日期: 2017-09-14





2. 东北林业大学野生动物资源学院, 哈尔滨 150040;
3. 江西省林业科学院, 南昌 330013
2. College of Wildlife Resources Conservation, Northeast Forestry University, Harbin 150040, China;
3. Jiangxi Academy of Forestry, Nanchang 330013, China
生境是为动物提供资源、庇护所及筑巢位置的生存空间, 生境的质量会直接影响种间竞争和种内竞争的强度, 影响疾病和捕食的代价。因此, 动物会有规律地选择适宜的生境。动物的生境选择是为了对资源实现最优化利用, 本质上是进化策略的体现[1]。
近年来, 人们逐渐意识到鸟类栖息地选择可以发生在多个尺度上, 单一尺度并不能完全反映其生境选择的影响因子[2]。如今, 鸟类繁殖栖息地选择研究通常会在不同的尺度下进行, 即宏栖息地尺度的某一物种的地理分布区和某一种群的社群巢域的研究以及微栖息地尺度的巢域范围内动物选择的不同生境类型的实际环境条件[3]。曹铭昌等从景观尺度、斑块尺度和微生境尺度对影响丹顶鹤(Grus japonensis)生境选择的因子进行了研究, 结果表明, 景观尺度对丹顶鹤生境选择的影响远远大于斑块尺度和微生境尺度[4]。李东来等对震旦鸦雀(Paradoxornis heudei)进行了多尺度的巢址选择研究[5]。Greenberg等对3种柳莺非繁殖期觅食行为和生境选择的研究, 表明生态位的差异使得柳莺对微生境的选择产生了差异[6]。
国外一些鸟类学者将栖息地选择研究与鸟类的资源管理和濒危物种保护相结合, 例如北美和欧洲的鸟类学家对一些雉科鸟类的研究应用与资源管理密切相关。Kostrzewa对蜂鹰(Pernis aqivorus)、苍鹰(Accipiter gentilis)和普通鵟(Buteo buteo)营巢位置的研究对猛禽保护提供了数据支持[7]。Doucette对澳洲裸鼻鸱(Aegotheles cristatus)巢域和活动范围的研究, 结果表明澳洲裸鼻鸱各季节的活动范围没有明显差异, 并且对巢域有很高的忠诚度[8]。Murray和Best对黄喉地莺(Geothlypis trichas)的生境选择和繁殖成功率的研究发现, 黄喉地莺选择草本盖度更高的位置筑巢, 然而繁殖成功率与草本盖度却呈负相关[9]。我国对鸟类生境选择的研究主要针对鹤料、雉科等一些珍稀物种。例如白鹤(Grus leucogeranus)[10]、丹顶鹤(Grus japonensis)[4, 11]、白头鹤(Grus monacha)[12]、中华秋沙鸭(Mergus squamatus)[13]和白冠长尾雉(Syrmaticus reevesii)等[14-16]。
蓝冠噪鹛(Garrulax courtosis)是雀形目(Passeriformes)画眉科(Timaliidae)噪鹛属(Garrulax)的鸟类。国际鸟盟(BirdLife International)将蓝冠噪鹛列入全球濒危鸟类名录, 并将它的受胁等级定为极危(Critically Endangered, CR), 关于蓝冠噪鹛的研究主要集中在繁殖期行为描述和繁殖生境的描述[17-20]。其相关生境选择特征研究尚未见报道, 相关资料的缺乏十分不利于其栖息地的保护, 本研究拟从多尺度探讨蓝冠噪鹛生境选择特征, 明晰影响其繁殖地选择的生境因子, 相关研究结果可为其引种、潜在栖息地保护及迁地保护等提供基础资料。
1 研究地概况江西省婺源县位于江西省东北部, 地理坐标为117°22′—118°12′E, 29°01′—29°34′N, 地处赣、浙、皖三省交界地带, 总面积2947.51km2。婺源县北部以五龙山脉承接黄山, 南靠怀玉山脉和武夷山脉, 境内地貌以中低山地和丘陵为主, 山地占总面积的83.09%。平均海拔100—150m。婺源境内天然水系发达, 河流溪涧众多, 最主要的河流乐安江是饶河的一条支流, 发源于婺源县北部的大庾山、五龙山, 其支流众多, 遍布婺源各地。
自1993年起, 婺源县政府陆续将传统保留下来的村落风水林划建为自然保护区, 这些风水林以阔叶林为主, 多为常绿阔叶林和针阔混交林, 少数是落叶阔叶林。常见植物有朴树(Celtis tetrandra)、枫香(Liquidambar formosana)、枫杨(Pterocarya stenoptera)、苦槠(Castanopsis sclerophylla)、杉木(Cunninghamia lanceolata)、小叶青冈(Cyclobalanopsis myrsinifolia)、毛竹(Phyllostachys heterocycla)等。
2 研究和分析方法 2.1 野外调查方法2013—2015年, 每年的繁殖期(4—7月)在婺源县寻找蓝冠噪鹛繁殖小群并确定其活动区域, 并用GPS定位;在各繁殖点内搜索蓝冠噪鹛巢, 对巢进行编号, 并用GPS定位。
(1) 斑块尺度
在蓝冠噪鹛繁殖地寻找并记录繁殖点位置, 跟踪观察其繁殖期的活动区域, 使用GPS定位, 结合Google Earth软件将每个繁殖点区域标注在地图上。对照区随机选择距离蓝冠噪鹛繁殖点5km以外的没有蓝冠噪鹛繁殖的阔叶林。测量繁殖区与对照区海拔, 每个斑块形状重心点距水源地、距干扰源和距最近的山地的距离。使用Fragstats 4.2软件计算斑块面积、周长、形状指数、距最近斑块距离等斑块水平指数数据。
形状指数的计算方法为:
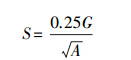
|
其中, S为形状指数, A为斑块面积, G为斑块周长。此公式适用于近似于正方形的斑块。蓝冠噪鹛的繁殖点大多为近似正方形或近似长方形, 适用此公式。
比较繁殖地和对照区斑块尺度各因子的差异, 分析蓝冠噪鹛偏爱的繁殖生境。
(2) 微生境尺度
繁殖点B是最为稳定, 也是繁殖数量最多的繁殖点。因此将其作为微生境尺度生境选择的研究点。
寻找蓝冠噪鹛的巢, 标记编号, 测量并记录巢高、巢上郁闭度、巢下郁闭度、海拔、距水源距离、距干扰源(村庄)距离, 同时记录营巢树种, 测量其胸径和树高, 仅对树高5m以上的乔木进行统计。
在蓝冠噪鹛的集群营巢区域选取4个10m×10m的巢址乔木样方, 在没有蓝冠噪鹛繁殖且距离营巢区较远的区域选取4个同样大小的对照样方, 统计和测量样方中的乔木种类、树高、胸径, 以及海拔、距干扰源距离、距水源距离。以巢为中心, 在半径5m的范围内随机选取2个1m×1m的草本样方, 在没有蓝冠噪鹛繁殖且距离营巢区较远的区域随机选取同样数量的草本样方, 测量样方内的草本种数、草本数量、草本高度、盖度。
海拔:用手持GPS(UniStrong)测定, 将海拔分为:<80m、80—90m和≥90m 3个等级。
距干扰源距离:样方中心距最近居民点的距离, 分为:<100m、100—200m和≥200m 3个等级。
距水源距离:样方中心距最近河流或池塘等水域的距离, 分为:<40m、40—80m和≥80m 3个等级。
树种:样区调查到树种共14种, 分别用1—14数字对各树种进行赋值。
树高:用估测法, 分为:<10m、10—20m、20—30m和≥30m 4个等级。
树径:采用卷尺测量, 分为:<20cm、20—40cm、40—60cm、60—80cm和≥80cm 5个等级。
草本种数:样区调查到的草种数, 分为:<5种、5—10种、10—15种、15—20种和≥20种5个等级。
草本密度:用样方中的草本株数表示, 分为:<1000株、1000—3000株、3000—6000株和≥6000株4个等级。
草本高度:采用卷尺测量, 分为:<20cm、20—40cm和40—60cm 3个等级。
草本盖度:用估测法, 分为:<30%、30%—60%和60%—90% 3个等级。
计算巢址样方和对照样方间各因素的差异, 使用资源选择函数以及Vanderploeg与Scavia选择系数两种方法对蓝冠噪鹛繁殖微生境特征进行分析。
2.2 数据分析 2.2.1 资源选择函数资源选择函数是基于Logistic方程而开发的[21]。Logistic方程选择自变量时, 要求各变量之间相互独立[22-23]。由于动物对资源的选择受食物、隐蔽物、水源、地形地貌等多个生境因子的影响, 资源选择函数可表示为多个独立生境变量的线性对数模型:
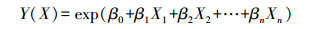
|
式中:X为不同的独立生境变量, β表示选择系数。那么, 物种对生境的选择概率为:
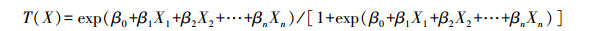
|
当T(X)的取值为1或0时, 即表示选择或不选择, 选择系数β可以由逻辑斯蒂回归系数来估计[24]。
2.2.2 Vanderploeg和Scavia选择系数使用Vanderploeg和Scavia选择系数Wi和选择指数Ei [25]作为衡量蓝冠噪鹛繁殖微生境喜好程度的指标, 其计算方法如下:
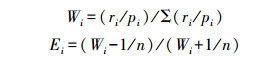
|
式中:Wi为选择系数;Ei为选择指数;i为特征值;n为特征值总数;pi为环境中具i特征的样方数;ri为蓝冠噪鹛繁殖区所选择具有i特征的样方数。Ei=-1表示不选择(用N表示), -1<Ei<0表示回避(用NP表示), Ei=0表示随机选择(用R表示), 接近0也表示随机选择。0<Ei<1表示选择(用S表示), Ei=1表示高度选择(用SP表示)。
所有数据都在SPSS 19.0中进行处理和分析。采用Kolmogorov Smirnov Z-检验对数字型因子进行正态分布检验, 对不符合正态分布的因子做log转换后, 再做正态性检验。当数据符合正态分布时, 采用单因素方差分析LSD(方差齐性)或Thamhane′s T2(方差不齐)多重比较方法, 找出差异显著性因子。
3 结果 3.1 斑块尺度经过3年共调查了9处蓝冠噪鹛的繁殖点。所有繁殖点海拔基本在100m以下, 巢区距离水源最远不足1km, 且多在200m以内, 距离干扰源均在250m以内。各斑块之间的距离在2—40km范围, 平均为(5.63±1.02)km, 基本超出繁殖期蓝冠噪鹛各繁殖点内的最远活动距离。斑块面积从0.05hm2到25.7hm2不等, 形状多样, 区域边缘不整齐。繁殖点A、C、F、G、I散落在村庄中, 其中除C和I近似圆形生境外,其余近似方形生境, 繁殖点B、D都是跨越河流的两个方形区域组成的生境, 繁殖点E是沿着河流的条状生境, 繁殖点H是村庄旁边的近似圆形斑块生境。各繁殖点的景观参数见表 1。
| 繁殖点 Sites | 海拔 Elevation /m | 距主要 水源距离 Distance to water /m | 距干扰 源距离 Distance to disturbance /m | 距山地距离 Distance to mountain/m | 面积 Area/hm2 | 周长 Perimeter /km | 距最近斑 块距离 Distance to nearest plaque/ km | 形状指数 Shape Index |
| A | 85 | 50 | 75 | 420 | 4.82 | 1.11 | 2.59 | 1.27 |
| B | 84 | 80 | 220 | 500 | 25.7 | 2.61 | 2.59 | 1.29 |
| C | 82 | 350 | 20 | 420 | 1.61 | 0.5 | 3.7 | 0.99 |
| D | 49 | 75 | 220 | 550 | 0.62 | 0.43 | 7.03 | 1.36 |
| E | 60 | 65 | 120 | 700 | 20.08 | 2.28 | 5.83 | 1.27 |
| F | 60 | 120 | 55 | 380 | 0.05 | 1.01 | 5.83 | 3.52 |
| G | 112 | 760 | 20 | 450 | 6.06 | 1.41 | 3.7 | 1.43 |
| H | 90 | 140 | 70 | 300 | 1.17 | 0.45 | 12.35 | 1.04 |
| I | 53 | 330 | 50 | 250 | 0.71 | 0.35 | 7.03 | 1.04 |
| 均值 Average | 75.0±6.91 | 218.0±77.33 | 94.4±25.76 | 441.1±44.64 | 6.81±3.14 | 1.13±0.23 | 5.63±1.02 | 1.47±0.26 |
将繁殖区与对照样区的7个因子进行比较, 结果表明海拔、距干扰源距离、距山地距离具有显著差异(海拔:t=-2.805, df=16, P=0.013;距干扰源距离:t=-3.040, df=70, P=0.003;距山地距离:t=-2.683, df=50.498, P=0.009)。繁殖点比对照区的海拔更低, 且距离干扰源和山地更近(表 2)。
| 因子Factors | F | P | t | df | P |
| 海拔Elevation/m | 0.003 | 0.959 | -2.805 | 16 | 0.013* |
| 距水源距离Distance to water/m | 5.263 | 0.025 | -1.950 | 46.739 | 0.057 |
| 距干扰源距离Distance to disturbance/m | 0.240 | 0.626 | -3.040 | 70 | 0.003** |
| 距山地距离Distance to mountain/m | 14.422 | 0.000 | -2.683 | 50.498 | 0.009** |
| 面积Area/hm2 | 1.264 | 0.277 | 0.128 | 16 | 0.900 |
| 周长Perimeter/km | 2.863 | 0.110 | 0.308 | 16 | 0.762 |
| 形状指数Shape Index | 0.007 | 0.934 | 0.755 | 16 | 0.461 |
| **:差异在0.01水平上显著, *:差异在0.05水平上显著 | |||||
(1) 巢址样方和对照样方比较
将巢址样方与对照样方的10个因子进行比较, 结果(表 3)表明仅草本密度、高度和树种三者差异显著(草本密度:t=4.230, df=78.108, P < 0.001;草本高度:t=-2.663, df=105, P=0.009;树种:t=-1.991, df=105, P=0.049)。
| 因子Factors | 巢区Nest area | 对照区Control area | t | df | P | ||||||
| 均值 | 标准误 | 下限 | 上限 | 均值 | 标准误 | 下限 | 上限 | ||||
| 海拔E Elevation | 81.89 | 1.186 | 79.585 | 84.211 | 79.57 | 0.203 | 79.200 | 79.975 | 1.931 | 63.490 | 0.058 |
| 树高TH Tree height | 22.28 | 0.615 | 21.015 | 23.403 | 22.78 | 0.663 | 21.480 | 24.146 | -0.561 | 105 | 0.576 |
| 胸径DBH Diameter at breast height | 46.57 | 2.775 | 41.161 | 52.140 | 50.04 | 3.604 | 43.137 | 57.358 | -0.764 | 105 | 0.447 |
| 干扰距离DD Distance to disturbance | 133.03 | 8.922 | 115.023 | 150.521 | 145.30 | 11.648 | 121.878 | 167.457 | -0.833 | 88.390 | 0.407 |
| 水源距离DW Distance to water | 39.85 | 2.263 | 35.556 | 44.321 | 36.74 | 4.856 | 27.752 | 47.114 | 0.568 | 65.721 | 0.572 |
| 树种TS Tree species | 1.98 | 0.110 | 1.771 | 2.209 | 2.41 | 0.200 | 2.052 | 2.837 | -1.991 | 105 | 0.049* |
| 草本种类HS Herb species | 9.36 | 0.369 | 8.687 | 10.105 | 9.36 | 0.369 | 8.687 | 10.105 | -0.456 | 105 | 0.649 |
| 草本密度HD Herb density | 2309.18 | 255.720 | 1844.253 | 2837.466 | 1109.83 | 105.533 | 897.112 | 1326.318 | 4.230 | 78.108 | 0.000** |
| 草本高度HH Herb height | 20.27 | 1.064 | 18.292 | 22.392 | 24.67 | 1.237 | 22.249 | 27.067 | -2.663 | 105 | 0.009** |
| 草本盖度HC Herb coverage | 0.73 | 0.030 | 0.675 | 0.791 | 0.64 | 0.039 | 0.565 | 0.713 | 1.924 | 105 | 0.057 |
| **:差异在0.01水平上显著,*:差异在0.05水平上显著 | |||||||||||
(2)资源选择函数:
为控制各变量之间的自相关性, 在拟合逻辑斯蒂方程之前对9个生境因子进行相关分析, 由于9个变量中各个变量均不符合正态分布(Kilmogorov-Smirnov), 因此使用Spearman秩相关检验, 生境因子两两比较相关系数绝对值均小于0.5(表 4), 因此把所有因子输入逻辑斯蒂回归模型, 对所有参数都进行标准化, 以使参数间没有量纲的差异, 计算出来的选择系数可相互比较, 之后根据模型给出的回归系数显著性水平高低来判断各因子的取舍。
| 因子Factors | E | TH | DBH | DD | DW | HS | HD | HH | HC |
| E | 1 | ||||||||
| TH | -0.128 | 1 | |||||||
| DBH | -0.187* | 0.393** | 1 | ||||||
| DD | -0.257** | 0.243* | 0.316** | 1 | |||||
| DW | 0.211* | -0.197* | -0.407** | -0.182 | 1 | ||||
| HS | -0.236* | 0.186* | 0.166* | 0.358** | -0.219* | 1 | |||
| HD | 0.18 | 0.055 | -0.043 | 0.015 | 0.002 | 0.191** | 1 | ||
| HH | -0.171 | -0.02 | 0.056 | -0.214* | 0.074 | 0.117 | 0.057 | 1 | |
| HC | -0.033 | 0.051 | -0.025 | -0.207* | 0.342** | 0.134 | 0.283** | 0.321** | 1 |
| **:相关性在0.01水平上显著, *:相关性在0.05水平上显著 | |||||||||
回归模型显示仅有草本密度和草本高度2项的回归系数达到显著水平(表 5)。因此用草本密度和草本高度两个变量建立的资源选择函数模型为:
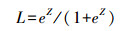
|
| 因子Factors | 选择系数 Regression coefficient | 标准误 SE | Wald卡方检验 χ2 | 显著性 Significance |
| 常数项Constant | 0.495 | 0.293 | 2.847 | 0.092 |
| HD | 1.260 | 0.519 | 5.908 | 0.015* |
| HH | -0.775 | 0.284 | 7.432 | 0.006** |
| E | 0.160 | 0.266 | 0.364 | 0.546 |
| TH | -0.272 | 0.320 | 0.723 | 0.395 |
| DBH | 0.197 | 0.325 | 0.369 | 0.543 |
| DD | -0.141 | 0.260 | 0.293 | 0.588 |
| DW | 0.128 | 0.263 | 0.237 | 0.627 |
| HS | 0.057 | 0.225 | 0.064 | 0.800 |
| HC | 0.512 | 0.295 | 3.008 | 0.083 |
| **:差异在0.01水平上显著, *:差异在0.05水平上显著 | ||||
其中Z=0.495+1.26×草本密度-0.775×草本高度, e为自然数, L表示生境选择概率。模型预测率为64.06%, 复相关系数为0.087(Nagelkerke R=0.116)。偏差分析值为252.65。结果显示选择概率与草本密度呈正相关, 与草本高度呈负相关, 说明蓝冠噪鹛巢址选择倾向于在草密度较大、草较低矮的生境中。
(3) Vanderploeg和Scavia选择系数
选择指数的结果为蓝冠噪鹛喜在朴树、枫杨和枫香3种树且树径较粗(40—80cm)的树筑巢, 筑巢树高偏好20m以上, 在筑巢区林下则偏好草本高度较低(20cm以下)及草本盖度较高(60%—90%)的生境。蓝冠噪鹛倾向于选择距水源40—80m的位点活动, 承受人类干扰的距离为100—200m(表 6)。
| 因子 Factors | i | Pi | ri | Wi | Ei | 生境偏好 Habitat preference |
| E | <80 | 46 | 20 | 0.22 | -0.21 | NP |
| 80—90 | 56 | 32 | 0.28 | -0.08 | NP | |
| ≥90 | 9 | 9 | 0.50 | 0.20 | S | |
| TH | <10 | 12 | 0 | 0 | -1 | N |
| 10—20 | 69 | 17 | 0.24 | -0.03 | NP | |
| 20—30 | 113 | 39 | 0.33 | 0.14 | S | |
| ≥30 | 11 | 5 | 0.43 | 0.27 | S | |
| DBH | <20 | 32 | 5 | 0.12 | -0.27 | NP |
| 20—40 | 64 | 19 | 0.22 | 0.05 | R | |
| 40—60 | 60 | 21 | 0.26 | 0.13 | S | |
| 60—80 | 35 | 12 | 0.25 | 0.12 | S | |
| ≥80 | 20 | 4 | 0.15 | -0.15 | NP | |
| DD | <100 | 38 | 19 | 0.32 | -0.02 | NP |
| 100—200 | 46 | 33 | 0.46 | 0.16 | S | |
| ≥200 | 27 | 9 | 0.22 | -0.22 | NP | |
| DW | <40 | 68 | 33 | 0.35 | 0.03 | R |
| 40—80 | 36 | 27 | 0.54 | 0.24 | S | |
| ≥80 | 7 | 1 | 0.10 | -0.53 | NP | |
| TC | 1—3 | 236 | 117 | 0.88 | 0.63 | S |
| 4—6 | 15 | 1 | 0.12 | -0.26 | NP | |
| 7—9 | 6 | 0 | 0 | -1 | N | |
| 10—12 | 8 | 0 | 0 | -1 | N | |
| 13—14 | 2 | 0 | 0 | -1 | N | |
| NCS | <5 | 2 | 1 | 0.2 | 0 | R |
| 5—10 | 132 | 66 | 0.2 | 0 | R | |
| 10—15 | 56 | 28 | 0.2 | 0 | R | |
| 15—20 | 2 | 1 | 0.2 | 0 | R | |
| HD | <1000 | 107 | 44 | 0.21 | -0.10 | NP |
| 1000—3000 | 63 | 37 | 0.29 | 0.08 | R | |
| 3000—6000 | 19 | 11 | 0.29 | 0.07 | R | |
| ≥6000 | 12 | 5 | 0.21 | -0.09 | NP | |
| HH | <20 | 91 | 64 | 0.48 | 0.19 | S |
| 20—40 | 94 | 30 | 0.22 | -0.20 | NP | |
| 40—60 | 7 | 3 | 0.30 | -0.06 | NP | |
| HC | <30 | 31 | 10 | 0.25 | -0.14 | NP |
| 30—60 | 43 | 15 | 0.27 | -0.10 | NP | |
| 60—90 | 118 | 72 | 0.48 | 0.18 | S | |
| i:特征值Eigenvalue; Pi:环境中具i特征的样方数the number of sample which have i characteristics in all survey areas; ri:蓝冠噪鹛繁殖区所选择具有i特征的样方数the number of sample which have i characteristics in breeding area of Blue-crowned Laughingthrush; Wi:选择系数Selectivity coefficient; Ei:选择指数Electivity index; S:选择Selected;NP:回避Not preferred;R:随机选择Random selection;N:不选择 Not selected | ||||||
3年期间共调查蓝冠噪鹛繁殖点9个, 总面积60.82km2, 破碎度为0.147。斑块的形状对于动物的扩散和觅食具有重要作用[26]。一般来说, 大的斑块比小型斑块承载更多的物种数, 但小型斑块的资源可能不足以吸引大型捕食动物, 从而使小型斑块可能成为某些小型物种的避难所[27]。从巢捕食风险的角度讲小型鸟类对片段化的生境更为敏感[28]。蓝冠噪鹛生境呈明显斑块状, 且斑块面积非常小, 有研究表明巢捕食率及捕食者活动率与岛屿面积呈正相关, 且小型卵较大型卵更容易遭到巢捕食, 而形状指数对小型卵的被捕食率无显著相关性[29-30]。因此, 小面积的繁殖生境对于蓝冠噪鹛这一产小型卵的鸟类而言, 对于降低天敌捕食压力可能具有重要作用, 而形状指数对天敌影响较小, 因此蓝冠噪鹛对生境斑块的形状选择性不强。此外, 蓝冠噪鹛飞行能力较弱, 繁殖期活动范围较小, 因此斑块面积对其生境选择没有显著影响。
当不同功能的斑块相互隔离时,动物需要在各种生境斑块中进行权衡,以满足其生活需要[31]。有研究表明靠近斑块生境外侧巢捕食风险更高[28], 从蓝冠噪鹛繁殖期区斑块形状指数上看, 多数斑块形状指数接近1, 表示近圆形, 圆形斑块边缘效应较高, 捕食风险的压力较小;但F点形状指数较大, 说明其形状较为复杂。
Grubb和Greenwald早在1985年即指出在哪里营巢是鸟类繁殖期的一个重要行为决策[31], 巢域附近的植被结构特征能影响巢的捕食率[32], 所以鸟类的繁殖栖息地选择, 尤其是巢址的选择, 常常取决于植被条件。
繁殖期蓝冠噪鹛在斑块尺度上对于低海拔区域表现出强烈的选择, 且更偏好距离村庄和山地更近的阔叶林。而对斑块的大小、形状等没有明显的选择性。对水源的选择性没有达到显著性水平, 可能与婺源水系丰富、河流众多有关。婺源县的村庄基本都在低海拔的山谷之间,且基本都沿河而建。蓝冠噪鹛作为亲人鸟类, 喜好选择这样的区域可能与生境条件适合有关。首先是婺源县森林景观类型以阔叶林为主, 其斑块数量和斑块面积均为最大[32], 而蓝冠噪鹛繁殖期喜在阔叶树上筑巢, 而阔叶林在婺源基本分布在村庄及其周边。因此, 从斑块尺度看, 林型可能是影响其生境选择的重要原因。其次, 靠近水边的阔叶林及村庄的菜地、果园等农耕区食物丰富度很高, 可以为蓝冠噪鹛提供丰富的食物来源。而且, 居民区相对山地猛禽等天敌较少, 对于蓝冠噪鹛而言捕食压力较低。且婺源全县相较于江西其他县市景观破碎度较低, 特别是阔叶林斑块连通性较强[34], 因此适宜蓝冠噪鹛栖息繁殖。
4.2 微生境尺度生境选择特征Cody早在1985年即指出在哪里营巢是鸟类繁殖期的一个重要行为决策[33],巢域附近的植被结构特征能影响巢的捕食率[34],所以鸟类的繁殖栖息地选择,尤其是巢址的选择,常常取决于植被条件。蓝冠噪鹛在选择繁殖生境时对村庄附近阔叶林及水源都有明显的选择偏好。阔叶树是其主要的筑巢树, 尤其喜选择树龄较高(树高和胸径较大)的乔木筑巢。高大乔木隐蔽性较高, 有利于其繁殖期躲避捕食压力。在微生境尺度, 树种、草本密度及草本高度是巢区区别于对照区的主要因子。同样说明微生境尺度蓝冠噪鹛对筑巢树种具有明显的选择性。资源选择函数分析结果表明蓝冠噪鹛巢址选择与草本密度及草本高度关系最为密切, Vanderploeg和Scavia选择系数分析结果表明草本盖度较大及草本高度较低是蓝冠噪鹛筑巢偏好的生境。而草本的差异与食物丰富度密切相关。在繁殖期初期, 蓝冠噪鹛喜爱在地面取食, 已有的一些研究表明, 昆虫以及无脊椎动物的丰富度与草本层植物的发育状况相关[35-36]。微生境尺度对草本的选择说明其繁殖期偏好在食物丰富度较高的区域筑巢, 一方面可以获得更好的食物资源, 另一方面可以减少觅食的活动距离。而巢区草本高度较对照区小则是人为干扰造成的结果。繁殖季节, 该研究地聚集很多拍鸟和观鸟爱好者, 其活动路线以追随蓝冠噪鹛为主, 因此造成巢区草本踩踏严重, 故草本高度低于对照区。其他因子没有显著差异与研究地的生境异质性较小有关, 繁殖点B大部分的树几乎都是同时人工栽植的, 因此树高和胸径之间差异不大。
综合以上结果, 斑块尺度蓝冠噪鹛选择在低海拔阔叶林繁殖, 且繁殖区域面积较小, 距离村庄和水源很近, 形状变异较大, 但多为近似方形。微生境尺度蓝冠噪鹛在营巢期的巢址选择时以树种、树高及胸径、草本密度及高度、距干扰源距离为主要影响因子, 说明蓝冠噪鹛偏好在村庄附近高大阔叶树上筑巢, 对树种(朴树、枫杨和枫香)有明显的选择性。巢下草本盖度较好, 反映了繁殖期蓝冠噪鹛对食物、隐蔽条件及天敌防御等方面的需求。
| [1] | Gilliam J F, Fraser D F. Habitat selection under predation hazard:test of a model with foraging Minnows. Ecology, 1987, 68(6): 1856–1862. DOI:10.2307/1939877 |
| [2] | William B, Kristan Ⅲ. Sources and expectations for hierarchical structure in bird-habitat associations. The Condor, 2006, 108(1): 5–12. DOI:10.1650/0010-5422(2006)108[0005:SAEFHS]2.0.CO;2 |
| [3] | 杨勇, 温俊宝, 胡德夫. 鸟类栖息地研究进展. 林业科学, 2011, 47(11): 172–180. DOI:10.11707/j.1001-7488.20111128 |
| [4] | 曹铭昌, 刘高焕, 徐海根. 丹顶鹤多尺度生境选择机制——以黄河三角洲自然保护区为例. 生态学报, 2011, 31(21): 6344–6352. |
| [5] | 李东来, 魏宏伟, 孙兴海, 张正旺. 震旦鸦雀在镶嵌型芦苇收割生境中的巢址选择. 生态学报, 2015, 35(15): 5009–5017. |
| [6] | Greenberg R, Gonzales C E, Bichier P, Reitsma R. Nonbreeding habitat selection and foraging behavior of the Black-Throated Green Warbler complex in southeastern Mexico. The Condor, 2001, 103(1): 31–37. DOI:10.1650/0010-5422(2001)103[0031:NHSAFB]2.0.CO;2 |
| [7] | Kostrzewa A. A comparative study of nest-site occupancy and breeding performance as indicators for nestinghabitat quality in three European raptor species. Ethology Ecology & Evolution, 1996, 8(1): 1–18. |
| [8] | Doucette L I. Home range and territoriality of Australian owlet-nightjars Aegotheles cristatus in diverse habitats. Journal of Ornithology, 2010, 151(3): 673–685. DOI:10.1007/s10336-010-0497-9 |
| [9] | Murray L D, Best L B. Nest-site selection and reproductive success of Common Yellowthroats in managed Iowa grasslands. The Condor, 2014, 116(1): 74–83. DOI:10.1650/CONDOR-13-047-R1.1 |
| [10] | 李枫, 杨红军, 张洪海, 高中信. 扎龙湿地丹顶鹤巢址选择研究. 东北林业大学学报, 1999, 27(6): 57–60. |
| [11] | 程雅畅. 基于GPS遥测的江西鄱阳湖越冬白枕鹤(Grus vipio)活动区和栖息地选择研究[D]. 北京: 北京林业大学, 2015. http://cdmd.cnki.com.cn/Article/CDMD-10022-1015319424.htm |
| [12] | 黄建, Huettmann F, 郭玉民. 黑龙江流域白头鹤繁殖栖息地选择模型预测. 北京林业大学学报, 2015, 37(8): 40–47. |
| [13] | 曾晴. 中华秋沙鸭越冬栖息地利用及越冬种群估计[D]. 北京: 北京林业大学, 2015. http://cdmd.cnki.com.cn/Article/CDMD-10022-1015319503.htm |
| [14] | 徐基良, 张晓辉, 张正旺, 郑光美. 白冠长尾雉育雏期的栖息地选择. 动物学研究, 2002, 23(6): 471–476. |
| [15] | 徐基良, 张晓辉, 张正旺, 郑光美, 阮祥锋, 朱家贵, 溪波. 白冠长尾雉越冬期栖息地选择的多尺度分析. 生态学报, 2006, 26(7): 2061–2067. |
| [16] | Xu J L, Zhang X H, Sun Q H, Zheng G M, Wang Y, Zhang Z W. Home range, daily movements and site fidelity of male Reeves's pheasants Syrmaticus reevesii in the dabie mountains, central China. Wildlife Biology, 2009, 15(3): 338–344. DOI:10.2981/08-032 |
| [17] | 洪元华, 郑磐基, 刘智勇, 何芬奇. 黄腹噪鹛在中国婺源的重新发现. 动物学研究, 2002, 23(5): 383-383, 404-404. |
| [18] | 刘智勇, 洪元华, 姜经宙. 江西省婺源县黄喉噪鹛调查初报. 四川动物, 2001, 20(4): 213–213. |
| [19] | 廖为明, 洪元华, 俞社保, 欧阳勋志, 贺桂先. 婺源黄喉噪鹛繁殖生态及其与村落风水林关系研究. 江西农业大学学报, 2007, 29(5): 837-841, 850-850. |
| [20] | 洪元华, 俞社保, 廖为明. 婺源黄喉噪鹛繁殖生境研究. 江西农业大学学报, 2006, 28(6): 907–911. |
| [21] | Manly B F J, Mcdonald L L, Thomas D L. Resource Selection By Animals:Statistical Design and Analysis for Field Studies. London: Chapman & Hall, 1993. |
| [22] | Lennon J J. Resource selection functions:taking space seriously. Trends in Ecology & Evolution, 1999, 14(10): 399–400. |
| [23] | 韩宗先, 魏辅文, 张泽钧, 李明, 张保卫, 胡锦矗. 蜂桶寨自然保护区小熊猫对生境的选择. 兽类学报, 2004, 24(3): 185–192. |
| [24] | Boyce M S, McDonald L L. Relating populations to habitats using resource selection functions. Trends in Ecology & Evolution, 1999, 14(7): 268–272. |
| [25] | Vanderploeg H A, Scavia D. Calculation and use of selectivity coefficients of feeding:zooplankton grazing. Ecological Modelling, 1979, 7(2): 135–149. DOI:10.1016/0304-3800(79)90004-8 |
| [26] | 徐化成. 景观生态学. 北京: 中国林业出版社, 1996. |
| [27] | 郭晋平, 周志翔. 景观生态学. 北京: 中国林业出版社, 2007. |
| [28] | Pasinelli G, Schiegg K. Fragmentation within and between wetland reserves:the importance of spatial scales for nest predation in reed buntings. Ecography, 2006, 29(5): 721–732. DOI:10.1111/eco.2006.29.issue-5 |
| [29] | 孙吉吉, 王思宇, 王彦平, 邵德钰, 丁平. 千岛湖栖息地片段化效应对鸟类巢捕食风险的影响. 生物多样性, 2011, 19(5): 528–534. |
| [30] | 张蒙, 孙吉吉, 王彦平, 蒋萍萍, 丁平, 徐高福. 千岛湖栖息地片段化对大山雀营巢资源利用的影响. 生物多样性, 2010, 18(4): 383–389. |
| [31] | Grubb Jr T C, Greenwald L. Sparrows and a brushpile:foraging responses to different combinations of predation risk and energy cost. Animal Behaviour, 1982, 30(3): 637–640. DOI:10.1016/S0003-3472(82)80133-4 |
| [32] | 欧阳勋志, 廖为明, 俞社保, 郑世跃, 彭世揆, 孙科辉. 婺源县森林景观空间格局及其与景观美学质量关系探析. 江西农业大学学报, 2005, 27(6): 880–884. |
| [33] | Cody M L. Habitat Selection in Birds. London: Academic Press, 1985. |
| [34] | Martin T E, Roper J J. Nest predation and nest-site selection of a western population of the hermit thrush. The Condor, 1988, 90(1): 51–57. DOI:10.2307/1368432 |
| [35] | Tscharntke T, Greiler H J. Insect communities, grasses, and grasslands. Annual Review of Entomology, 1995, 40(1): 535–558. DOI:10.1146/annurev.en.40.010195.002535 |
| [36] | Morris D L, Thompson Ⅲ F R. Effects of habitat and invertebrate density on abundance and foraging behavior of brown-headed cowbirds. The Auk, 1998, 115(2): 376–385. DOI:10.2307/4089196 |