 2018, Vol. 38
2018, Vol. 38文章信息
- 王颖, 宗宁, 何念鹏, 张晋京, 田静, 李良涛.
- WANG Ying, ZONG Ning, HE Nianpeng, ZHANG Jinjing, TIAN Jing, LI Liangtao.
- 青藏高原高寒草甸不同海拔梯度下土壤微生物群落碳代谢多样性
- Soil microbial functional diversity patterns and drivers along an elevation gradient on Qinghai-Tibet, China
- 生态学报. 2018, 38(16): 5837-5845
- Acta Ecologica Sinica. 2018, 38(16): 5837-5845
- http://dx.doi.org/10.5846/stxb201707261343
-
文章历史
- 收稿日期: 2017-07-26
- 网络出版日期: 2018-05-11





2. 河北工程大学园林与生态工程学院, 邯郸 056001;
3. 吉林农业大学资源与环境学院, 长春 130118
2. College of Landscape and Ecological Engineering, Hebei University of Engineering, Handan 056001, China;
3. College of Resources and Environmental Science, Jilin Agricultural University, Changchun 130118, China
土壤微生物作为土壤中重要的生物组成部分, 是土壤有机质和养分循环的主要驱动力[1-3], 在调控生物地球化学循环过程和维持生态系统功能方面起着关键作用[4-5]。近年来, 土壤微生物群落和功能多样性已经成为生态学领域的研究热点[6], 揭示土壤微生物群落和功能多样性的变化规律以及其影响机制, 可为生态系统服务以及管理提供重要科学依据[7-8]。
海拔梯度的上升会引起气候、植被以及土壤因素的变化, 进而可能引起土壤微生物功能多样性的改变[9-13]。近年来一些研究报道了土壤微生物功能多样性海拔分布格局及影响因子。例如宋贤冲等[14]采用Biolog法研究了广西猫儿山不同海拔植被带土壤微生物碳源利用能力, 研究发现随着海拔上升土壤微生物碳源利用能力呈现线性下降趋势, 其中土壤全钾和土壤含水量是主要影响因子。相反, 通过研究贺兰山不同海拔植被带的土壤微生物碳代谢功能, 发现海拔最高的亚高山草甸土壤微生物群落利用碳源的能力最强, 而处于最低海拔处的荒漠草原的平均颜色变化率(average well color development, AWCD)最低, 并指出地上植被类型对土壤微生物群落碳源利用类型多样性有重要影响[8]。通过分析西藏色季拉山西坡不同海拔下的土壤微生物碳源代谢, 也发现土壤微生物碳源代谢活性随海拔的升高而降低, 其中土壤pH是主要影响因素[6]。借助功能基因芯片, 杨云峰等发现青藏高原土壤微生物功能基因在不同海拔梯度上存在差异[15]。研究发现土壤微生物活性(如参与C、N和P循环的酶)呈现随海拔上升而升高[16-17]或降低[17-18]的趋势。以上研究表明, 微生物功能多样性在海拔梯度变化中会表现不同的变化趋势, 主要影响因子也存在很大不同。这些研究的差异可能和不同地区不同海拔梯度下的地形条件、土壤环境、气候因子以及地上植被状况存在密切关系, 因此有待加强不同地区不同海拔土壤微生物功能多样性变化格局和影响机制的研究。
青藏高原是地球陆地生态系统的重要组成部分, 是生态环境最为奇特、生物资源最为丰富的自然资源宝库之一, 具有独特的海拔、气候和生态系统类型, 不仅对全球气候变化十分敏感, 而且在亚洲气候乃至全球气候变化过程中扮演着重要角色[19-20]。作为青藏高原较为典型的植被类型, 高寒草甸是适应高原隆起和长期低温环境形成的特殊产物, 是典型的高原地带性和山地垂直地带性植被, 是青藏高原高寒草地生态系统的主体[21], 其微生物群落功能多样性与其他区域存在巨大差异, 因而具有极高的科学研究价值并逐渐被人们所关注[19]。据此, 本研究目标在于:(1)运用Biolog微平板技术研究青藏高原高寒草甸土壤微生物群落碳代谢多样性沿海拔梯度变化的规律;(2)阐明环境因子对微生物群落碳代谢多样性的影响机制。在全球气候变暖和高寒草甸趋于退化的大背景下, 期望借助该研究为青藏高原高寒草甸地区的土壤质量监测、植被恢复和可持续生态系统管理建设等提供理论科学依据。
1 研究地区与研究方法 1.1 研究区域概况研究区域位于西藏自治区拉萨市北当雄县草原站北侧山峰的南向山坡(30°30′—30°32′N, 91°03′E), 背靠念青唐古拉山脉, 基面海拔4300 m, 最高海拔可达6000 m以上, 草线位置大致在5210 m, 属于高原亚寒带季风半干旱气候。多年平均气温1.8℃, 最冷月(1月)均温-9.1℃, 最热月(7月)11.0℃, 气温年均差达到20℃。多年年平均降水量479 mm, 其中90%集中在5—9月, 雨热同期, 干湿季明显[22]。海拔4600 m以上的主要土壤类型是高山草甸土, 4600 m以下为草原草甸土。4300—4650 m海拔处的植被类型是以丝颖针(S. capillacea)为主的山地草甸, 4650—5100 m海拔处是以高山嵩草(K. pygmaea)为主的典型草甸, 其他优势种还有垫状点地梅(Androsace tapaete)、澜沧雪灵芝(Arenaria lancangensis)、雪白委陵菜(Potentilla nivea)等[22]。
1.2 土壤样品的采集与处理 1.2.1 土壤样品采集2016年6月, 在位于不同海拔(4300、4400、4650、4800、4950、5100 m)的6个长期围封场地内进行土壤取样。每个长期围封样地内设置4个样方小区作为重复, 每个小区间隔不少于2 m, 共设立24个样方, 每个样方面积为5 m×5 m=25 m2。在设立样地时, 尽量使坡度、坡位等生态因子保持一致, 以增加实验的可比性和科学性。在每个小区内采用“五点混合法”取样(土钻直径4 cm, 取样深度10 cm), 将每个小区采集的土壤样品混合为1个样品。将取出的土壤样品装入自封袋, 然后用4℃冰盒低温保存运往实验室。回到实验室后, 仔细地将肉眼可见的根系、石砾和动植物残体去除, 然后土壤过2 mm筛。将用于土壤微生物群落碳源代谢多样性、土壤硝态氮(Nitrate, NO3-N)、氨态氮(Ammonium, NH4+-N)以及溶解性有机碳(Dissolved organic carbon, DOC)测定的样品4℃冰箱储存, 并在一周内完成所有的样品前处理, 另一部分土样置于庇荫处风干用于土壤理化性质的测定。
1.2.2 土壤理化性质具体测定方法:称取5 g土壤在105℃烘12 h测土壤含水量。pH值用水浸提电位法(水土比为2.5:1)。将过2 mm筛后风干土壤样品用球磨仪磨碎, 称取适量样品, 用元素分析仪(ElementarVario EL Ⅲ)测定土壤有机碳(Soil organic carbon, SOC)和全氮(Total nitrogen, TN)。土壤DOC根据Jones方法[23]测定, 10 g湿土加入40 mL 0.05 mol/L K2SO4溶液, 200 r/min震荡60 min, 2500 r/min离心10 min, 将取得的一部分上清液用日本岛津公司TOC-VCPH型分析仪分析。另一部分上清液中NO3-N和NH4+-N含量用流动分析仪(Futura)测定。
1.2.3 土壤微生物碳源利用多样性测定土壤微生物群落碳代谢多样性采用Biolog方法进行[24-25]:称取10 g新鲜土样加入100 mL 0.85%无菌NaCl溶液的三角瓶中, 封口, 180 r/min, 28℃振荡30 min。静置2 min, 取上清液2 mL加入18 mL 0.85%无菌NaCl溶液中, 如此重复稀释1次, 制得1:1000的提取液。取150 μm提取液接种到生态板的每一个孔中, 最后将接种好的板置于28℃的恒温培养箱中培养, 前72 h每隔12 h在Biolog仪读取590 nm的光密度值, 之后改为每24 h读取1次密度值, 培养时间共为168 h。
土壤微生物的代谢活性用每孔颜色平均变化率(AWCD)来描述, 计算公式如下[26]:

|
式中, C为每个有培养基孔的光密度值, R为对照孔的光密度值, n为碳源的数目, Biolog生态板的C源数目为31[27]。本研究采用培养72 h光密度值分析土壤微生物群落功能多样性, 计算公式如下[28]:

|
式中, Pi为第i个孔的相对吸光值与整个微平板相对吸光值的比值, 计算公式为:
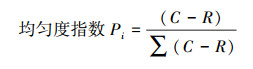
|
通过PCA(Principle component analysis, PCA)对不同海拔微生物群落碳源代谢多样性进行分析。利用VPA(Variation partitioning analysis, VPA)和偏曼特尔检验(Partial mantel test)分析评估不同环境因子对不同海拔微生物碳源代谢多样性的影响大小。PCA、VPA和偏曼特尔检验主要采用R软件的vegan程序包。
2 结果与分析 2.1 不同海拔土壤微生物碳源代谢活性及功能多样性指数变化AWCD反映了微生物群落碳源的利用强度。由图 1可知, 六种不同海拔梯度土壤微生物AWCD值均随时间的延长而升高。不同海拔下AWCD大小顺序为:4800 m>4950 m>4400 m>4650 m>5100 m>4300 m, 表明4800—4950 m海拔地区土壤微生物碳源代谢强度最高。随着海拔上升, 除均匀度指数变化不太明显外, Shannon多样性指数和碳源利用丰富度指数差异显著, 其中在4800—4950 m海拔的三类多样性指数明显高于其他海拔梯度(表 1)。

|
| 图 1 不同海拔土壤微生物群落单孔平均颜色变化率 Fig. 1 Variation in AWCD of soil microbial community over time at different elevations |
| 海拔/m Elevation |
Shannon指数 Shannon index |
均匀度指数 Evenness |
丰富度指数 Richness |
| 4300 | 3.00±0.183c | 1.88±0.076b | 5.00±0.408d |
| 4400 | 4.36±0.065b | 1.79±0.032b | 11.50±0.289bc |
| 4650 | 4.43±0.050b | 1.75±0.022b | 12.50±0.289b |
| 4800 | 6.57±0.215a | 2.33±0.069a | 16.75±0.479a |
| 4950 | 6.17±0.120a | 2.24±0.026a | 15.75±1.031a |
| 5100 | 4.43±0.069b | 1.87±0.020b | 10.75±0.629c |
| 同行不同字母表示差异显著(P<0.05) | |||
通过对不同海拔下土壤微生物对6种主要碳源类型的利用程度分析可以看出(图 2), 不同海拔下土壤微生物对碳源的利用主要集中碳水化合物类、氨基酸类和胺类三大类物质, 对聚合物类和酚酸类物质的利用程度相对较低。不同海拔土壤微生物群落对同一类型碳源的利用强度也有明显差异(图 2)。除了聚合物类, 随着海拔升高, 微生物群落对其他五种类型碳源利用均呈现先上升而后下降的单峰变化趋势, 表现为4800—4950 m碳源利用率达到最高, 表明不同海拔土壤微生物对不同碳源的利用呈现一致的规律, 即4800—4950 m海拔地区土壤微生物群落拥有高的碳源利用能力, 特别是对碳水化合物类、羧酸类、氨基酸类和胺类碳源的利用强度受海拔影响较大。

|
| 图 2 不同海拔下土壤微生物6类碳源利用强度 Fig. 2 Utilization of 6 types of carbon sources of soil microbial community 柱形上方不同小写字母表示不同海拔之间吸光值差异显著(P<0.05) |
PCA分析表明不同海拔土壤微生物群落在碳源利用上有明显的分异, 可见海拔变化对土壤微生物群落碳代谢特征有显著影响(图 3)。其中主成分1(PC1)和主成分2(PC2)的方差贡献率分别为47.6%和19.7%, 累计方差贡献率为67.3%。

|
| 图 3 不同海拔土壤微生物群落代谢主成分分析 Fig. 3 Principle component analysis(PCA)of carbon utilization of soil microbial community at different elevations |
初始载荷因子反映主成分与碳源利用的相关系数, 载荷因子越高表示碳源对主成分的影响越大。从表 2可以看出, 与PC1有较高相关性的碳源有9种, 其中碳水化合物类5种、氨基酸类1种、羧酸类1种、聚合物类1种和酚酸类1种;与PC2有较高相关性的碳源仅有2种, 分别是羧酸类1种和氨基酸类1种。综合以上发现我们也发现, 对土壤微生物群落代谢特征起分异作用的主要碳源类型是碳水化合物类、羧酸类和氨基酸类, 即微生物碳源代谢多样性的垂直地带性差异主要体现在碳水化合物类、氨基酸类和羧酸类, 其中碳水化合物类尤为突出。
| 碳源类型 Carbon source |
主成分1 PC1 |
主成分2 PC2 |
|
| β-甲基-D-葡萄糖苷 | 0.9156 | 0.1558 | |
| D-木糖/戊醛糖 | -0.3508 | 0.0528 | |
| i-赤藓糖醇 | 0.7901 | 0.3944 | |
| D-甘露醇 | 0.8715 | -0.2971 | |
| N-乙酰-D葡萄糖氨 | 0.9338 | 0.1381 | |
| 碳水化合物 | D-纤维二糖 | 0.7462 | -0.5537 |
| Carbohydrate | α-D-乳糖 | 0.3916 | -0.677 |
| D-半乳糖酸γ-内酯 | 0.468 | 0.6184 | |
| D-木糖/戊醛糖 | 0.6542 | -0.1256 | |
| 1-磷酸葡萄糖 | 0.804 | -0.4915 | |
| D, L-α-磷酸甘油 | 0.6854 | 0.0671 | |
| 酚酸类 | 2-羟基苯甲酸 | 0.3872 | -0.5207 |
| Phenolic compounds | 4-羟基苯甲酸 | 0.6634 | 0.6102 |
| 丙酮酸甲酯 | 0.4518 | 0.0472 | |
| γ-羟丁酸 | 0.799 | -0.4353 | |
| 衣康酸 | 0.4588 | 0.7753 | |
| 羧酸类 | α-丁酮酸 | -0.4955 | 0.286 |
| Carboxylic acids | D-苹果酸 | 0.1473 | 0.2734 |
| D-葡糖胺酸 | 0.5077 | 0.541 | |
| L-精氨酸 | 0.4625 | 0.7161 | |
| L-天门冬酰胺 | 0.6877 | 0.509 | |
| 氨基酸类 | L-苯丙氨酸 | 0.7041 | -0.3318 |
| Amino acids | L-丝氨酸 | 0.7958 | 0.4564 |
| L-苏氨酸 | 0.1113 | -0.1432 | |
| 甘氨酰-L-谷氨酸 | 0.4881 | 0.5048 | |
| 胺类 | 苯乙胺 | 0.7302 | 0.5711 |
| Amines | 腐胺 | 0.4448 | 0.1263 |
| 吐温40 | 0.6061 | -0.1625 | |
| 聚合物类 | 吐温80 | 0.7718 | -0.3658 |
| Polymer | α-环式糊精 | 0.663 | -0.0959 |
| 肝糖 | 0.4211 | -0.1665 |
将与不同海拔土壤微生物碳源利用相关的12个环境因子归为土壤、气候和植物三组进行VPA分析(图 4), 其中土壤因子包括土壤含水量、pH、DOC、SOC、TN、NH4+-N、NO3-N, 气候因子包括MAT(Mean annual tempreature, MAT)和MAP(Mean annual precipitation, MAP), 植物因子包括植被Shannon、植被丰富度、植被盖度。发现三组影响因子单独作用以及他们之间的交互作用总共解释了79%的不同海拔的碳源代谢多样性变异, 其中土壤解释了31.7%, 植物和气候分别解释了9.5%和11.5%。表明土壤理化性质是导致不同海拔碳源利用变异的重要原因。进一步的显著性分析表明, 土壤和气候与微生物碳源利用功能呈现极显著相关性(P=0.002、0.003), 植物呈显著相关性(P=0.046)。

|
| 图 4 不同海拔梯度下碳源利用受环境因子影响的分类变异分析 Fig. 4 Variation partitioning analysis(VPA) of carbon source utilization and environmental factors at different elevations 括号里的值表示显著性, *, P<0.05;**, P<0.01 |
为了排除各种环境因子的多重作用及其之间的交互作用, 更好的明确影响土壤微生物碳源利用的主要环境因子, 将与之相关的12个环境因子进行偏曼特尔检验(表 3)。结果表明, 土壤含水量(r=0.043, P=0.001)、植被丰富度(r=0.360, P=0.001)和MAP(r=0.272, P=0.004)是影响土壤微生物碳源利用的最主要的环境因子(P<0.05)。
| 环境因子 Enviromental characteristics |
相关系数(r) Correlation coefficient |
显著性(P) Significance |
| 年平均气温Mean annual temperature | -0.081 | 0.817 |
| 年平均降水Mean annual precipitation | 0.272 | 0.004 |
| 植被多样性Vegetation Shannon-Wiener | 0.132 | 0.066 |
| 植被盖度Vegetation coverage | 0.029 | 0.383 |
| 植被丰富度Vegetation richness | 0.360 | 0.001 |
| 土壤有机碳Soil organic carbon | 0.031 | 0.348 |
| 全氮Total nitrogen | -0.164 | 0.983 |
| 硝态氮Nitrate nitrogen | -0.004 | 0.472 |
| 铵态氮Ammonium nitrogen | 0.283 | 0.017 |
| pH值pH Value | 0.160 | 0.045 |
| 溶解性有机碳Dissolved organic carbon | 0.132 | 0.112 |
| 土壤含水量Soil moisture content | 0.403 | 0.001 |
| 根据不同海拔碳源利用率和环境因子之间进行9999次排列得出相关系数和显著性值 | ||
随着海拔高度的增加, 不同海拔梯度下土壤微生物AWCD值均随时间的延长而上升, 其中4800 m海拔地区的AWCD值显著高于其他海拔梯度, 表明该海拔地区土壤微生物碳源利用强度最高(图 1)。宋贤冲等[14]对广西猫儿山不同海拔植被带土壤微生物单一碳源利用能力的研究表明, 随着海拔上升土壤微生物碳源利用能力呈现下降趋势。刘秉儒等[8]研究表明贺兰山不同海拔植被带的土壤微生物碳代谢功能随海拔的升高而增大。本研究结果表明, 随着海拔升高, 土壤微生物碳源利用能力呈现先上升后下降的单峰变化趋势, 与上述研究结果不一致。一方面, 气候与水文等各种自然地理因素在垂直地带上具有复杂的梯度变化, 许多环境因素和生物特性会随海拔变化而发生改变[17]。除此之外, 低温高湿的气候条件而又极少受到人为干扰也可能是4800 m海拔地区土壤微生物碳源利用最高的重要原因。同时研究还发现微生物碳源利用的垂直地带性差异主要体现在碳水化合物、羧酸、氨基酸和胺类四类碳源的利用上(图 2, 图 3), 这与朱平等[29]的研究结果一致。这可能是由于4800 m海拔地区低温高湿的气候条件限制了土壤中好气性微生物的活动, 不利于难氧化有机碳的矿化, 因而微生物对聚合物类和酚酸类等物质的利用程度相对较低。
3.2 不同海拔对土壤微生物碳源利用多样性影响以及影响机制本研究发现, 海拔显著影响了土壤微生物碳源利用, 同时4800 m海拔处土壤微生物Shannon多样性和碳源利用丰富度指数显著高于其他海拔梯度(表 1, 图 3), 表明该海拔地区土壤微生物碳源利用功能多样性最高。以往大量研究发现, 植物和动物在垂直地带上的多样性分布存在明显的中峰优势[30-32], 即山地生态学中的“中部膨胀”理论[33-34]。本研究结果说明青藏高原高寒草甸草原生态系统的土壤微生物碳源利用多样性存在着与动植物多样性类似的中峰优势分布规律。斯贵才等[30]对藏东南林芝地区色季拉山森林土壤微生物群落结构与土壤酶活性沿海拔梯度变化的研究中发现, 森林土壤微生物量在3900 m海拔地区达到最高, 是第一次研究表明森林生态系统土壤微生物量在海拔梯度上呈现出与动植物类似的中峰优势。于建龙等[35]在对青海玉树不同海拔高度草毡土微生物数量及影响因子的研究中也发现, 随着海拔高度的增加, 土壤微生物含量呈现先上升后下降的变化趋势, 同样表现为明显的中峰优势。但也有研究报道青藏高原色季拉山西坡3105—4556 m土壤微生物群落组成和结构的研究中没有表现出明显的海拔分布特征[36]。
不同海拔土壤微生物生存环境的不同是导致土壤微生物功能多样性分布存在明显梯度差异的主要原因[37]。该研究中微生物功能多样性呈现的中峰优势表明青藏高原高寒草甸土壤微生物的生存环境在中间海拔4800 m处达到最佳, 在此基础上海拔的升高或降低, 都会造成土壤微生物环境的改变进而影响微生物的碳源多样性。研究发现该地区呈现的中峰优势是由多种环境因子共同作用的结果, 其中包括土壤、植物和气候的多重作用且各种环境因子之间存在交互影响(图 4)。VPA分析表明, 土壤、植物和气候因子的多重及交互作用共解释了微生物碳源利用多样性在海拔梯度上变异的79%, 其中土壤因子对土壤微生物碳源代谢的贡献率最大(图 4)。研究发现, 气候和植被可通过改变土壤状况来影响土壤养分进而对微生物碳源代谢产生影响[37], 比如, 气候因子(MAT和MAP)中的MAP可能通过增加土壤湿度直接影响微生物活性[16, 38], 植物群落组成和结构则可能通过影响根际土壤的理化性质进而影响微生物功能多样性[16, 39]。所以, 在各种环境因子多重作用及其交互作用的复杂关系中, 土壤要素能对微生物碳源利用产生更大的影响。
为了排除各种环境因子的多重作用及其之间的交互作用, 偏曼特尔检验表明土壤含水量、植被丰富度和MAP是影响土壤微生物碳源代谢最重要的环境因子(表 3)。土壤含水量是影响土壤微生物代谢活性及功能多样性的重要因素, 而降水增加土壤湿度, 同时降水的变化还能引起土壤中速效磷含量的变化[40-41], 说明降水量的变化能够明显改善土壤微生物生存的营养环境, 从而对土壤微生物活性产生影响[41-42], 所以土壤含水量和MAP可以共同影响土壤微生物碳源利用。在青藏高原的高寒草甸生态系统中, 4800 m海拔处的降水最为丰富, 土壤含水量最高, 温度也相对较低, 土壤微环境有明显的低温高湿特征, 同时土壤蒸发量也相应减少, 所以土壤中可以保持较多的水分[43], 进而可能引起土壤含水量和MAP与微生物碳源利用呈现极显著相关(P=0.001, 0.004)。相关研究显示, 土壤含水量在调节微生物活性与多样性方面至关重要, 它直接影响微生物的生理状态, 限制微生物分解某些化合物的能力, 同时调节土壤酶和土壤理化性质, 进而影响微生物组成与活性[44]。Cederlunda等[45]的研究表明, 含水量高的土壤有较高微生物活性, 王杰等[38]对贝加尔针茅草原土壤微生物碳源利用多样性对氮素和水分添加的响应的研究表明水分有利于氮肥肥效的发挥, 能提高土壤微生物的活性, 是影响微生物碳源利用多样性的重要因素。文东新等[39]对衡阳紫色土丘陵坡地植被恢复对土壤微生物碳源代谢多样性的影响的研究发现土壤含水量的增加有利于群落结构优化并提高群落地上生物量, 另外土壤含水量还可以通过影响土壤总碳、TN等理化性质, 促进土壤微生物大量活动, 加快土壤C元素的循环过程和土壤的矿化过程, 从而提高土壤微生物活性与功能多样性。植被丰富度也是影响土壤微生物碳源利用多样性的主要影响因子(表 3)。这和以往研究结果类似, 如冯泉清[46]等研究发现地上植被类型对土壤微生物碳源利用类型多样性的影响较大。一方面是由于植物群落的组成和结构可能会改变植物根际土壤微生物的多样性, 从而使微生物群落功能多样性产生相应的变化[47], 另一方面, 地上的植物多样性可能会通过凋落物的差异对土壤微生物碳源代谢产生重要影响[7, 12]。
4 结论青藏高原高寒草甸土壤微生物碳源代谢多样性具有明显的垂直地带性差异, 土壤微生物对碳源的利用在中间海拔4800 m处取得最高。土壤微生物对不同类型碳源利用强度存在一定的差异, 微生物功能多样性的垂直地带性差异主要体现在对碳水化合物类、氨基酸类和羧酸类碳源的利用上。土壤微生物碳源代谢活性和多样性随海拔上升均呈现先上升后下降的单峰变化趋势。该研究发现, 不同海拔土壤微生物碳源利用是土壤、植物和气候等多重环境因子综合作用结果, 其中土壤含水量、植被丰富度和MAP是导致不同海拔土壤微生物碳源利用差异的关键环境因子。
| [1] | Li F L, Liu M, Li Z P, Jiang C Y, Han F X, Che Y P. Changes in soil microbial biomass and functional diversity with a nitrogen gradient in soil columns. Applied Soil Ecology, 2013, 64: 1–6. DOI:10.1016/j.apsoil.2012.10.006 |
| [2] | Gryta A, Frąc M, Oszust K. The application of the BiologEcoPlate approach in ecotoxicological evaluation of dairy sewage sludge. Applied Biochemistry and Biotechnology, 2014, 174(4): 1434–1443. DOI:10.1007/s12010-014-1131-8 |
| [3] | Djukic I, Zehetner F, Mentler A, Gerzabek M H. Microbial community composition and activity in different Alpine vegetation zones. Soil Biology and Biochemistry, 2010, 42(2): 155–161. DOI:10.1016/j.soilbio.2009.10.006 |
| [4] | Bell T, Newman J A, Silverman B W, Turner S L, Lilley A K. The contribution of species richness and composition to bacterial services. Nature, 2005, 436(7054): 1157–1160. DOI:10.1038/nature03891 |
| [5] | Zhang X, Zhao X, Zhang M. Functional diversity changes of microbial communities along a soil aquifer for reclaimed water recharge. FEMS Microbiology Ecology, 2012, 80(1): 9–18. DOI:10.1111/j.1574-6941.2011.01263.x |
| [6] | 徐梦. 海拔梯度及土地利用方式对藏东南地区土壤有机碳和微生物群落的影响[D]. 北京: 中国农业大学, 2016. http://cdmd.cnki.com.cn/Article/CDMD-10019-1016085268.htm |
| [7] | 傅伯杰, 张立伟. 土地利用变化与生态系统服务:概念、方法与进展. 地理科学进展, 2014, 33(4): 441–446. |
| [8] | 刘秉儒, 张秀珍, 胡天华, 李文金. 贺兰山不同海拔典型植被带土壤微生物多样性. 生态学报, 2013, 33(22): 7211–7220. |
| [9] | Gaston K J. Global patterns in biodiversity. Nature, 2000, 405(6783): 220–227. DOI:10.1038/35012228 |
| [10] | Stevens G C. The elevational gradient in altitudinal range:an extension of Rapoport's latitudinal rule to altitude. The American Naturalist, 1992, 140(6): 893–911. DOI:10.1086/285447 |
| [11] | Lomolino MV. Elevation gradients of species-density:historical and prospective views. Global Ecology and Biogeography, 2001, 10(1): 3–13. DOI:10.1046/j.1466-822x.2001.00229.x |
| [12] | 吴则焰, 林文雄, 陈志芳, 刘金福, 方长旬, 张志兴, 吴林坤, 陈婷. 武夷山不同海拔植被带土壤微生物PLFA分析. 林业科学, 2014, 50(7): 105–112. |
| [13] | 贺金生, 陈伟烈. 陆地植物群落物种多样性的梯度变化特征. 生态学报, 1997, 17(1): 91–99. DOI:10.3321/j.issn:1000-0933.1997.01.014 |
| [14] | 宋贤冲, 陈晓鸣, 郭丽梅, 邓小军, 赵连生, 曹继钊. 猫儿山不同海拔典型植被带土壤酶活性变化特征. 基因组学与应用生物学, 2016, 35(12): 3545–3551. |
| [15] | Yang Y F, Gao Y, Wang S P, XuD P, Yu H, Wu L W, Lin Q Y, Hu Y G, Li X Z, He Z L, Deng Y, Zhou J Z. The microbial gene diversity along an elevation gradient of the Tibetan grassland. The ISME Journal, 2014, 8(2): 430–440. DOI:10.1038/ismej.2013.146 |
| [16] | 斯贵才, 袁艳丽, 王建, 夏燕青, 雷天柱, 张更新. 藏东南森林土壤微生物群落结构与土壤酶活性随海拔梯度的变化. 微生物学通报, 2014, 41(10): 2001–2011. |
| [17] | 谷晓楠, 贺红士, 陶岩, 靳英华, 张心昱, 徐志伟, 王钰婷, 宋祥霞. 长白山土壤微生物群落结构及酶活性随海拔的分布特征与影响因子. 生态学报, 2017, 37(24): 8374–8384. |
| [18] | Xu ZW, Yu GR, Zhang X Y, Ge JP, He NP, WangQF, WangD. The variations in soil microbial communities, enzyme activities and their relationships with soil organic matter decomposition along the northern slope of Changbai Mountain. Applied Soil Ecology, 2015, 86: 19–29. DOI:10.1016/j.apsoil.2014.09.015 |
| [19] | 周宁一. 青藏高原微生物多样性研究. 微生物学通报, 2014, 41(11): 2378–2378. |
| [20] | 余莲, 封彩云. 青藏高原近期气候变化研究进展. 高原山地气象研究, 2012, 32(3): 84–88. DOI:10.3969/j.issn.1674-2184.2012.03.018 |
| [21] | 牛犇, 张立峰, 马荣荣, 刘晓琴, 张翔, 赵亮, 古松. 高寒草甸土壤微生物量及酶活性的研究. 南开大学学报:自然科学版, 2016, 49(4): 53–60. |
| [22] | Li R C, Luo T X, Mölg T, Zhao J X, Li X, Cui X Y, Du M Y, Tang Y H. Leaf unfolding of Tibetan alpine meadows captures the arrival of monsoon rainfall. Scientific Reports, 2016, 6: 20985. DOI:10.1038/srep20985 |
| [23] | Jones D L, Willett V B. Experimental evaluation of methods to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil. Soil Biology and Biochemistry, 2006, 38(5): 991–999. DOI:10.1016/j.soilbio.2005.08.012 |
| [24] | Garland J L, MillsA L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Applied and Environmental Microbiology, 1991, 57(8): 2351–2359. |
| [25] | Zheng HL, Weiner L M, Bar-Am O, Epsztejn S, Cabantchik Z I, Warshawsky A, YoudimM B H, Fridkin M. Design, synthesis, and evaluation of novel bifunctional iron-chelators as potential agents for neuroprotection in Alzheimer's, Parkinson's, and other neurodegenerative diseases. Bioorganic & Medicinal Chemistry, 2005, 13(3): 773–783. |
| [26] | Jin Z, Ji F Y, Xu X, Xu X Y, Chen Q K, Li Q. Microbial and metabolic characterization of a denitrifying phosphorus-uptake/side stream phosphorus removal system for treating domestic sewage. Biodegradation, 2014, 25(6): 777–786. DOI:10.1007/s10532-014-9698-x |
| [27] | Garland J L. Analytical approaches to the characterization of samples of microbial communities using patterns of potential C source utilization. Soil Biology and Biochemistry, 1996, 28(2): 213–221. DOI:10.1016/0038-0717(95)00112-3 |
| [28] | 马克平. 生物群落多样性的测度方法——Ⅰα多样性的测度方法(上). 生物多样性, 1994, 2(3): 162–168. |
| [29] | 朱平, 陈仁升, 宋耀选, 刘光琇, 陈拓, 张威. 祁连山不同植被类型土壤微生物群落多样性差异. 草业学报, 2015, 24(6): 75–84. |
| [30] | Heaney L R. Small mammal diversity along elevational gradients in the Philippines:an assessment of patterns and hypotheses. Global Ecology and Biogeography, 2001, 10(1): 15–39. DOI:10.1046/j.1466-822x.2001.00227.x |
| [31] | Goodman S M, Andrianarimisa A, Olson L E, Soarimalala V. Patterns of elevational distribution of birds and small mammals in the humid forests of Montagne d'Ambre, Madagascar. Ecotropica, 1996, 2: 87–98. |
| [32] | Rickart E A. Elevational diversity gradients, biogeography and the structure of montane mammal communities in the intermountain region of North America. Global Ecology and Biogeography, 2001, 10(1): 77–100. DOI:10.1046/j.1466-822x.2001.00223.x |
| [33] | Colwell R K, Lees D C. The mid-domain effect:geometric constraints on the geography of species richness. Trends in Ecology & Evolution, 2000, 15(2): 70–76. |
| [34] | 王襄平, 方精云, 唐志尧. 中域效应假说:模型、证据和局限性. 生物多样性, 2009, 17(6): 568–578. |
| [35] | 于健龙, 石红霄. 青海玉树不同海拔高度草毡土微生物数量及影响因子. 中国草地学报, 2011, 33(6): 46–50. |
| [36] | 杨钙仁, 童成立, 张文菊, 吴金水. 陆地碳循环中的微生物分解作用及其影响因素. 土壤通报, 2005, 36(4): 605–609. DOI:10.3321/j.issn:0564-3945.2005.04.033 |
| [37] | 曹成有, 邵建飞, 蒋德明, 崔振波. 围栏封育对重度退化草地土壤养分和生物活性的影响. 东北大学学报:自然科学版, 2011, 32(3): 427-430, 451-451. DOI:10.3969/j.issn.1005-3026.2011.03.031 |
| [38] | 王杰, 李刚, 修伟明, 宋晓龙, 赵建宁, 杨殿林. 氮素和水分对贝加尔针茅草原土壤酶活性和微生物量碳氮的影响. 农业资源与环境学报, 2014, 31(3): 237–245. |
| [39] | 文东新, 杨宁, 杨满元. 衡阳紫色土丘陵坡地植被恢复对土壤微生物功能多样性的影响. 应用生态学报, 2016, 27(8): 2645–2654. |
| [40] | 王楠楠, 杨雪, 李世兰, 隋心, 韩士杰, 冯富娟. 降水变化驱动下红松阔叶林土壤真菌多样性的分布格局. 应用生态学报, 2013, 24(7): 1985–1990. |
| [41] | 王宁, 王美菊, 李世兰, 王楠楠, 冯富娟, 韩士杰. 降水变化对红松阔叶林土壤微生物生物量生长季动态的影响. 应用生态学报, 2015, 26(5): 1297–1305. |
| [42] | 孙雪, 韩冬雪, 刘岩, 冯富娟, 隋心. 原始红松林土壤理化及微生物碳代谢特征对生长季动态的响应. 南京林业大学学报:自然科学版, 2017, 41(5): 18–26. |
| [43] | 马和平, 郭其强, 刘合满, 钱登锋. 西藏色季拉山土壤微生物量碳和易氧化态碳沿海拔梯度的变化. 水土保持学报, 2012, 26(4): 163-166, 171-171. |
| [44] | Sardans J, Peñuelas J, Estiarte M. Changes in soil enzymes related to C and N cycle and in soil C and N content under prolonged warming and drought in a Mediterranean shrubland. Applied Soil Ecology, 2008, 39(2): 223–235. DOI:10.1016/j.apsoil.2007.12.011 |
| [45] | Cederlund H, Thierfelder T, Stenström J. Functional microbial diversity of the railway track bed. Science of the Total Environment, 2008, 397(1/3): 205–214. |
| [46] | 冯泉清, 伍建榕, 张俊忠, 韩庆莉, 刘丽, 马焕成. 高黎贡山旱冬瓜林下土壤微生物碳源代谢多样性分析. 西部林业科学, 2015, 44(4): 47–52. |
| [47] | Choi K H, Dobbs F C. Comparison of two kinds ofBiologmicroplates (GN and ECO) in their ability to distinguish among aquatic microbial communities. Journal of Microbiological Methods, 1999, 36(3): 203–213. DOI:10.1016/S0167-7012(99)00034-2 |