 2018, Vol. 38
2018, Vol. 38文章信息
- 张静, 温仲明, 李鸣雷, 朱朵菊, 陶宇, 曾鸿文.
- ZHANG Jing, WEN Zhongming, LI Minglei, ZHU Duoju, TAO Yu, ZENG Hongwen.
- 外来物种刺槐对土壤微生物功能多样性的影响
- Effects of the exotic black locust on the functional diversity of soil microorganisms
- 生态学报. 2018, 38(14): 4964-4974
- Acta Ecologica Sinica. 2018, 38(14): 4964-4974
- http://dx.doi.org/10.5846/stxb201706171102
-
文章历史
- 收稿日期: 2017-06-17
- 网络出版日期: 2018-04-04





2. 中国科学院教育部水土保持与生态环境研究中心, 杨凌 712100;
3. 西北农林科技大学水土保持研究所, 杨凌 712100;
4. 西北农林科技大学林学院, 杨凌 712100
2. Research Center of Soil and Water Conservation and Ecological Environment, Chinese Academy of Sciences and Ministry of Education, Yangling 712100, China;
3. Institute of Soil and Water Conservation, Northwest A & F University, Yangling 712100, China;
4. College of Forestry, Northwest A & F University, Yangling 712100, China
土壤微生物是生态系统的重要组成部分, 是陆地生态系统养分转化和循环的主要驱动者, 影响土壤有机质的分解、腐化和矿化等过程[1], 对养分循环与平衡、土壤改良发挥重要作用。土壤微生物群落组成不仅与地上植物群落组成结构有关, 也深受温度、降雨等环境条件的影响, 并且通过对系统生物地球化学循环过程的影响, 与地上植物群落形成相互作用的反馈体系[2], 对生态系统过程与功能产生重要影响。通常条件下, 随着环境梯度及植物群落结构的变化, 微生物群落的结构与功能都会发生相应的变化。
外来物种的引入往往也会引起群落物种结构的变化, 进而引起土壤微生物结构与功能的变化。外来物种可通过影响养分利用率及碳储量减少来促进氮循环, 增加土壤氮库及氮通量[3], 从而影响土壤微生物多样性与活性[4-5]以及植物固氮和释放氮的数量和质量[6]。大量研究表明, 外来物种由于其特殊的性状特征, 会对本地生态系统产生重要影响, 如Lee等[6]研究发现外来物种特殊的叶性状和枯落物性状会影响土壤氮循环过程, 影响本地生态系统土壤微生物的活性, 最终影响本地生态系统的功能;Hawkes等[5]研究发现外来物种会改变土壤微生物群落结构, 对生态系统过程产生影响。尤其是一些固氮物种[7], 其特殊的性状使他们比乡土树种有更强的氮利用策略[8-9], 影响氮循环的速率, 减少土壤养分保持能力和土壤碳储量, 增加温室气体的排放[10], 并最终影响到整个系统的结构与功能。
刺槐(Robinia pseudoacacia)作为固氮植物, 因其具有特殊的根瘤结构及快速生长等特点, 在世界各地广泛引种栽植。在我国黄土高原地区也有大面积种植, 是黄土高原人工造林的主要树种之一。作为生态系统的重要组成部分, 刺槐引种对土壤微生物群落的影响自然也引起众多学者关注, 并开展了较多的研究, 如胡婵娟等[11]比较刺槐林与沙棘(Hippophae rhamnoides)、杏树(Armeniaca vulgaris)人工林下土壤微生物, 发现刺槐林下微生物量碳、微生物量氮均高于其他两种林地, 翟辉等[12]比较了刺槐林与柠条(Caragana korshinskii)、油松(Pinus tabulaeformis)等其他植被类型下土壤微生物, 发现除柠条林, 刺槐林下土壤微生物活性高于其他植被类型;但张海涵等[13]将刺槐与柠条、沙棘等5种造林树种根际土壤微生物进行研究, 发现刺槐林下土壤微生物活性和微生物功能多样性均低于其他造林树种, 曾全超等[14]比较刺槐林与辽东栎(Quercus wutaishanica)、油松等天然次生林土壤微生物, 发现刺槐林土壤微生物氮库低于其他天然次生林。从上述研究可见, 刺槐对土壤微生物群落结构或功能的影响在认识上存在很大差异, 产生这样差异的原因, 可能是这些研究多局限于特定区域, 没有进行较大环境梯度下的比较研究。同时, 大量研究表明, 土壤微生物群落结构与功能不仅取决于地上植物群落的物种组成, 也取决于温度、降雨等环境条件的变化[15]。与乡土植物植物群落比较, 刺槐对土壤微生物功能多样性的影响在不同的环境梯度下可能不同, 但目前尚未有系统的比较研究。鉴于此, 本研究以黄土丘陵区不同植被区(草原区、森林草原区和森林区)为研究区, 利用Biolog ECO微孔板的方法对不同植被区内的人工刺槐群落和乡土植物群落土壤微生物群落功能多样性进行研究, 以期揭示不同环境梯度下人工刺槐林分土壤微生物群落功能多样性的变化规律, 并通过比较各植被区内人工刺槐群落与乡土植物群落的土壤微生物功能多样性, 评估外来物种刺槐对土壤微生物功能多样性影响的表现规律, 为在大尺度上了解人工引种刺槐林对土壤生态过程与功能的影响提供依据。
1 研究地区与方法 1.1 研究区概况选择延河流域为研究区(36.23°—37.17°N, 108.45°—110.28°E), 流域属黄土高原典型丘陵沟壑区, 为暖温带半湿润向半干旱过渡区。气候属大陆性季风气候, 年平均气温8.8 ℃, 年平均降水量505.3 mm, 无霜期157 d, 流域内海拔1142—1411 m。区内土壤为黄绵土, 质地均一。从东南到西北, 随降雨量减少, 温度降低, 植被分布呈现明显的地带性规律, 依次为森林区、森林草原区和草原区。森林区内主要分布辽东栎、侧柏(Platycladus orientalis)、山杨(Populus davidiana)等大中型乔木及黄刺玫(Rosa xanthine)、虎榛子(Ostryopsis davidiana)等中小型灌木;森林草原带在坡面上无自然乔木林发育, 仅有人工灌丛植被在沟谷和峁顶发育, 植被主要为虎榛子、狼牙刺(Sophora viciifolia)、柠条等灌木及草本以白羊草(Bothriochloa ischaemum)、铁杆蒿(Artemisia sacrorum)为主;典型草原带以铁杆蒿、长芒草(Stipa bungeana)、大针茅(Stipa grandis)等草本植物群落为主。该区也是黄土高原植被恢复重建的核心区, 在不同的植被区都分布有大量的人工刺槐林分, 为了解刺槐对土壤微生物结构与功能的影响提供了基础。
1.2 样地设置与样品采集根据水热环境梯度变化和植被类型的地带性差异, 在每个植被区内选择人工刺槐林群落和乡土植物群落(草原区、森林草原区和森林区)作为研究对象, 在每个植被区内至少选择3个立地环境尽量相近(坡位、坡向等)的20 m×20 m的刺槐林群落样地及与其一一对应的乡土植物群落样地, 在各样地内设置相应大小的植物群落样方(表 1)。在样地内进行土壤样品提取:去除表面植物残体和石头, 呈“S”形选取五个同一深度的土壤样品, 混合均匀为一个样品。分别选取0—10、10—20 cm两层土壤样品, 过2 mm筛, 装入无菌样品袋, 用冰盒冷藏带回实验室。一部分风干后用于理化性质的测定, 另一部分储存在4℃冰箱, 用于土壤微生物群落特征测定。
| 样点 Sites |
样地大小 Size/m |
经度 Longitude |
纬度 Latitude |
海拔 Altitude/m |
坡位 Position |
主要林下植被 Main vegetation |
| 草原区刺槐 | 10×10 | 109°03′E | 37°04′N | 1318 | 中坡 | 赖草、旱生芦苇 |
| Steppe locust(SL) | 10×10 | 109°04′E | 37°06′N | 1317 | 峁顶 | 赖草、铁杆蒿、丛生隐子草 |
| 10×10 | 109°03′E | 37°09′N | 1321 | 峁顶 | 赖草、茭蒿、蒙古蒿 | |
| 10×10 | 109°03′E | 37°09′N | 1325 | 中坡 | 赖草、蒙古蒿 | |
| 10×10 | 109°20′E | 36°51′N | 1213 | 上坡 | 铁杆蒿 | |
| 草原区乡土植物 | 2×2 | 109°03′E | 37°05′N | 1327 | 中坡 | 长芒草、白羊草 |
| Steppe native | 2×2 | 109°04′E | 37°06′N | 1313 | 峁顶 | 铁杆蒿、白羊草 |
| species (SN) | 2×2 | 109°03′E | 37°09′N | 1314 | 峁顶 | 冷蒿、沙参棘豆 |
| 2×2 | 109°03′E | 37°09′N | 1317 | 中坡 | 长芒草、达乌里胡枝子、尧花 | |
| 2×2 | 109°20′E | 36°51′N | 1201 | 上坡 | 达乌里胡枝子、阿尔泰狗娃花 | |
| 森林草原区刺槐 | 10×10 | 109°15′E | 36°49′N | 1353 | 峁顶 | 杠柳、长芒草、铁杆蒿 |
| Forest-steppe | 10×10 | 109°15′E | 36°45′N | 1154 | 上坡 | 长芒草、丛生隐子草、铁杆蒿 |
| locust (EL) | 10×10 | 109°15′E | 36°44′N | 1248 | 上坡 | 悬钩子、灰灰菜、黑龙胆 |
| 森林草原区乡土植物 | 5×5 | 109°15′E | 36°49′N | 1354 | 中坡 | 狼牙刺、铁杆蒿 |
| Forest-steppe native | 5×5 | 109°15′E | 36°45′N | 1142 | 中坡 | 狼牙刺、铁杆蒿 |
| species (EN) | 5×5 | 109°15′E | 36°44′N | 1231 | 中坡 | 虎榛子 |
| 森林区刺槐 | 10×10 | 109°31′E | 36°26′N | 1324 | 上坡 | 黄刺玫、野芨芨草、野菊花 |
| Forest locust(FL) | 10×10 | 109°31′E | 36°25′N | 1382 | 上坡 | 黄刺玫、野芨芨草、披针苔草 |
| 10×10 | 109°16′E | 36°29′N | 1411 | 上坡 | 黄刺玫、野草莓 | |
| 10×10 | 109°17′E | 36°29′N | 1367 | 上坡 | 紫花地丁、杠柳 | |
| 森林区乡土植物 | 10×10 | 109°31′E | 36°25′N | 1391 | 上坡 | 栒子、披针苔草 |
| Forest native species(FN) | 10×10 | 109°31′E | 36°25′N | 1381 | 上坡 | 栒子、野樱桃、披针苔草 |
| 10×10 | 109°16′E | 36°29′N | 1391 | 上坡 | 栒子、披针苔草 | |
| 10×10 | 109°16′E | 36°29′N | 1396 | 上坡 | 披针苔草、牛奶子 |
为研究土壤微生物群落多样性关系随环境梯度的变化, 选择对土壤微生物有重要影响的气候和土壤因子。土壤含水率采用烘干法测定;土壤有机质含量使用重铬酸钾氧化法;全氮使用全自动凯氏定氮仪(FOSS全自动凯氏定氮仪KJELTEC 2300)测定;全磷含量采用硫酸高氯酸氧化法[16]。分别在每个样地1.5 m和地下0.5 m处安装iButton电子纽扣来记录样地不同高度下的温度和湿度。
1.4 土壤微生物功能多样性的测定土壤微生物功能多样性用Biolog Eco微孔板方法测定[17]。此方法操作简便, 培养时间短, 是研究土壤微生物群落功能多样性的主要方法之一[18]。在所测的结果中, 每孔读数减去第一个水空白的读数, 以除去本底干扰, 负值归零。利用每孔颜色平均变化率(Average Well-Color Development, AWCD)来计算土壤微生物代谢活性, 计算公式为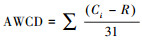
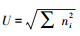
数据分析基于Microsoft Excel 2010, SPSS 18.0以及R 3.3.1进行。采用单因素方差分析和Tukey检验比较3个植被区内人工刺槐群落和乡土植物群落土壤之间的温度、湿度、理化性质、多样性指数及碳源利用类型的差异。采用Pearson相关系数分析土壤微生物功能多样性指数与环境因子的关系。显著性设为α=0.05。应用主成分分析获取土壤微生物主要利用碳源类型。应用典范对应分析碳源与环境因子的关系, 再使用蒙特卡洛置换来检验环境因子与底物碳源之间的显著性关系[19]。
2 结果和分析 2.1 土壤微生物群落代谢活性变化特征AWCD可以很好地表征微生物群落的整体代谢活动, 是土壤微生物活性及群落功能多样性的重要计算指标。本研究表明, 不同植被区刺槐与乡土植物群落AWCD值变化差异明显(图 1), 在13 d的培养期内, 总体上AWCD值随着培养时间的延长而增加, 即土壤微生物活性都随培养时间延长而提高。在培养初期的第一天内, AWCD值变化不明显, 培养一天之后, AWCD值急剧上升, 并随培养时间的延长, AWCD值逐渐增加, 之后趋于平缓。但是相较于表层, 深层10—20 cm土壤AWCD值上升更缓慢。

|
| 图 1 不同植被区刺槐与乡土植物0—10、10—20 cm土壤微生物AWCD值随培养时间变化 Fig. 1 AWCD changes with incubation time under different vegetation zone at 0—10 cm and 10—20 cm |
从刺槐植物群落来看, 随着温度升高、降雨量增加(从草原区到森林区), 刺槐群落AWCD值没有显著变化, 基本保持一致。但与乡土植物群落比较, 不同环境梯度下刺槐植物群落与乡土植物群落AWCD值差异明显。在草原区, 刺槐植物群落AWCD显著大于乡土植物群落AWCD值;在森林草原区, 刺槐植物群落AWCD与乡土植物群落AWCD值差异很小;但是在森林区, 刺槐植物群落AWCD却显著小于乡土植物群落AWCD值。相较于表层土壤, 深层10—20 cm土壤AWCD值差异均不显著。总体而言, 刺槐可以显著提高草原区土壤微生物代谢活性, 对森林草原区土壤微生物无显著影响, 但在森林区其土壤微生物代谢活性却显著低于乡土植物群落。
2.2 土壤微生物群落功能多样性指数分析根据第4天的AWCD值计算土壤微生物群落的丰富度指数、Shannon-Wiener指数、Simpson指数和McIntosh指数(表 3)。结果表明, 表层0—10 cm土壤微生物群落的丰富度指数、Shannon-Wiener指数、Simpson指数和McIntosh指数在不同植被区之间存在显著差异, 但在不同物种之间不存在显著性差异。人工刺槐林土壤微生物群落的丰富度指数、Shannon-Wiener指数、Simpson指数在草原区和森林草原区均大于乡土植物群落, 且在草原区差异显著, 但在森林区却小于乡土植物群落, 表明刺槐显著增加草原区土壤微生物常见种的数量, 以及显著提高土壤微生物多样性, 但在森林区效果并不明显。人工刺槐林土壤微生物群落的McIntosh指数在草原区显著大于乡土植物群落, 在森林草原区与乡土植物群落差异不明显, 但是在森林区却显著小于乡土植物群落, 表明刺槐显著提高草原区碳源的利用程度, 但在森林区对碳源的利用程度减弱。深层10—20 cm土壤微生物群落功能多样性指数变化规律基本与表层0—10 cm变化规律一致, 但该层土壤微生物群落的Shannon-Wiener指数、Simpson指数和McIntosh指数在不同植被区和物种之间都不存在显著性差异, 且在森林区深层土壤微生物群落中, 刺槐林土壤微生物群落McIntosh指数与乡土植物群落差异不显著, 这可能是由于随着土层深度增加, 土壤容重增大, 孔隙度减小, 土壤微生物受到枯落物的影响变小, 土壤理化性质的差异也变小, 导致土壤微生物对碳源利用程度的差距变小。
| 样点 Sites |
全碳 Total Carbon /(g/kg) |
全氮 Total Nitrogen /(g/kg) |
全磷 Total Phosphorus/(g/kg) |
土壤温度 Soil temperature/℃ |
土壤含水量 Moisture content/% |
|||||
| 0—10 cm | 10—20 cm | 0—10 cm | 10—20 cm | 0—10 cm | 10—20 cm | |||||
| SL | 7.68±1.65a | 4.81±0.57a | 0.42±0.08ab | 0.30±0.03a | 0.56±0.01a | 0.54±0.01a | 22.22±0.76a | 0.04±0.01a | ||
| SN | 7.01±2.08a | 4.92±1.44a | 0.38±0.11a | 0.28±0.08a | 0.58±0.04ab | 0.53±0.03a | 25.91±0.48b | 0.05±0.02a | ||
| EL | 15.45±6.95a | 7.33±1.24a | 0.88±0.36ab | 0.46±0.04a | 0.63±0.02b | 0.59±0.02a | 21.69±0.94a | 0.07±0.01ab | ||
| EN | 20.25±7.54a | 11.59±5.38ab | 1.10±0.40b | 0.70±0.35ab | 0.59±0.05ab | 0.55±0.07a | 23.19±2.64a | 0.08±0.02ab | ||
| FL | 14.01±3.77a | 8.52±2.53a | 0.82±0.10ab | 0.51±0.11a | 0.61±0.003ab | 0.58±0.002a | 21.10±0.88a | 0.13±0.03c | ||
| FN | 42.49±12.49b | 19.15±7.21b | 2.08±0.54abc | 1.03±0.31b | 0.61±0.02ab | 0.58±0.04a | 20.72±0.73a | 0.11±0.02bc | ||
| 同列不同小写字母表示差异显著(P < 0.05) | ||||||||||
| 林分 Stand |
0—10 cm | 10—20 cm | |||||||
| H | S | D | U | H | S | D | U | ||
| SL(单因素方差分析) | 2.69±0.22b | 11.91±5.89b | 0.91±0.02b | 3.16±1.58b | 2.47±0.43a | 10.62±4.11b | 0.88±0.07a | 3.08±1.08b | |
| SN | 2.24±0.50a | 3.17±2.55a | 0.84±0.07a | 1.16±0.77a | 2.29±0.63a | 3.17±2.48a | 0.82±0.13a | 1.35±0.80a | |
| EL | 2.71±0.23b | 11.86±5.15b | 0.91±0.03b | 3.10±1.43b | 2.57±0.16a | 9.78±2.49b | 0.90±0.02a | 2.39±0.42ab | |
| EN | 2.57±0.32ab | 10.71±5.68b | 0.89±0.04ab | 3.09±1.18b | 2.39±0.25a | 8.29±2.06ab | 0.88±0.03a | 3.50±1.52b | |
| FL | 2.79±0.24b | 13.8±6.29bc | 0.92±0.03b | 2.96±1.03b | 2.51±0.41a | 10.50±6.40b | 0.88±0.08a | 2.80±1.30ab | |
| FN | 2.97±0.17b | 19.64±4.72c | 0.94±0.01b | 4.99±1.27c | 2.66±0.27a | 10.22±5.09b | 0.91±0.03a | 2.60±1.59ab | |
| 物种(双因素方差分析) | 2.73 | 0.97 | 5.19* | 0.00 | 0.37 | 7.96** | 0.53 | 0.82 | |
| 植被区 | 9.91*** | 17.88*** | 9.30*** | 12.05*** | 1.35 | 3.94* | 2.02 | 2.12 | |
| 物种*植被区 | 5.75** | 11.24*** | 7.81** | 14.89*** | 0.98 | 4.75* | 2.10 | 7.57** | |
| 同列不同小写字母表示差异显著; *表示0.05水平显著差异, **表示0.01水平显著差异, ***表示0.001水平显著差异 | |||||||||
综合4种多样性指数能更客观地反映土壤微生物功能多样性变化规律。从刺槐植物群落来看, 随着温度升高、降雨量增加(从草原区到森林区), 刺槐植物群落土壤微生物功能多样性没有显著变化, 基本保持一致。但与乡土植物群落比较, 刺槐显著提高了草原区土壤微生物功能多样性, 却降低了森林区土壤微生物功能多样性。
2.3 土壤微生物群落碳源利用情况分析刺槐在不同植被区对土壤微生物群落碳源利用产生的影响不同(图 2)。所有研究样地对碳源的利用比例大致相同, 即CH(糖类Carbohydrates) > AA(氨基酸类Amino acids) > CA(羧酸类Carboxylic acids) > PL(多聚物类Polymers) > AM(胺类Amine) > MM(多酚类Polyhenols)。总体而言, 糖类和氨基酸类碳源是各样点土壤微生物利用的主要碳源。

|
| 图 2 不同植被区刺槐与乡土植物土壤微生物碳源利用 Fig. 2 The carbon source utilization by soil microbial community under different vegetation zones SL:草原区刺槐,steppe locust;SN:草原区乡土植物,steppe native species;EL:森林草原区刺槐,forest-steppe locust;EN:森林草原区乡土植物,forest-steppe native species;FL:森林区刺槐,forest locust;FN:森林区乡土植物,forest native species |
从刺槐植物群落来看, 随着温度升高、降雨量增加(从草原区到森林区), 刺槐植物群落土壤微生物主要利用的碳源类型从草原区的糖类, 过渡到森林草原区和森林区的糖类和氨基酸类。对于同一植被区内, 草原区刺槐林和乡土植物群落土壤微生物都主要利用糖类, 刺槐林土壤微生物对碳源利用效率明显高于乡土植物群落;森林草原区刺槐林和乡土植物群落土壤微生物都主要利用糖类和氨基酸类, 且对碳源的利用效率没有显著差异;森林区刺槐林土壤微生物主要利用糖类和氨基酸类, 而乡土植物群落主要利用糖类, 且乡土植物群落对碳源的利用效率明显高于人工刺槐群落。
2.4 土壤微生物群落碳源利用的主成分分析利用培养第4天后的单孔光密度值进行主成分分析(PCA), 共提取出7个主成分, 累计贡献率达到79.54%。其中, 第一主成分(PC1)的方差贡献率为39.06%, 第二主成分(PC2)的方差贡献率为15.22%。各不同植被区间主成分得分的方差分析表明, 以PC1、PC2对各处理作图, 来区分样品间的微生物群落特征。由图 3可知, PC1得分存在及显著差异(P=0.004), PC2得分不存在显著性差异。在PC1得分上, SN和SL、SN和FN之间存在显著差异, 其他各处理之间没有显著差异。在PC2得分上, 各组之间都不存在显著差异。

|
| 图 3 不同植被区刺槐和乡土植物土壤微生物碳源利用的主成分分析 Fig. 3 PCA for carbon source utilization of soil microbial communities under different vegetation zones |
PC1中载荷值大于0.8的碳源中糖类3种, 氨基酸类4种, 羧酸类1种, 胺类2种, 主要碳源利用类型为糖类、氨基酸类和羧酸类。PC2载荷值大于0.8的碳源中糖类2种, 主要碳源利用类型为糖类。所以, 在主成分分离中起主要贡献作用的是糖类和氨基酸类碳源。
2.5 环境因子对土壤微生物群落功能多样性和碳源利用的影响对第4天的平均颜色变化率AWCD值、S、H、D、U与环境因子进行相关分析结果如表 4所示, 土壤微生物群落功能多样性各指标与土壤全碳、全氮、全磷土壤含水量、地上湿度之间存在正相关关系, 与地上温度、土壤温度之间存在负相关关系。其中, 所有功能多样性指数与土壤碳、土壤氮、土壤温度之间的相关性达到显著水平, 除McIntosh指数外的其他4个多样性指标与地上温度呈显著相关, AWCD值和丰富度指数与土壤含水量间相关性显著。由此可知, 土壤全碳、土壤全氮、土壤温度对土壤微生物群落功能多样性的影响较大, 土壤含水量会影响土壤微生物活性。
| 相关系数 Correlation coefficient |
全碳 Total C |
全氮 Total N |
全磷 Total P |
土壤含水量 Moisture content |
地上温度 Temperature |
地上湿度 Humidity |
土壤温度 Soil temperature |
| AWCD | 0.397 | 0.438* | 0.202 | 0.485* | -0.545** | 0.400 | -0.610** |
| S | 0.523** | 0.528** | 0.403 | 0.460* | -0.657** | 0.503* | -0.806** |
| H | 0.456* | 0.442* | 0.399 | 0.339 | -0.637** | 0.395 | -0.677** |
| D | 0.438* | 0.427* | 0.270 | 0.217 | -0.451* | 0.313 | -0.639** |
| U | 0.488* | 0.485* | 0.367 | 0.238 | -0.401 | 0.309 | -0.711** |
| *, P < 0.05;**, P < 0.01; AWCD:平均颜色变化率,Average Well-Color Development;H:Shannon-Wiener指数,the Shannon-Wiener index;S:群落丰富度指数,the Richness index;D:Simpson指数,the Simpson index;U:McIntosh指数,the McIntosh index | |||||||
土壤微生物碳源利用类型与环境因子的典范对应分析(CCA)结果如图 4所示, 糖类碳源和土壤温度、地上温度呈正相关关系, 多分布在温度较高, 湿度较低的地方, 氨基酸类碳源多分布在湿度较高的固氮植物附近, 羧酸类碳源和土壤全碳、全氮、全磷呈正相关, 多分布在湿度较高的地方, 土壤全碳、土壤全氮、土壤全磷在底物分布中的影响较小。草原区的土壤温度和地上温度都高于其他植被区, 森林区的土壤含水量和地上湿度高于其他植被区。由此可知, 土壤含水量及温湿度对底物碳源利用类型有很大影响。

|
| 图 4 环境因子与底物碳源之间的关系 Fig. 4 The relationship between environmental factors and substrate T:温度, ST:土壤温度, H:湿度, MC:土壤含水量, C:土壤全碳, N:土壤全氮, P:土壤全磷, A1—A10:糖类, B1—B6:氨基酸, C1—C7:羧酸, D1—D4:多聚物, E1—E2:多酚, F1—F2:胺类 |
外来植物可以通过直接或间接的途径改变土壤微生物群落结构, 进而影响到整个生态系统的物质循环与能量转化过程, 对生态系统结构与功能产生重要影响。本研究表明, 作为外来物种, 刺槐在不同环境梯度(植被区)内对土壤微生物群落产生的影响不同:在草原区, 刺槐的引入显著提高了土壤微生物的代谢活性和功能多样性, 在森林草原区, 其对土壤微生物代谢活性和功能多样性也有所提高, 但在森林区的影响却显著低于乡土植物群落。刺槐对微生物功能多样性的影响存在明显的环境梯度效应。
产生这种环境梯度效应的原因, 一方面与不同环境条件下原有的植物群落结构变化有关[20-21], 另一方面与不同环境条件下微生境温度、湿度等环境条件变化有关。从刺槐对环境梯度变化的响应看:随着温度的升高、降雨量的增加(从草原区到森林区), 刺槐林下土壤微生物活性和功能多样性变化不显著, 而乡土植物群落土壤微生物活性和功能多样性则呈现增加的趋势。之所以产生这种差异, 可能是因为在不同环境梯度下, 人工刺槐植物群落结构变化较小, 植被类型基本相同, 土壤全磷和有机质含量基本一致(表 2), 其立地土壤微环境一致, 导致刺槐林下土壤微生物功能多样性变化不显著[22], 且刺槐作为一种固氮植物, 可以通过根瘤固氮作用, 提高土壤有机氮含量[23], 改善土壤养分状况, 促进微生物活性, 因而刺槐林下土壤微生物功能多样性在不同环境梯度下变化不显著。此外, 曹杨等[24]研究发现刺槐细根具有垂直分布并向四周伸展特征, 这种根系的分布优势, 促使刺槐具有更大的水分吸收空间, 较强的水分竞争能力, 对土壤水分含量要求降低, 对环境梯度变化响应不明显, 因此刺槐林下土壤微生物功能多样性变化不显著;而乡土植物群落的分布则主要受制于温度降雨梯度变化, 随着温度升高、降雨量增大, 乡土植物群落结构逐渐丰富, 植被类型逐渐向森林植被过渡, 林下植物多样性、物种丰富度和生物量都呈现增加趋势, 土壤养分氮含量也呈现增大趋势[25], 导致土壤微生物功能多样性呈增大趋势。也有研究发现乡土植物地下根系差异较大, 根系分泌物和死根提供给土壤微生物的能源物质也不同, 导致乡土植物群落土壤微生物功能多样性随环境梯度变化[26]。杨宁等[27]对不同植被恢复阶段土壤微生物群落多样性变化进行研究, 发现不同植被类型下土壤微生物活性和功能多样性的顺序为:乔木群落>灌木群落>草本群落, 这与我们的研究结果一致。
本研究还发现在同一环境梯度内, 刺槐与乡土植物群落土壤微生物代谢活性和功能多样性存在差异。在草原区和森林草原区内, 刺槐林土壤微生物代谢活性和功能多样性均高于乡土植物群落。而在森林区内, 刺槐林土壤微生物代谢活性和功能多样性却低于乡土植物群落。这可能是因为与乡土植物群落相比, 在草原区和森林草原区, 刺槐林群落结构丰富、物种丰富度高, 具有较高的叶片养分含量、根养分含量、较大的比叶面积和比根长等性状特征, 从而具有更强的水分养分竞争能力[28], 同时凋落物的分解使土壤养分得到更多的补充;且刺槐作为固氮物种, 可以通过固氮作用提高土壤有机质含量, 加之根系较浅且成扩散状分布于浅土层, 有利于改善表层土壤的结构, 同时根系分泌物及根际释放碳源均可为主要分布于土壤表层的微生物提供更多的营养基质, 改善了土壤养分状况[29-30], 因此刺槐林下土壤微生物代谢活性和功能多样性均高于同一环境梯度内乡土植物群落。除此之外, 刺槐在草原区和森林草原区对林下小气候(温度、湿度)的改善也会影响土壤微生物代谢活性和功能多样性。本研究监测数据表明(图 5), 7—9月份刺槐林地上温度与对照的草地群落和灌木群落虽差异不明显, 但均低于对照乡土群落, 湿度则与对照群落无差别;而刺槐林土壤温度显著低于对照的草地群落和灌木群落。且本研究发现土壤微生物代谢活性、功能多样性和碳源利用类型均受到土壤温度的影响, 此外, 土壤微生物碳源利用类型还受到土壤含水量的影响。张乃莉等[31]研究发现林下小气候的改变会直接影响植物的生长状况, 进而影响土壤微生物活性和多样性, 也会通过改变土壤温度来直接影响土壤微生物活性和多样性。Gordon等[32]、Xiang等[33]研究发现增加土壤含水量能提高土壤微生物活性。鲁顺保等[34]研究发现土壤含水量能够影响微生物对不同类型碳源利用的选择。土壤微生物的生长需要适宜的温湿度范围, 刺槐引入改善草原区和森林草原区温度状况, 因此刺槐林下土壤微生物生长代谢高于同一环境梯度内乡土植物群落。在森林区, 与乡土植物群落相比, 刺槐群落结构简单, 树种单一, 物种丰富度较低, 生物量整体上低于乡土植物群落, 对土壤微生物的影响较小[35], 且乡土植物多为高大乔木, 具有庞大根系, 根系分泌物和死根给微生物提供了丰富的能源物质, 对土壤微生物的影响较大[26], 上述这些条件导致刺槐林下土壤微生物代谢活性和功能多样低于乡土植物群落。除此之外, 刺槐在森林区改善林下小气候的功能不及乡土植物群落强。本试验研究数据表明, 7—9月份刺槐林地上和地下温度与乔木群落虽差异不明显, 但均高于对照的乔木群落, 乡土植物群落林下小气候更适合土壤微生物的生存, 土壤微生物代谢活性和功能多样性更高。

|
| 图 5 不同环境梯度下温湿度变化 Fig. 5 The change of temperature and humidity under different environmental gradient |
综上所述, 人工引种刺槐对土壤微生物群落的影响存在明显的环境梯度效应, 同时土壤温度、土壤含水量对土壤微生物群落结构起到调控作用, 在评价刺槐对土壤生态过程与功能的影响时, 不同环境梯度所产生的差异必须要考虑, 这样能更好的评价黄土丘陵区刺槐引入对自然植物群落土壤微生物的影响, 为黄土丘陵区合理进行植被恢复提供科学依据。
| [1] | Wardle D A, Bardgett R D, Klironomos J N, Setälä H, van der Putten W H, Wall D H. Ecological linkages between aboveground and belowground biota. Science, 2004, 304(5677): 1629–1633. DOI:10.1126/science.1094875 |
| [2] | 张燕燕, 曲来叶, 陈利顶, 卫伟. 黄土丘陵沟壑区不同植被类型土壤微生物特性. 应用生态学报, 2010, 21(1): 165–173. |
| [3] | Liao C Z, Peng R H, Luo Y Q, Zhou X H, Wu X W, Fang C M, Chen J K, Li B. Altered ecosystem carbon and nitrogen cycles by plant invasion:a meta-analysis. New Phytologist, 2008, 177(3): 706–714. DOI:10.1111/nph.2008.177.issue-3 |
| [4] | Knops J M H, Bradley K L, Wedin D A. Mechanisms of plant species impacts on ecosystem nitrogen cycling. Ecology Letters, 2002, 5(3): 454–466. DOI:10.1046/j.1461-0248.2002.00332.x |
| [5] | Hawkes C V, Wren I F, Herman D J, Firestone M K. Plant invasion alters nitrogen cycling by modifying the soil nitrifying community. Ecology Letters, 2005, 8(9): 976–985. DOI:10.1111/ele.2005.8.issue-9 |
| [6] | Lee M R, Bernhardt E S, van Bodegom P M, Cornelissen J H C, Kattge J, Laughlin D C, Niinemets V, Peñuelas J, Reich P B, Yguel B, Wright J P. Invasive species' leaf traits and dissimilarity from natives shape their impact on nitrogen cycling:a meta-analysis. New Phytologist, 2017, 213(1): 128–139. DOI:10.1111/nph.14115 |
| [7] | González-Muñoz N, Castro-Díez P, Parker I M. Differences in nitrogen use strategies between native and exotic tree species:predicting impacts on invaded ecosystems. Plant and Soil, 2013, 363(1/2): 319–329. |
| [8] | Funk J L. The physiology of invasive plants in low-resource environments. Conservation Physiology, 2013, 1(1): cot026. |
| [9] | Van Kleunen M, Weber E, Fischer M. A meta-analysis of trait differences between invasive and non-invasive plant species. Ecology Letters, 2010, 13(2): 235–245. DOI:10.1111/ele.2010.13.issue-2 |
| [10] | Ehrenfeld J G. Effects of exotic plant invasions on soil nutrient cycling processes. Ecosystems, 2003, 6(6): 503–523. DOI:10.1007/s10021-002-0151-3 |
| [11] | 胡婵娟, 傅伯杰, 靳甜甜, 刘国华. 黄土丘陵沟壑区植被恢复对土壤微生物生物量碳和氮的影响. 应用生态学报, 2009, 20(1): 45–50. |
| [12] | 翟辉, 张海, 张超, 周旭. 黄土峁状丘陵区不同类型林分土壤微生物功能多样性. 林业科学, 2016, 52(12): 84–91. DOI:10.11707/j.1001-7488.20161210 |
| [13] | 张海涵, 唐明, 陈辉, 杜小刚. 黄土高原5种造林树种菌根根际土壤微生物群落多样性研究. 北京林业大学学报, 2008, 30(3): 85–90. |
| [14] | 曾全超, 李鑫, 董扬红, 李娅芸, 安韶山. 黄土高原不同乔木林土壤微生物量碳氮和溶解性碳氮的特征. 生态学报, 2015, 35(11): 3598–3605. |
| [15] | Sherman C, Steinberger Y. Microbial functional diversity associated with plant litter decomposition along a climatic gradient. Microbial Ecology, 2012, 64(2): 399–415. DOI:10.1007/s00248-012-0037-7 |
| [16] | 鲍士旦. 土壤农化分析.第三版. 北京: 中国农业出版社, 2000. |
| [17] | 郑华, 欧阳志云, 方治国, 赵同谦. BIOLOG在土壤微生物群落功能多样性研究中的应用. 土壤学报, 2004, 41(3): 456–461. DOI:10.11766/trxb200309010320 |
| [18] | 张燕燕, 曲来叶, 陈利顶. Biolog EcoPlateTM实验信息提取方法改进. 微生物学通报, 2009, 36(7): 1083–1091. |
| [19] | Li Y Y, Jiao J X, Wang Y, Yang W, Meng C, Li B Z, Li Y, Wu J S. Characteristics of nitrogen loading and its influencing factors in several typical agricultural watersheds of subtropical China. Environmental Science and Pollution Research, 2015, 22(3): 1831–1840. DOI:10.1007/s11356-014-3446-y |
| [20] | 张超, 刘国彬, 薛萐, 王国梁. 黄土丘陵区不同植被类型根际微生物群落功能多样性研究. 草地学报, 2015, 23(4): 710–717. DOI:10.11733/j.issn.1007-0435.2015.04.007 |
| [21] | Waid J S. Does soil biodiversity depend upon metabiotic activity and influences?. Applied Soil Ecology, 1999, 13(2): 151–158. DOI:10.1016/S0929-1393(99)00030-X |
| [22] | 黄元元, 曲来叶, 曲秀春, 杜淑芹. 镜泊湖岩溶台地不同植被类型土壤微生物群落特征. 生态学报, 2012, 32(9): 2827–2836. |
| [23] | 胡延杰, 翟明普, 武觐文, 贾黎明. 杨树刺槐混交林及纯林土壤微生物数量及活性与土壤养分转化关系的研究. 土壤, 2002, 34(1): 42-46, 50-50. http://kns.cnki.net/KCMS/detail/detail.aspx?filename=tura200201007&dbname=CJFD&dbcode=CJFQ |
| [24] | 曹扬, 赵忠, 渠美, 成向荣, 王迪海. 刺槐根系对深层土壤水分的影响. 应用生态学报, 2006, 17(5): 765–768. |
| [25] | 邢肖毅, 黄懿梅, 安韶山, 张宏. 黄土高原沟壑区森林带不同植物群落土壤氮素含量及其转化. 生态学报, 2013, 33(22): 7181–7189. |
| [26] | 刘秉儒, 张秀珍, 胡天华, 李文金. 贺兰山不同海拔典型植被带土壤微生物多样性. 生态学报, 2013, 33(22): 7211–7220. |
| [27] | 杨宁, 邹冬生, 杨满元, 付美云, 林仲桂, 赵林峰. 衡阳紫色土丘陵坡地不同植被恢复阶段土壤微生物群落多样性的变化. 林业科学, 2016, 52(8): 146–156. |
| [28] | 王红霞. 黄土高原刺槐植物功能性状变化与环境适应的生态学意义[D]. 杨凌: 西北农林科技大学, 2016. http://cdmd.cnki.com.cn/Article/CDMD-10712-1016157288.htm |
| [29] | 胡婵娟, 郭雷, 刘国华. 黄土丘陵沟壑区不同植被恢复格局下土壤微生物群落结构. 生态学报, 2014, 34(11): 2986–2995. |
| [30] | 胡婵娟, 傅伯杰, 刘国华, 靳甜甜, 刘宇. 黄土丘陵沟壑区典型人工林下土壤微生物功能多样性. 生态学报, 2009, 29(2): 727–733. |
| [31] | 张乃莉, 郭继勋, 王晓宇, 马克平. 土壤微生物对气候变暖和大气N沉降的响应. 植物生态学报, 2007, 31(2): 252–261. DOI:10.17521/cjpe.2007.0029 |
| [32] | Gordon H, Haygarth P M, Bardgett R D. Drying and rewetting effects on soil microbial community composition and nutrient leaching. Soil Biology and Biochemistry, 2008, 40(2): 302–311. DOI:10.1016/j.soilbio.2007.08.008 |
| [33] | Xiang S R, Doyle A, Holden P A, Schimel J P. Drying and rewetting effects on C and N mineralization and microbial activity in surface and subsurface California grassland soils. Soil Biology and Biochemistry, 2008, 40(9): 2281–2289. DOI:10.1016/j.soilbio.2008.05.004 |
| [34] | 鲁顺保, 郭晓敏, 芮亦超, 周小奇, 陈成榕, 徐志红, 牛德奎. 澳大利亚亚热带不同森林土壤微生物群落对碳源的利用. 生态学报, 2012, 32(9): 2819–2826. |
| [35] | Ladygina N, Hedlund K. Plant species influence microbial diversity and carbon allocation in the rhizosphere. Soil Biology and Biochemistry, 2010, 42(2): 162–168. DOI:10.1016/j.soilbio.2009.10.009 |