 2018, Vol. 38
2018, Vol. 38文章信息
- 李凡, 徐炳庆, 吕振波, 王田田.
- LI Fan, XU Bingqing, LÜ Zhenbo, WANG Tiantian.
- 莱州湾鱼类群落优势种生态位
- Ecological niche of dominant species of fish assemblages in Laizhou Bay, China
- 生态学报. 2018, 38(14): 5195-5205
- Acta Ecologica Sinica. 2018, 38(14): 5195-5205
- http://dx.doi.org/10.5846/stxb201408281711
-
文章历史
- 收稿日期: 2014-08-28
- 修订日期: 2018-02-08





2. 烟台市水产研究所, 烟台 264003
2. Yantai Fisheries Research Institute, Yantai 264003, China
莱州湾位于渤海南端, 面积6215.4 km2, 湾内大部分区域水深小于10 m, 最大水深仅18 m[1]。莱州湾有黄河低盐水体注入, 营养物质丰富, 是我国近海渔业生产力最高的区域之一, 自20世纪50年代末以来, 随着人类活动(包括渔业捕捞、陆源污染、工程建设)的不断加剧, 莱州湾乃至整个黄、渤海的渔业资源已发生严重衰退, 鱼类群落结构发生了重大的变化, 低龄化、小型化明显[2-4]。
生态位(niche)理论在研究群落结构和功能、群落内物种间关系、生物多样性和群落动态演替等方面具有重要意义, 是解释群落中种间共存与竞争机制的基本理论之一[5]。生态位宽度和生态位重叠值的计算对了解各种类在群落中的地位、作用以及种类间的相互关系等具有重要意义[6], 是进行资源保护、可持续利用和生态修复的重要依据。
群落物种生态位的研究在陆生植物群落[7-9]和动物群落[10-12]应用比较广泛, 在水域生态系统中的研究主要集中于对潮间带和围垦滩涂大型底栖动物群落生态位的研究, 如对灵昆岛[13]、椒江口[14]、深圳湾[15]、渔山岛岩礁基质[16]潮间带以及围垦滩涂潮沟[17-18]大型底栖动物群落生态位的研究。应用生态位对鱼类群落的研究有于振海等[19]对黄海中南部鱼类群落主要鱼种时空生态位、李显森等[20]对长江口及其毗邻海域鱼类群落优势种的时空生态位以及韩东燕等[21]对胶州湾5种虾虎鱼类时空生态位和营养生态位的研究。本文利用2011年在莱州湾开展的鱼类底拖网调查资料, 开展了优势种的时空生态位和营养生态位研究, 以期为莱州湾渔业资源保护提供基础资料, 同时也为目前开展的生态修复提供技术支撑。
1 材料与方法 1.1 数据来源及调查方法数据来自2011年莱州湾(119°05′—120°00′E、37°12′—37°40′N)20站位(图 1)调查资料。调查时间分别为春季(5月)、夏季(8月)、秋季(10月)和冬季(12月)。调查船只功率260 kW, 调查网具为单船底拖网, 网口周长30.6 m, 囊网网目20 mm。每站拖曵1 h, 拖速3节。拖曵时, 网口宽度约8 m。调查均在白天进行。采集样品冰鲜保存, 带回实验室进行分类和生物学测定。记录每一种类的数量和重量, 将其换算为单位时间的生物量(kg/h)和丰度(尾/小时)。样品采样及分析均按《海洋调查规范第6部分:海洋生物调查》(GB/T 12763.6—2007)执行。鱼类名称以Nelson分类系统[22]为依据, 并参考Fishbase数据库[23]。

|
| 图 1 调查站位 Fig. 1 Sample station |
同位素测定样品来自夏季、秋季取样。待测样品带回实验室后解冻后用蒸馏水洗涤, 取适量背部白肌肉用于分析。所有样品-80℃冷冻后在冷冻干燥机(亿倍YB-FD-1)中真空干燥, 用石英研钵充分磨匀并加入1 mol/dm3的盐酸酸化处理后再烘干, 最后装入离心管中, 并放入干燥器中保存以备稳定同位素分析。实验样品的稳定同位素比值由中国林业科学院稳定同位素实验室测定。
1.2 分析方法 1.2.1 数据标准化根据各站位拖网时间和各鱼种的可捕系数, 对调查数据进行标准化[24]。中上层鱼类可捕系数取0.3, 近底层鱼类取0.5, 底层鱼类取0.8。
1.2.2 优势种和优势种的聚集强度采用Pinkas等的相对重要性指数(IRI)确定群落的优势种, 计算公式为[25]:
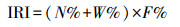
|
(1) |
式中, N为某一种类的渔获尾数占总渔获尾数的百分比;W为某一种类的生物量占总生物量的百分比;F为某一种类出现的站数占调查总站数的百分比。将IRI值大于等于1000的种类定义为优势种。
优势种的聚集强度是反映鱼类空间分布的一个指标, 当其为正值时, 说明该种空间格局非随机程度较大, 即有一定聚集, 正值数值越大, 则聚集强度越高。优势种聚集强度使用以下公式计算[26]:
丛生指数(clumping index):
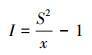
|
(2) |
平均拥挤度(mean crowding):
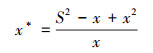
|
(3) |
式中, x为某种类的渔获尾数平均数, S2为方差。
1.2.3 生态位生态位宽度指数用Shannon-Wiener公式[27]:
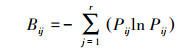
|
(5) |
生态位重叠指数用Pinaka公式[28]:
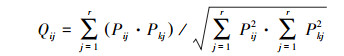
|
(6) |
式中, Pij=nij/Nij, 它代表种i在第j个资源状态下的个体数占该种所有个体数的比例, 其中j在计算时间生态位宽度、时间生态位的重叠时表示某个调查航次;j在计算空间生态位宽度、空间生态位重叠时, j表示某个调查站位。Qij为重叠指数, 其值介于0与1之间, 其值越大代表重叠越高, 当Qij>0.6时, 视为显著重叠[29]。
1.2.4 等级聚类对各站位鱼类尾数作log(x+1)处理, 使用Primer 6软件基于欧氏距离进行等级聚类分析。
1.2.5 主成分分析(PCA)利用主成分分析(Principal Component Analysis, PCA)影响鱼类群落的主要种类, 其图件绘制使用Canoco 4.5。
1.2.6 营养生态位分析稳定同位素值是该个体所有营养过程的累计, 因此, 测定稳定同位素是描述营养生态位的有效手段。使用δ15N比值范围(δ15N range, NR)、δ13C比值范围(δ13C range, CR)和生态位总面积(Total area, TA)度量营养生态位特征[30]:
δ15N比值范围:δ15N最大值和δ15N最小值之间的绝对距离。NR是营养垂直结构的表示, 通常更大的δ15N值变幅表明该种更宽泛的营养级和更大程度上的营养多元化。
δ13C比值范围:δ13C最大值和δ13C最小值之间的绝对距离。CR值较大表明拥有更多的食物来源(具备不同的δ13C值)和更多元的营养生态位。
生态位总面积:δ13C-δ15N二维坐标图中所有个体所围成的多边形面积, 是所有个体所占据的生态位的一个度量值, 也表征着营养多样性的总程度。
2 结果与分析 2.1 优势种及其聚集强度调查共捕获鱼类49种, 隶属于8目27科44属, 其中春季调查32种, 夏季33种, 秋季34种, 冬季21种。各季节优势种种类数不同, 为3至5种不等。春季, 3种优势种丛生指数均较高, 空间聚集特征明显, 赤鼻棱鳀(Thryssa kammalensis)丛生指数超过2, 集中聚集区主要在调查海域东北部。夏季, 各优势种丛生指数较低, 仅斑尾刺虾虎鱼(Acanthogobius ommaturus)接近1, 其在湾底的站位分布更为集中;各优势种平均拥挤度较高, 夏季第1优势种斑鱼祭(Konosirus punctatus)的平均拥挤度超过4000, 但其分布非常均匀, 因此丛生指数很低;蓝点马鲛(Scomberomorus niphonius)虽然成为该季节的优势种, 但其数量较少, 平均拥挤度较低, 分布也较为分散, 丛生指数较低。秋季, 青鳞小沙丁鱼(Sardinella zunasi)和赤鼻棱鳀平均拥挤度均较高, 但青鳞小沙丁鱼分布较为均匀, 丛生指数低, 而赤鼻棱鳀主要聚集于调查海域西部, 丛生指数较高。冬季, 黄鳍刺虾虎鱼(Acanthogobius flavimanus)数量大, 且集中分布在调查海域中部, 丛生指数和平均拥挤度均较高;绯鱼衔(Callionymus beniteguri)平均拥挤度虽较低, 但其集中分布在调查海域中部, 丛生指数亦较高, 斑尾刺虾虎鱼数量虽较多, 但其分布异常均匀, 丛生指数低但平均拥挤度虽高;矛尾虾虎鱼(Chaeturichthys stigmatias)丛生指数和平均拥挤度均较低。总体而言, 夏、冬季优势种的丛生指数较低, 春、秋季较高;夏、秋季优势种的平均拥挤度高, 冬、春季较低(表 1)。
| 季节 Season |
种类 Species |
相对重要性指数 Index of relative importance(IRI) |
重量百分比 Percentage of weight(W%) |
尾数百分比 Percentage of number(N%) |
丛生指数 Index of cluster(I) |
平均拥挤度 Average crowding(x*) |
| 春季Spring | 赤鼻棱鳀Thryssa kammalensis | 5348 | 34.3 | 42.1 | 2.032 | 295.9 |
| 绯鱼衔 Callionymus beniteguri |
3800 | 13.5 | 26.5 | 0.997 | 185.9 | |
| 青鳞小沙丁鱼 Sardinella zunasi |
1151 | 18.3 | 10.5 | 1.738 | 74.7 | |
| 夏季Summer | 斑鱼祭 Konosirus punctatus |
9474 | 54.0 | 40.8 | 0.071 | 4176.9 |
| 矛尾虾虎鱼 Chaeturichthys stigmatias |
3875 | 12.4 | 26.4 | 0.247 | 2703.2 | |
| 青鳞小沙丁鱼 Sardinella zunasi |
1614 | 4.7 | 11.4 | 0.565 | 1172.4 | |
| 蓝点马鲛 Scomberomorus niphonius |
1239 | 11.5 | 0.9 | 0.466 | 96.4 | |
| 斑尾刺虾虎鱼 Acanthogobius ommaturus |
1161 | 6.4 | 6.5 | 0.928 | 667.1 | |
| 秋季Autumn | 青鳞小沙丁鱼 Sardinella zunasi |
8771 | 51.8 | 51.4 | 0.873 | 7203.5 |
| 赤鼻棱鳀 Thryssa kammalensis |
4031 | 13.4 | 37.0 | 2.735 | 6541.7 | |
| 矛尾虾虎鱼 Chaeturichthys stigmatias |
2402 | 17.8 | 7.5 | 1.125 | 1286.9 | |
| 冬季Winter | 斑尾刺虾虎鱼 Acanthogobius ommaturus |
8594 | 63.8 | 22.2 | 0.047 | 109.4 |
| 黄鳍刺虾虎鱼 Acanthogobius flavimanus |
3386 | 5.5 | 36.9 | 1.251 | 183.0 | |
| 矛尾虾虎鱼 Chaeturichthys stigmatias |
2031 | 15.6 | 13.4 | 0.395 | 66.3 | |
| 绯鱼衔 Callionymus beniteguri |
1872 | 9.0 | 19.8 | 1.480 | 98.9 |
将生态优势度排序最高的12个种类丰度进行聚类(图 2), 结果表明聚类结果和生态优势度排序结果较吻合, 生态优势度最高的4个种聚为1组, 生态优势度排序第5、6位的2个种聚为1组, 其余6种聚为1组。协强系数(2D Stress)为0.1, 排序结果可信(图 2)。

|
| 图 2 以种间欧氏距离为基础的物种等级聚类图和NMDS排序图(数据经log(x+1)转换) Fig. 2 The hierarchical cluster dendrogram of fish species base on the Euclid distance(Data was transformed by log(x+1)) S1:青鳞小沙丁鱼Sardinella zunasi; S2:斑鱼祭Konosirus punctatus; S3:矛尾虾虎鱼Chaeturichthys stigmatias; S4:赤鼻棱鳀Thryssa kammalensis; S5:斑尾刺虾虎鱼Acanthogobius ommaturus; S6:绯鱼衔Callionymus beniteguri; S7:蓝点马鲛Scomberomorus niphonius; S8:短吻红舌鳎Cynoglossus joyneri; S9:鲬Platycephalus indicus; S10:银姑鱼Pennahia argentata; S11黄鲫Setipinna taty; S12:皮氏叫姑鱼Johnius belengerii |
图 3为各种类主成分分析(Principal components analysis, PCA)结果, 青鳞小沙丁鱼(S1)和赤鼻棱鳀(S4)分别是影响第一轴和第二轴的主要种类。第一轴解释72.9%的变量, 第二轴解释20.5%的变量, 累计可解释93.4%的变量。

|
| 图 3 莱州湾主要鱼类丰度PCA排序 Fig. 3 PCA ordination of fish abundance in Laizhou Bay |
表 2列出了莱州湾生态优势度最高的12个种类(占总生物量的97.2%和总尾数的98.4%)的生态位宽度和组成特征。时间生态位宽度最高的种类为绯鱼衔, 空间生态位宽度最高的种类为鲬(Platycephalus indicus), 时空二维生态位宽度最高种类为绯鱼衔。季节洄游种青鳞小沙丁鱼、斑鱼祭、赤鼻棱鳀因其时间生态位宽度较低, 时空生态位宽度并不高。鲬、绯鱼衔等周年定居种虽然数量比例较低, 但其在莱州湾全年均分布, 空间分布也较为均匀, 其生态位宽度较高。
| 代码Code | 种类Species | Nt | Ns | Nsp | IRI | W/% | N/% |
| S1 | 青鳞小沙丁鱼Sardinella zunasi | 0.45 | 2.36 | 1.06 | 3302 | 23.0 | 35.7 |
| S2 | 斑鱼祭Konosirus punctatus | 0.27 | 2.78 | 0.75 | 2768 | 32.8 | 15.4 |
| S3 | 矛尾虾虎鱼Chaeturichthys stigmatias | 0.72 | 2.86 | 2.05 | 2352 | 14.0 | 14.1 |
| S4 | 赤鼻棱鳀Thryssa kammalensis | 0.33 | 1.12 | 0.37 | 1841 | 7.5 | 24.6 |
| S5 | 斑尾刺虾虎鱼Acanthogobius ommaturus | 0.67 | 2.62 | 1.76 | 835 | 6.8 | 2.8 |
| S6 | 绯鱼衔Callionymus beniteguri | 1.29 | 3.33 | 4.29 | 233 | 1.3 | 1.6 |
| S7 | 蓝点马鲛Scomberomorus niphonius | 0.58 | 1.85 | 1.07 | 182 | 6.3 | 0.3 |
| S8 | 短吻红舌鳎Cynoglossus joyneri | 0.79 | 3.28 | 2.59 | 162 | 1.4 | 0.7 |
| S9 | 鲬Platycephalus indicus | 0.85 | 3.41 | 2.89 | 122 | 1.7 | 0.2 |
| S10 | 银姑鱼Pennahia argentata | 0.08 | 2.37 | 0.18 | 58 | 1.0 | 0.9 |
| S11 | 黄鲫Setipinna taty | 0.62 | 1.63 | 1.00 | 41 | 1.1 | 1.7 |
| S12 | 皮氏叫姑鱼Johnius belengerii | 0.36 | 2.11 | 0.76 | 40 | 0.5 | 0.6 |
| Nt:时间生态位temporal niche;Ns:空间生态位spatial niche;Nsp:时空生态位temporal-spatial niche;IRI:相对重要性指数index of relative importance;W%:重量百分比percent of weight;N%:尾数百分比percent of number | |||||||
从表 3可以看出, 时间生态位重叠值超过0.6的共有40组(其中重叠值超过0.9的35组), 占60.6%, 银姑鱼(Pennahia argentata)和蓝点马鲛重叠值达到1, 青鳞小沙丁鱼、赤鼻棱鳀与其他种类重叠值较低。空间生态位重叠值超过0.6的共有11组, 最大值出现于银姑鱼和皮氏叫姑鱼(Johnius belengerii)之间(0.861)。从表 4可以看出, 时空生态位显著重叠发生在银姑鱼和皮氏叫姑鱼(0.798)、银姑鱼和黄鲫(Setipinna taty)(0.798)、黄鲫和皮氏叫姑鱼(0.793)、短吻红舌鳎(Cynoglossus joyneri)和皮氏叫姑鱼(0.771)、矛尾虾虎鱼和皮氏叫姑鱼(0.770)、斑鱼祭和蓝点马鲛(0.763)以及斑鱼祭和短吻红舌鳎(0.625)之间, 其他种类间时空生态位重叠值均低于0.6。
| 种类 Species |
生态位重叠Niche overlap | |||||||||||
| S1 | S2 | S3 | S4 | S5 | S6 | S7 | S8 | S9 | S10 | S11 | S12 | |
| S1 | 0.434 | 0.268 | 0.377 | 0.147 | 0.380 | 0.246 | 0.454 | 0.186 | 0.202 | 0.137 | 0.218 | |
| S2 | 0.225 | 0.576 | 0.360 | 0.306 | 0.534 | 0.764 | 0.634 | 0.537 | 0.395 | 0.248 | 0.406 | |
| S3 | 0.569 | 0.929 | 0.071 | 0.397 | 0.370 | 0.487 | 0.797 | 0.497 | 0.436 | 0.157 | 0.561 | |
| S4 | 0.992 | 0.107 | 0.465 | 0.070 | 0.656 | 0.140 | 0.180 | 0.159 | 0.133 | 0.042 | 0.057 | |
| S5 | 0.227 | 0.987 | 0.922 | 0.110 | 0.421 | 0.192 | 0.345 | 0.650 | 0.257 | 0.029 | 0.153 | |
| S6 | 0.285 | 0.568 | 0.591 | 0.246 | 0.631 | 0.306 | 0.466 | 0.657 | 0.320 | 0.109 | 0.238 | |
| S7 | 0.167 | 0.998 | 0.906 | 0.047 | 0.985 | 0.553 | 0.521 | 0.332 | 0.243 | 0.135 | 0.209 | |
| S8 | 0.350 | 0.987 | 0.965 | 0.238 | 0.981 | 0.658 | 0.977 | 0.575 | 0.715 | 0.514 | 0.779 | |
| S9 | 0.598 | 0.911 | 0.995 | 0.501 | 0.902 | 0.645 | 0.885 | 0.959 | 0.528 | 0.270 | 0.439 | |
| S10 | 0.175 | 0.999 | 0.909 | 0.055 | 0.985 | 0.554 | 1.000 | 0.978 | 0.889 | 0.861 | 0.800 | |
| S11 | 0.531 | 0.945 | 0.999 | 0.425 | 0.934 | 0.584 | 0.924 | 0.975 | 0.992 | 0.927 | 0.835 | |
| S12 | 0.242 | 1.000 | 0.935 | 0.124 | 0.987 | 0.581 | 0.997 | 0.990 | 0.919 | 0.997 | 0.950 | |
| 等角线上为空间生态位重叠值, 等角线下为时间生态位重叠值 | ||||||||||||
| 种类 Species |
时空生态位重叠值Spatio-temporal niche overlap | ||||||||||
| S2 | S3 | S4 | S5 | S6 | S7 | S8 | S9 | S10 | S11 | S12 | |
| S1 | 0.098 | 0.153 | 0.374 | 0.033 | 0.108 | 0.041 | 0.159 | 0.111 | 0.035 | 0.073 | 0.053 |
| S2 | 0.535 | 0.039 | 0.302 | 0.303 | 0.762 | 0.625 | 0.489 | 0.395 | 0.235 | 0.406 | |
| S3 | 0.033 | 0.366 | 0.219 | 0.441 | 0.769 | 0.495 | 0.397 | 0.157 | 0.525 | ||
| S4 | 0.008 | 0.161 | 0.007 | 0.043 | 0.080 | 0.007 | 0.018 | 0.007 | |||
| S5 | 0.265 | 0.189 | 0.338 | 0.586 | 0.253 | 0.027 | 0.151 | ||||
| S6 | 0.169 | 0.307 | 0.424 | 0.177 | 0.064 | 0.138 | |||||
| S7 | 0.509 | 0.294 | 0.243 | 0.124 | 0.209 | ||||||
| S8 | 0.551 | 0.699 | 0.501 | 0.771 | |||||||
| S9 | 0.469 | 0.268 | 0.403 | ||||||||
| S10 | 0.798 | 0.798 | |||||||||
| S11 | 0.793 | ||||||||||
青鳞小沙丁鱼δ15N值变幅最大(7.1), 皮氏叫姑鱼δ15N值变幅最小(1.7), 表明青鳞小沙丁鱼拥有更宽泛的营养级。斑鱼祭δ13C值变幅最大(9.2), 蓝点马鲛和皮氏叫姑鱼δ13C值变幅较小(< 2), 表明斑鱼祭有更多元的食物来源, 而蓝点马鲛、皮氏叫姑鱼食物来源单一。青鳞小沙丁鱼和斑鱼祭生态位总面积较大, 均超过20, 皮氏叫姑鱼最小, 仅1.38(表 5)。
| 代码Code | 种类Species | δ15N值变幅 δ15N range (NR) |
δ13C值变幅 δ13C range (CR) |
总面积 Total area (TA) |
| S1 | 青鳞小沙丁鱼Sardinella zunasi | 7.1 | 4.5 | 20.01 |
| S2 | 斑鱼祭Konosirus punctatus | 5.4 | 9.2 | 27.74 |
| S3 | 矛尾虾虎鱼Chaeturichthys stigmatias | 5.5 | 3.9 | 12.30 |
| S4 | 赤鼻棱鳀Thryssa kammalensis | 3.5 | 4.2 | 8.07 |
| S5 | 斑尾刺虾虎鱼Acanthogobius ommaturus | 5.9 | 3.0 | 9.73 |
| S6 | 绯鱼衔Callionymus beniteguri | 4.4 | 3.7 | 10.48 |
| S7 | 蓝点马鲛Scomberomorus niphonius | 3.1 | 1.8 | 2.22 |
| S8 | 短吻红舌鳎Cynoglossus joyneri | 4.7 | 3.5 | 7.94 |
| S9 | 鲬Platycephalus indicus | 5.1 | 4.9 | 17.52 |
| S10 | 银姑鱼Pennahia argentata | 4.7 | 3.8 | 13.21 |
| S11 | 黄鲫Setipinna taty | 2.9 | 2.9 | 3.80 |
| S12 | 皮氏叫姑鱼Johnius belengerii | 1.7 | 1.8 | 1.38 |
图 4列出了时空生态位重叠超过0.6的种类间营养生态位分布图。从图中可以看出, 银姑鱼与皮氏叫姑鱼镶嵌在一起, 但银姑鱼生态位面积更大, 涵盖了皮氏叫姑鱼的生态位。银姑鱼与黄鲫营养生态位重叠较高, 银姑鱼主要占据偏上方的区域, 生态位总面积更大, 黄鲫主要分布在偏下方区域;黄鲫与皮氏叫姑鱼2者营养生态位略有重叠, 但重叠程度不高, 黄鲫营养生态位分布偏下方, 皮氏叫姑鱼偏上方;短吻红舌鳎与皮氏叫姑鱼2者营养生态位重叠较大, 短吻红舌鳎的生态位基本包含皮氏叫姑鱼的生态位, 其生态位总面积远大于皮氏叫姑鱼;矛尾虾虎鱼生态位几乎涵盖皮氏叫姑鱼生态位, 但矛尾虾虎鱼生态位面积更大;斑鱼祭和蓝点马鲛营养生态位重叠较小, 斑鱼祭的生态位总面积更大, 分布在图右下侧, 蓝点马鲛生态位总面积小, 分布在, 重叠较小;斑鱼祭和短吻红舌鳎营养生态位镶嵌在一起, 斑鱼祭生态位总面积略大。

|
| 图 4 莱州湾鱼类的营养生态位 Fig. 4 Trophic niches of fishes in Laizhou Bay |
春季, 优势种的产卵群体通常集群, 密集分布于浅水区(特别是黄河口以南的浅水区), 但数量较少, 因此丛生指数较高而平均拥挤度较低。夏季, 产卵后亲鱼和大量当年生幼鱼在调查海域分散索饵, 分布范围广且较平均, 因此这一时期优势种丛生指数较低而平均拥挤度较高。秋季, 调查海域洄游性鱼类尚未离开, 并且更多的当年生幼鱼进入网具规格, 因此平均拥挤度达到全年最高值, 而丛生指数则和种群空间分布密切相关, 如赤鼻棱鳀和青鳞小沙丁鱼虽然尾数均较高, 但赤鼻棱鳀主要集中分布在调查海域西北部, 而青鳞小沙丁鱼分布较均匀, 其丛生指数大大低于赤鼻棱鳀。冬季, 莱州湾水温已降至5℃左右, 洄游种类已离开莱州湾, 该时期群落优势种为虾虎鱼等定居种, 但数量较少, 因此平均拥挤度较低, 其中黄鳍刺虾虎鱼由于集中分布在10 m等深线附近, 平均拥挤度虽然较低但丛生指数较高。
3.2 生态位宽度时间生态位宽度值是物种在调查海域出现时间的度量值[11]。本研究中绯鱼衔在4季度调查中均出现, 并且其种群数量季节间差异较小, 成为时间生态位宽度最高的种类。而优势种中的青鳞小沙丁鱼、赤鼻棱鳀等洄游性鱼类在冬季数量较少, 矛尾虾虎鱼和斑尾刺虾虎鱼则因为春季数量非常低(产卵后亲体死亡), 因此这些优势种的时间生态位宽度并不高。
空间生态位宽度值是物种空间维度分布范围的度量值[11]。本研究中鲬的空间生态位宽度最高, 其次为绯鱼衔和短吻红舌鳎, 这些种类是莱州湾定居种, 空间分布较广(出现频率均超过60%)且较均匀。出现频率最高的斑尾刺虾虎鱼(87.5%)和矛尾虾虎鱼(83.8%), 由于空间分布极不均匀, 其空间生态位宽度并不高。
时空生态位宽度值是反映物种对环境适应性及资源利用广泛性的度量值[11]。本研究中绯鱼衔、鲬、短吻红舌鳎和矛尾虾虎鱼等底层鱼类时空生态位宽度较高(>2), 这些种类均为莱州湾周年定居种且在整个调查海域均有分布, 集群特征不明显;青鳞小沙丁鱼、斑鱼祭、黄鲫等洄游性种类, 它们春季进入莱州湾, 夏、秋季在莱州湾广泛分布, 而在秋末洄游离开莱州湾, 时间生态位宽度较低, 它们的时空生态位宽度较低。
3.3 不同指标的生态含义相对重要性指数、丛生指数、生态位宽度、聚类、排序、PCA分析所反映的群落特征各不相同, 均从某一或某几个方面反映群落结构特征。相对重要性指数综合考虑鱼类重量比例、尾数比例以及出现频率, 但其并不考虑时间、空间分布的均衡性;平均拥挤度指数仅考虑鱼类的尾数, 能在一定程度上反映空间分布均衡性;时空二维生态位宽度反映鱼类数量在时间、空间分布均匀度。等级聚类、排序与PCA分析, 反映的是物种分布和数量状况以及对群落构成的影响程度。本文中, 等级聚类、排序以及PCA分析得到结果与优势种分析得到结果较为接近, 而与时空生态位宽度分析结果差异较大, 但均表明莱州湾鱼类群落结构受洄游鱼类的影响较大。
3.4 优势种类的竞争分析经典共存理论认为, 在资源有限的环境中, 完全的竞争者不能共存, 生态位相似的物种之间会发生激烈的种间竞争, 处于劣势的物种常常会面临被淘汰的风险, 因此生态地位相似的物种为了实现共存, 各优势种类必将通过生态位分化以达到减轻竞争、共存的目的[21]。根据定义, Pinaka生态位重叠值范围为0—1, 当重叠值为0时, 表明2种生物对资源的利用方式完全不同或者在所研究的尺度下未同时出现;当重叠着为1时, 表明2种生物对资源利用方式完全相同或2者完全排斥, 不能共存[14]。有些生物因利用独特的生境而导致重叠值为0, 或因共栖、互利或共生而导致重叠值为1, 本研究中并不存在这种关系, 各种类间生态位重叠在0.007—0.798之间, 表明既无资源利用完全相同的种, 也无完全排斥的种。
由于莱州湾水深较浅, 各种类之间或多或少存在一定的食物重叠, 传统意义的中上层鱼类, 如赤鼻棱鳀、黄鲫、斑鱼祭、蓝点马鲛也摄食一部分底栖虾蟹类、双壳类等底层饵料生物, 传统意义的底层鱼类, 如矛尾虾虎鱼也会摄食部分浮游动物[31]。各种类食物虽有所重叠, 但由于摄食对象广泛, 又形成了一定程度的营养生态位分化。功能群(通常根据摄食习性划分)由一群生态学特征上很相似的物种组成, 彼此之间的生态位有明显重叠, 因而同一功能群内种间竞争很剧烈, 而与其他功能群之间种间竞争不明显。本研究中青鳞小沙丁鱼、赤鼻棱鳀和黄鲫同属浮游生物食性种团, 绯鱼衔和短吻红舌鳎属底栖生物种团, 矛尾虾虎鱼、斑尾刺虾虎鱼、鲬、银姑鱼和皮氏叫姑鱼则同属底栖动物/游泳动物食性种团[32]。在优势种类之中, 同属相同功能群且时空生态位重叠和营养生态位重叠均较高的情况仅发生在皮氏叫姑鱼与银姑鱼、皮氏叫姑鱼与矛尾虾虎鱼之间。皮氏叫姑鱼的营养生态位较窄, 加之种群规模较小, 对其他种类竞食压力不大, 对群落影响较小, 但其营养生态位和多个种类重叠, 因而其种群规模扩增可能会受到极大限制。生态位重叠仅是种间竞争的前提条件, 主要种类之间存在的生态位重叠, 但可以通过栖息水层不同、空间分布差异、进出莱州湾时间不同等时空生态位的分化, 以及营养生态位的分化(摄食对象的不同), 有效避免直接竞争。葛宝明等[13]对灵昆岛、焦海峰[16]对渔山岛潮间带大型底栖动物研究表明, 生态位重叠值较大种类可以通过食性、生活型等分化, 达到共存。因此, 生态位重叠导致的竞争并不是莱州湾群落结构现状的决定性因素, 群落变化更多是捕捞过度[2-4]、污染加剧[1]和生境丧失的综合结果。
群落物种组成情况反映了动物对生境的适应性, 生态位分化包括栖息地分化、领域分化、食性分化、生理分化以及体型分化, 生境复杂程度越高, 能够提供的生态位分化程度越高。本研究中时空生态位重叠值普遍较高的结果也在一定程度上反映了莱州湾鱼类群落生境同质程度较高的现实, 在进行生态修复时亦应着重考虑。
本研究根据优势种的时空生态位、营养生态位, 研究了莱州湾鱼类群落优势种的生态位现状。在分析营养生态位的主要依据是生态位面积(TA), TA是由稳定同位素极值决定的, 而极值易受测量误差的影响。下一步工作可根据优势种的时空分布, 结合胃含物分析法和同位素分析法开展营养生态位研究, 从时间-空间-营养三维开展莱州湾鱼类生态位重叠分析, 能够更好的反映种间的竞争。
| [1] | 侯英民. 山东海情. 北京: 海洋出版社, 2010. |
| [2] | 金显仕, 邓景耀. 莱州湾渔业资源群落结构和生物多样性的变化. 生物多样性, 2000, 8(1): 65–72. |
| [3] | Jin X S, Shan X J, Li X S, Wang J, Cui Y, Zuo T. Long-term changes in the fishery ecosystem structure of Laizhou Bay, China. Science China Earth Sciences, 2013, 56(3): 366–374. DOI:10.1007/s11430-012-4528-7 |
| [4] | 李凡, 张焕君, 吕振波, 徐炳庆, 郑亮. 莱州湾游泳动物群落种类组成及多样性. 生物多样性, 2013, 21(5): 537–546. |
| [5] | 张金屯. 数量生态学(第二版). 北京: 科学出版社, 2011. |
| [6] | 徐春燕, 俞秋佳, 徐凤洁, 胡雪芹, 由文辉. 淀山湖浮游植物优势种生态位. 应用生态学报, 2012, 23: 2550–2558. |
| [7] | 王仁忠. 放牧影响下羊草草地主要植物种群生态位宽度与生态位重叠的研究. 植物生态学报, 1997, 21(04): 9–16. |
| [8] | 李峰, 谢永宏, 陈心胜, 邓正苗, 李旭. 黄河三角洲湿地水生植物组成及生态位. 生态学报, 2009, 29: 6257–6265. DOI:10.3321/j.issn:1000-0933.2009.11.061 |
| [9] | 钱逸凡, 伊力塔, 胡军飞, 张超, 余树全, 沈露, 彭东琴. 普陀山主要植物种生态位特征. 生态学杂志, 2012, 31(3): 561–568. |
| [10] | 马英, 魏有文, 罗军, 鲁亮, 刘起勇. 青海省海东地区小型兽空间生态位分析. 动物学杂志, 2011, 46(04): 126–130. |
| [11] | 李锐, 李生才, 田瑞钧. 菜田蜘蛛群落组成及生态位分析. 蛛形学报, 2007, 16(02): 116–120. DOI:10.3969/j.issn.1005-9628.2007.02.010 |
| [12] | 杜超, 赵惠燕, 高欢欢, 李建峰, 罗坤, 胡祖庆, 胡想顺. 猕猴桃园节肢动物群落重建及主要类群的生态位. 生态学报, 2011, 31: 3246–3254. |
| [13] | 葛宝明, 鲍毅新, 郑祥, 程宏毅. 灵昆岛潮间带大型底栖动物群落结构与生态位分析. 生态学报, 2005, 5: 3037–3043. DOI:10.3321/j.issn:1000-0933.2005.11.034 |
| [14] | 赵永强, 曾江宁, 高爱根, 陈全震, 寿鹿, 廖一波, 黄逸君. 椒江口潮间带大型底栖动物的生态位. 应用生态学报, 2009, 20: 1176–1183. |
| [15] | 厉红梅, 蔡立哲, 林丽珠, 姚建彬. 深圳湾潮间带底栖动物群落结构的等级聚类与非度量多维标度排序. 厦门大学学报(自然科学版), 2001, 40: 735–740. DOI:10.3321/j.issn:0438-0479.2001.03.015 |
| [16] | 焦海峰, 施慧雄, 尤仲杰, 黄滨. 渔山岛岩礁基质潮间带大型底栖动物优势种生态位. 生态学报, 2011, 31: 3928–3936. |
| [17] | 葛宝明, 郑祥, 程宏毅, 鲍毅新, 陈良. 灵昆岛围垦滩涂潮沟大型底栖动物群落和物种生态位分析. 水生生物学报, 2007, 31: 675–681. DOI:10.3321/j.issn:1000-3207.2007.05.011 |
| [18] | 胡知渊, 鲍毅新, 葛宝明, 李欢欢. 围垦滩涂潮沟秋季大型底栖动物群落和生态位分析. 动物学报, 2006, 52: 800–809. |
| [19] | 于振海, 金显仕, 李显森. 黄海中南部主要鱼种的生态位分析黄海中南部主要鱼种的生态位分析. 渔业科学进展, 2010, 31(6): 1–8. |
| [20] | 李显森, 于振海, 孙珊, 金显仕. 长江口及其毗邻海域鱼类群落优势种的生态位宽度与重叠. 应用生态学报, 2013, 24: 2353–2359. |
| [21] | 韩东燕, 薛莹, 纪毓鹏, 徐宾铎, 刘贺, 麻秋云. 胶州湾5种虾虎鱼类的营养和空间生态位. 中国水产科学, 2013, 20(1): 148–156. |
| [22] | Nelson J S. Fishes of the World. 4th edition. New Jersey:John Wiley & Sons Inc, 2006. |
| [23] | Froese, R. and D. Pauly. Editors. 2017. FishBase. World Wide Web electronic publication. www. fishbase. org, version (10/2017). |
| [24] | 詹秉义. 渔业资源评估. 北京: 中国农业出版社, 1995. |
| [25] | Pinkas L, Oliphant MS, Iverson ILK. Food habits of albacore, bluefin tuna and bonito in California waters. Fish Bulletin, 1971, 152: 1–105. |
| [26] | 林龙山, 郑元甲, 马春艳. 台湾海峡夏秋季游泳动物资源分布及群落结构. 应用生态学报, 2005, 16(10): 1948–1951. DOI:10.3321/j.issn:1001-9332.2005.10.029 |
| [27] | Shannon C E, Weiner W. The mathematical theory of vommunication. Urbana: University of Illinois Press, 1949. |
| [28] | Pianka E R. The structure of lizard communities. Annual Review of Ecology and Systematics, 1973, 4: 53–74. DOI:10.1146/annurev.es.04.110173.000413 |
| [29] | Krebs C J. Ecological methodology. Menlo Park: Addison-Welsey, 1999. |
| [30] | Layman C A, Arrington D A, Mont A C G, Post P M. Can stable isotope ratios provide for community-wide measures of trophic structure?. Ecology, 2007, 88(1): 42–48. DOI:10.1890/0012-9658(2007)88[42:CSIRPF]2.0.CO;2 |
| [31] | 张波, 吴强, 金显仕. 莱州湾鱼类群落的营养结构及其变化. 渔业科学进展, 2013, 34(2): 1–9. |
| [32] | 李凡, 徐炳庆, 马元庆, 吕振波, 王田田. 莱州湾鱼类群落同功能种团的季节变化. 生态学报, 2014, 34(07): 1736–1745. |