 2018, Vol. 38
2018, Vol. 38文章信息
- 周子建, 江源, 董满宇, 陶岩, 王明昌, 丁新原.
- ZHOU Zijian, JIANG Yuan, DONG Manyu, TAO Yan, WANG Mingchang, DING Xinyuan.
- 长白山北坡不同海拔红松径向生长-气候因子关系对气温突变的响应
- Response of the relationship between radial growth and climatic factors to abrupt change of temperature along an altitudinal gradient on the northern slope of Changbai Mountain, Northeast China
- 生态学报. 2018, 38(13): 4668-4676
- Acta Ecologica Sinica. 2018, 38(13): 4668-4676
- http://dx.doi.org/10.5846/stxb201706171104
-
文章历史
- 收稿日期: 2017-06-17
- 网络出版日期: 2018-03-16





2. 北京师范大学地理科学学部, 北京 100875;
3. 东北师范大学地理科学学院, 长春 130024
2. Faculty of Geographical Science, Beijing Normal University, Beijing 100875, China;
3. School of Geographical Science, Northeast Normal University, Changchun 130024, China
近130年间, 全球海陆表面平均温度呈线性上升趋势, 中高纬地区是全球变暖最明显的地区之一[1]。运用树木年轮学方法, 建立树轮年表与气候因子的相关关系, 能够揭示气温升高对区域植被的影响。由于树木寿命长, 其生长会对升温表现出不同的响应[2]。Herrero等对地中海盆地欧洲赤松(Pinus sylvestris)和欧洲黑松(Pinus nigra)的研究发现, 该区夏季气温升高引起了树木生长下降和死亡[3]。Vila等对法国圣博姆山不同海拔地中海松(Pinus halepensis)的研究发现, 气温上升有助于其在各个海拔生境中的径向生长[4]。然而, 随着全球变暖加剧, 越来越多的研究发现北半球高纬度地区树木径向生长与温度相关性减弱的现象[5-6]。在全球气候变化的背景下, 非常有必要在不同区域内检验树木径向生长对限制气候因子响应关系的稳定性, 为森林资源管理等提供理论依据。
近50年来我国东北地区温度升高速率为0.30℃/10 a, 高于全球和全国的平均增温速率[7]。长白山地区地处中高纬度, 位于我国东北部, 是我国较早开展树木年轮学研究的区域之一。在早期, 利用树木年轮资料对温度和降水等气象因子的重建工作已取得了很大进展[8-9]。随着全球变暖问题的加剧, 学者们开展了关于温度对树木生长影响的研究[10]。目前的研究主要集中在通过不同海拔、不同树种、不同林型及不同年龄树木径向生长与气候因子的相关关系, 推测未来长白山地区树木径向生长变化趋势[11-13]。但是针对气温突变背景下, 对比分析不同海拔树木径向生长-气候因子关系的稳定性探讨还较少。因此, 本文以红松(Pinus koraiensis)为研究对象, 揭示其在不同海拔生境中径向生长—气候因子关系对气温突变的响应差异, 为预测红松径向生长的动态变化及区域气候重建等研究提供科学依据。
1 材料与方法 1.1 研究区概况长白山(41°23′—42°36′ N, 126°55′—129°08′ E)位于我国吉林省东南部(图 1), 最高海拔约为2700 m。本区气候属于温带大陆季风型高山气候, 冬季寒冷而漫长, 夏季温暖多雨而短暂。由于山体高, 气候随海拔高度变化很大, 随海拔的升高, 年降水量变幅在800—1400 mm之间, 年均温度在-7.6—3.7℃之间。

|
| 图 1 采样点和气象站分布 Fig. 1 Sample sites and meteorological stations |
红松是长白山针阔混交林优势树种, 在北坡海拔分布约为700—1300 m。用于本研究的红松树轮样本均采自长白山北坡, 海拔分别为745、1134 m和1280 m(图 1)。3个样地的郁闭度约为0.75—0.85, 平均树高约为20—25 m。在每棵树胸高处用生长锥钻取2个样芯(少数样树采集1个样芯)。采样时间为2015年8月。
1.3 气象数据整理采样点附近有二道、东岗、天池3个气象站点(图 1)。距离采样点最近的天池气象站自1989年冬季撤站, 造成冬季气象观测数据缺失。1958—2014年间东岗和二道气象站对应月份的平均气温和降水量相关系数平均分别达到0.93和0.79(P < 0.01)。因此, 气象资料选用了与采样点海拔较为接近的东岗气象站。考虑到树木生长对气候因子响应的“滞后现象”, 选取前一年9月到当年8月的气候因子进行分析。
1.4 树轮年表建立将红松树轮样芯在实验室里依次进行固定、打磨、交叉定年。使用LinTab 5年轮分析仪器在0.001 mm精度上测量树轮宽度。运用COFECHA程序对交叉定年作进一步检验[14], 剔除问题较大的样芯。运用ARSTAN软件, 采用步长30年的样条函数进行滑动去除与树龄相关的生长趋势和树木之间干扰竞争等生长趋势的影响, 并对去趋势后的序列以双重平均法合成标准年表(STD)[15]。
1.5 数据分析采用Mann-Kendall方法对东岗气象站1957—2014年的年均气温进行突变检验, 确定研究区可能发生气温突变的年份。
通过SPSS程序计算树轮年表与气候因子的Pearson相关系数, 分析树木径向生长对气候因子的响应。
通过μ值计算检验了温度突变前后两个时间段树轮年表与相同气候因子相关系数的变化情况。μ值是标准正态分布的分位数, 计算公式如下:
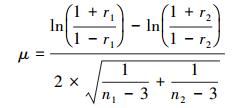
|
式中, r1和r2代表两个时段树轮年表与气候因子的相关系数, n1和n2代表两个时段的样本个数, 如果|μ| > 1.96则代表两个时段树轮年表与相同气候因子相关系数有显著差异(P < 0.05)[16]。
利用Dendroclim 2002程序的滑动相关函数功能分析了树轮年表对气候因子响应随时间变化的动态关系(滑动窗口25年)[17]。
2 研究结果 2.1 气温与降水变化对东岗气象站1957—2014年的年均气温进行Mann-Kendall检验, 结果表明1987年为研究区年均气温升高的突变点(图 2)。1957—1986年多年平均年均温为3.3℃, 1987—2014年多年平均年均温为4.1℃, 较突变前升高了0.8℃。1957—1986年多年平均年降水量为822.2 mm, 1987—2014年多年平均年降水量为827.4 mm, 基本保持不变。

|
| 图 2 东岗气象站1957—2014年年均气温Mann-Kendall检验结果 Fig. 2 The results of annual mean temperature by Mann-Kendall test from 1957 to 2014 Ufk正常的统计值时间序列The normal statistical time sequence; Ubk逆序的统计值时间序列The inverted statistical time sequence.图中虚线表示0.05显著性水平 |
本研究通过树木年轮学方法得到3个海拔红松标准年表(图 3), 其特征值及公共区间分析结果见表 1。低海拔(HS1)、中海拔(HS2)和高海拔(HS3)年表的平均敏感度分别为0.191、0.144和0.116, 样本总体代表性分别为0.956、0.950和0.945, 第一主成分解释方差量分别为36.1%、40.6%和33.8%。因此, 年表中包含一定的气候信息, 适用于气候因子的响应分析。

|
| 图 3 不同海拔红松标准年表 Fig. 3 Standardized chronologies of P. koraiensis at different elevations |
| 样点Sample site | HS1 | HS2 | HS3 |
| 标准差Standard deviation | 0.121 | 0.124 | 0.122 |
| 平均敏感度Mean sensitivity | 0.191 | 0.144 | 0.116 |
| 一阶自相关系数First order autocorrelation | 0.280 | 0.492 | 0.253 |
| 树间相关系数Correlation between trees | 0.334 | 0.373 | 0.307 |
| 信噪比Signal-to-noise ratio | 21.528 | 18.871 | 17.302 |
| 样本总体代表性Expressed population signal | 0.956 | 0.950 | 0.945 |
| 第一主成分方差量Variance in first eigenvector/% | 36.1 | 40.6 | 33.8 |
| HS1低海拔红松年表Standardized chronology of P. koraiensis at low elevation; HS2中海拔红松年表Standardized chronology of P. koraiensis at middle elevation; HS3高海拔红松年表Standardized chronology of P. koraiensis at high elevation | |||
气温突变前, 低海拔年表与当年6、7月降水显著正相关, 与前一年9月平均最低气温显著正相关。中海拔年表与当年7月平均最低气温显著正相关, 与当年3月降水显著负相关。高海拔年表与当年7月平均气温、平均最低气温显著正相关, 与前一年9月降水显著正相关(图 4)。气温突变后, 低海拔年表与前一年11月气温因子显著正相关, 与当年6、7月降水正相关减弱。中海拔年表与前一年11月降水显著负相关, 与7月平均最低气温正相关减弱。高海拔年表与7月平均气温、平均最低气温正相关减弱(图 5)。

|
| 图 4 红松标准年表与月气候因子的相关系数(1957—1986年) Fig. 4 Correlation coefficients between standardized chronologies and monthly climate factors(1957—1986) T平均气温Mean temperature; Tmax平均最高气温Maximum mean temperature; Tmin平均最低气温Minimum mean temperature; P降水量Precipitation; p:上一年Past;c:当年Current |

|
| 图 5 红松标准年表(STD)与月气候因子的相关性(1987—2014年) Fig. 5 Correlation coefficients between standardized chronologies and monthly climate factors(1987—2014) |
通过μ值计算检验气温突变前后两个时间段红松标准年表与相同气候因子相关系数的差异情况, 若|μ| >1.96则代表两个时段树轮年表与相同气候因子相关系数有显著差异(P < 0.05)。气温突变前后各海拔年表与月平均气温、月平均最高气温、月平均最低气温的相关关系并没有发生显著改变, 但中海拔年表与前一年11月降水的相关关系发生了显著改变, 高海拔年表与当年5月降水的相关关系发生了显著改变(图 6)。

|
| 图 6 1957—1986年和1987—2014年红松标准年表与相同月气候因子相关系数的差异性检验结果(|μ|) Fig. 6 Test results of the correlation differences between standardized chronologies and climate factors for 1957—1986 and 1987—2014 (|μ|) 方块大小表示|μ|大小Sizes of the diamond indicated the strength of the |μ|; 实心表示0.05显著性水平 |
通过μ值计算检验结果, 对红松标准年表—气候因子关系发生显著变化的中、高海拔进行年表与月降水量和月平均气温的滑动相关分析(滑动窗口为25年)。中海拔年表与当年5、6月降水的负相关转变为正相关, 与当年7月降水的正相关增强, 与当年3月降水量的负相关逐渐减弱, 与前一年11月降水的相关系数由正相关转变为负相关;与当年5—7月平均气温的正相关转变为负相关。高海拔年表与当年7、8月降水的正相关较为稳定, 与5、6月降水的负相关转变为正相关, 与前一年9月降水的正相关逐渐减弱;与当年7月平均气温的正相关逐渐减弱(图 7)。

|
| 图 7 中、高海拔红松标准年表(STD)与月降水量、月平均气温的滑动相关分析 Fig. 7 Moving correlated analysis of standardized chronologies with monthly climate factors at middle and high altitude 图a为中海拔红松标准年表与月平均气温的滑动相关Moving correlated analysis of standardized chronology with monthly average temperature at middle altitude; 图b为中海拔红松标准年表与月降水量的滑动相关Moving correlated analysis of standardized chronology with monthly precipitation at middle altitude; 图c为高海拔红松标准年表与月平均气温的滑动相关Moving correlated analysis of standardized chronology with monthly average temperature at high altitude; 图d为高海拔红松标准年表与月降水量的滑动相关Moving correlated analysis of standardized chronology with monthly precipitation at high altitude; 黑色实心圆圈表示0.01显著性水平; 黑色空心圆圈表示0.05显著性水平 |
分析红松树轮年表与气候因子的相关关系(图 4、图 5), 结果显示低海拔红松径向生长主要受生长季降水限制, 中海拔和高海拔红松径向生长主要受当年7月气温限制, 尤其是低温的限制作用明显。气温突变以后各海拔树轮宽度年表与气候要素的相关关系减弱, 显著相关的月份减少。王辉等通过对红松细胞尺度的树轮指标研究也发现了气温突变以后树轮指标与气候要素相关关系减弱的现象[18]。
低海拔年表与当年6、7月降水正相关, 与前一年9月最低气温正相关。6—7月是红松生长最为旺盛的时期, 此时充足的降水有利于树木光合作用, 生产足够的光合产物, 促进树木生长[11]。前一年9月温暖的气候条件, 有助于树木对碳水化合物的积累, 为来年树木径向生长提供养分[19]。气温突变以后, 年表与前一年11月气温因子显著正相关, 与当年6、7月降水的正相关减弱。前一年11月温度升高有助于树木减少冬芽等的损失, 减少对树木体内同化物的消耗, 有利于树木来年生长[20]。
中海拔年表与当年7月平均最低气温显著正相关, 与当年3月降水显著负相关。7月较高的温度能够增强树木生理活动能力, 产生生长调节剂促进细胞增大, 产生宽轮[21]。春季过多的降水会导致气温下降, 推后树木进入生长季的时间[10]。气温突变以后, 年表与前一年11月降水显著负相关, 与当年7月平均最低气温的正相关减弱。11月过多的降水, 导致树木呼吸作用增强, 使处于休眠状态的树木消耗增加, 不利于营养物质存积[12]。
高海拔年表与当年7月平均气温、平均最低气温显著正相关, 与前一年9月降水显著正相关。高海拔降水量较多, 前一年9月丰富的降水有利于光合产物的积累以及土壤水分涵养, 为来年树木的生长提供充足的水分和良好的条件。这种生长季末期降水的“滞后效应”在青海云杉(Picea crassifolia)等其他针叶树种上也有所体现[22]。气温突变以后, 年表与7月气温因子的正相关减弱。气温的显著升高导致了高海拔红松径向生长对当年7月气温因子的正响应减弱。
3.2 探讨气温突变以后红松径向生长对气候因子响应关系发生显著改变的原因对μ值检验结果进行分析(图 6), 发现气温突变前后低海拔年表与气候因子的相关关系较为稳定, 并未发现显著改变(|μ|<1.96, 在0.05显著水平)。中海拔年表与前一年11月降水的相关关系发生显著改变(|μ|>1.96, 在0.05显著水平), 由正相关转变为负相关。高海拔年表与当年5月降水的相关关系发生显著改变(|μ|>1.96, 在0.05显著水平), 由负相关转变为正相关。低海拔红松径向生长主要受生长季降水限制, 而中海拔和高海拔红松径向生长主要受低温限制。气温显著升高对中、高海拔红松径向生长对气候因子响应关系的影响要强于低海拔。因此, 气温突变背景下, 低海拔红松树轮年表更适用于区域气候重建等研究。但是随着气温的持续升高, 低海拔红松径向生长对气候因子的响应关系是否会发生显著改变, 还需进一步的研究与探讨。
造成树轮年表与气候因子相关关系发生显著改变的原因非常复杂。在本研究中, 高海拔年表与当年5月降水的相关关系发生显著改变, 可以解释为是气温的显著升高引起水分胁迫造成的。气温的显著升高加快土壤水分的蒸散速度, 引起土壤可利用水分减少[23]。通过滑动相关的结果(图 7), 高海拔红松标准年表与当年5、6月降水的负相关转变为正相关, 以及与当年7月平均气温正相关减弱, 说明气温显著升高引起了水分胁迫[24]。焦亮等对北天山东段西伯利亚落叶松(Larix sibirica)的研究发现, 在气温突变以后生长季高温引起的土壤水分亏缺是限制树木径向生长的主要因素[25]。在中海拔地区, 年表与生长季月降水量和月平均气温的滑动相关关系也说明了气温显著升高引起了中海拔区当年生长季的水分胁迫(图 7)。但是中海拔年表与前一年11月降水相关系数由正相关变为负相关的改变, 意味着气温升高引起的水分胁迫不能很好的解释中海拔区红松径向生长对前一年11月降水相关系数的变化。因此, 导致树木径向生长对气候因子响应关系发生显著改变的原因及机制还需进一步的研究与探讨。
3.3 探讨气温突变对不同海拔红松径向生长的影响气温突变后, 长白山地区气温显著升高, 降水量基本保持不变。低海拔红松径向生长主要受生长季降水限制, 气温升高加快了土壤水分蒸散速度, 引起土壤可利用水分减少, 在研究区降水基本保持不变的条件下有可能导致红松径向生长下降[19]。中、高海拔红松径向生长主要受当年7月气温限制, 气温突变以后, 气温的开始升高本可以增强树木光合效率, 促进树木径向生长[26]。但是随着气温的持续升高, 中、高海拔红松径向生长对生长季气温的正相关逐渐减弱, 并有转向负相关的趋势, 这说明原本有利于中、高海拔红松径向生长的气候条件随着气温的持续增加而逐渐转变为不利于树木径向生长。未来研究区的气温持续升高, 中、高海拔红松径向生长有可能由增加趋势转变为下降趋势。
| [1] | IPCC. Climate Change 2013:The Physical Science Basis:Contribution of Working Group Ⅰ to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, 2013. |
| [2] | Linares J C, Tíscar P A. Climate change impacts and vulnerability of the southern populations of Pinus nigra subsp. salzmannii. Tree Physiology, 2010, 30(7): 795–806. DOI:10.1093/treephys/tpq052 |
| [3] | Herrero A, Rigling A, Zamora R. Varying climate sensitivity at the dry distribution edge of Pinus sylvestris and P. nigra. Forest Ecology and Management, 2013, 308: 50–61. DOI:10.1016/j.foreco.2013.07.034 |
| [4] | Vila B, Vennetier M, Ripert C, Chandioux O, Liang E Y, Guibal F, Torre F. Has global change induced divergent trends in radial growth of Pinus sylvestris and Pinus halepensis at their bioclimatic limit? The example of the Sainte-Baume forest (South-east France). Annals of Forest Science, 2008, 65(7): 709–709. DOI:10.1051/forest:2008048 |
| [5] | Briffa K R, Schweingruber F H, Jones P D, Osborn T J, Shiyatov S G, Vaganov E A. Reduced sensitivity of recent tree-growth to temperature at high northern latitudes. Nature, 1998, 391(6668): 678–682. DOI:10.1038/35596 |
| [6] | Franceschini T, Bontemps J D, Leban J M. Transient historical decrease in earlywood and latewood density and unstable sensitivity to summer temperature for Norway spruce in northeastern France. Canadian Journal of Forest Research, 2012, 42(2): 219–226. DOI:10.1139/x11-182 |
| [7] | 胡琦, 潘学标, 张丹, 杨宁, 李秋月, 邵长秀. 东北地区不同时间尺度下气温和无霜期的变化特征. 中国农业气象, 2015, 36(1): l–8. |
| [8] | 邵雪梅, 吴祥定. 利用树轮资料重建长白山区过去气候变化. 第四纪研究, 1997, 17(1): 76–85. |
| [9] | 张寒松, 韩士杰, 李玉文, 张军辉. 利用树木年轮宽度资料重建长白山地区240年来降水量的变化. 生态学杂志, 2007, 26(12): 1924–1929. |
| [10] | 李广起, 白帆, 桑卫国. 长白山红松和鱼鳞云杉在分布上限的径向生长对气候变暖的不同响应. 植物生态学报, 2011, 35(5): 500–511. |
| [11] | 陈列, 高露双, 张赟, 张思行, 赵秀海. 长白山北坡不同林型内红松年表特征及其与气候因子的关系. 生态学报, 2013, 33(4): 1285–1291. |
| [12] | 陈力, 尹云鹤, 赵东升, 苑全治, 吴绍洪. 长白山不同海拔树木生长对气候变化的响应差异. 生态学报, 2014, 34(6): 1568–1574. |
| [13] | 王晓明, 赵秀海, 高露双, 姜庆彪. 长白山北坡不同年龄红松年表及其对气候的响应. 生态学报, 2011, 31(21): 6378–6387. |
| [14] | Holmes R L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin, 1983, 43: 69–78. |
| [15] | 王晓明, 赵秀海, 高露双, 姜庆彪. 长白山北坡沿海拔梯度岳桦径向生长对气候的响应. 应用与环境生物学报, 2013, 19(6): 929–934. |
| [16] | Lindeman R H, Merenda P F, Gold R Z. Introduction to Bivariate and Multivariate Analysis. Glenview Illinois: Scott, Foresman, 1980. |
| [17] | Biondi F, Waikul K. DENDROCLIM2002:A C++ program for statistical calibration of climate signals in tree-ring chronologies. Computers & Geosciences, 2004, 30(3): 303–311. |
| [18] | 王辉, 邵雪梅, 方修琦, 尹志勇, 陈力, 赵东升, 吴绍洪. 长白山红松年轮细胞尺度参数对气候要素的响应. 应用生态学报, 2011, 22(10): 2643–2652. |
| [19] | Yu D P, Wang Q W, Wang Y, Zhou W M, Ding H, Fang X M, Jiang S W, Dai L M. Climatic effects on radial growth of major tree species on Changbai Mountain. Annals of Forest Science, 2011, 68: 921. DOI:10.1007/s13595-011-0098-7 |
| [20] | 于大炮, 王顺忠, 唐立娜, 代力民, 王庆礼, 王绍先. 长白山北坡落叶松年轮年表及其与气候变化的关系. 应用生态学报, 2005, 16(1): 14–20. |
| [21] | 陈力, 吴绍洪, 戴尔阜. 长白山红松和落叶松树轮宽度年表特征. 地理研究, 2011, 30(6): 1147–1155. |
| [22] | 彭剑峰, 勾晓华, 陈发虎, 刘普幸, 张永, 方克艳. 阿尼玛卿山地不同海拔青海云杉(Picea crassifolia)树轮生长特性及其对气候的响应. 生态学报, 2007, 27(8): 3268–3276. |
| [23] | Deslauriers A, Beaulieu M, Balducci L, Giovannelli A, Gagnon M J, Rossi S. Impact of warming and drought on carbon balance related to wood formation in black spruce. Annals of Botany, 2014, 114(2): 335–345. DOI:10.1093/aob/mcu111 |
| [24] | Wang H, Shao X M, Jiang Y, Fang X Q, Wu S H. The impacts of climate change on the radial growth of Pinus koraiensis along elevations of Changbai Mountain in northeastern China. Forest Ecology and Management, 2013, 289: 333–340. DOI:10.1016/j.foreco.2012.10.023 |
| [25] | Jiao L, Jiang Y, Zhang W T, Wang M C, Zhang L N, Zhao S D. Divergent responses to climate factors in the radial growth of Larix sibirica in the eastern Tianshan Mountains, northwest China. Trees, 2015, 29(6): 1673–1686. DOI:10.1007/s00468-015-1248-6 |
| [26] | Rossi S, Girard M J, Morin H. Lengthening of the duration of xylogenesis engenders disproportionate increases in xylem production. Global Change Biology, 2014, 20(7): 2261–2271. DOI:10.1111/gcb.2014.20.issue-7 |