 2017, Vol. 37
2017, Vol. 37文章信息
- 邱廉, 陶婷婷, 韩善锐, 杨文宇, 栾夏丽, 邱燕宁, 刘茂松, 徐驰.
- QIU Lian, TAO Tingting, HAN Shanrui, YANG Wenyu, LUAN Xiali, QIU Yanning, LIU Maosong, XU Chi.
- 宏生态尺度上景观破碎化对物种丰富度的影响
- Effects of local landscape fragmentation on species richness at a macroecological scale
- 生态学报. 2017, 37(22): 7595-7603
- Acta Ecologica Sinica. 2017, 37(22): 7595-7603
- http://dx.doi.org/10.5846/stxb201609131855
-
文章历史
- 收稿日期: 2016-09-13
- 网络出版日期: 2017-07-12

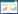




越来越多的研究表明, 人类活动已成为第六次全球大规模物种灭绝的主要驱动因素[1]。导致生物多样性丧失的主要因素中大部分与人类活动直接相关, 包括气候变化、生境减少和破碎化、环境污染、外来物种入侵等[2]。其中, 由土地利用和自然资源开采等人类活动导致原本连续的生境趋于破碎化是威胁生物多样性的关键因素之一[3-4]。
生境破碎化的后果体现在诸多方面, 例如生境斑块面积减小, 生境斑块数量增加, 斑块形状趋于不规则, 核心生境面积缩小, 生境隔离程度增加等[4]。这些景观结构的改变可能对一系列重要的生态过程产生显著影响, 例如在生境斑块的隔离效应的作用下, 生物种群的迁移和定殖过程受到限制, 生境斑块所支持的集合种群规模变小从而限制种群的基因交流[5]。生境破碎化的多种负面效应使得景观内物种存续能力下降, 最终导致整个系统的物种多样性减小[2]。景观破碎化对物种多样性的影响已经成为当前保护生物学研究的重要课题。其中大量研究表明, 在景观尺度上(空间幅度通常从几公里到数百公里)的破碎化对物种多样性具有显著的负面影响, 然而在更大的宏生态学尺度上(空间幅度为全国、大陆至全球), 破碎化的生态效应仍不明确。相关研究发现, 在宏观尺度上, 景观格局因子在整体上对物种丰富度格局具有显著的解释能力[6-9], 但同时也有研究表明这种关系相对较弱[10]。总体上, 景观破碎化对物种多样性宏生态格局的影响尚有待深入研究。
为揭示景观破碎化在物种多样性宏生态格局的形成中的作用, 本文以我国大陆为研究区域, 利用景观指数作为度量景观破碎化的变量, 研究了景观破碎化对鸟类及哺乳动物物种丰富度的影响, 以期深入理解物种多样性地理格局的形成机制, 并为生物多样性保护实践提供参考。
1 研究方法 1.1 研究区概况本研究的研究区域为中国大陆(纬度范围约35°, 经度范围约60°, 海拔跨度近7 km)。我国具有复杂多样的气候和地形, 也是世界上生物多样性最丰富的国家之一。由于中国在末次盛冰川期中受到冰川气候的影响较小, 适合进行环境与物种丰富度之间的关联关系检验[11]。
1.2 数据来源及预处理 1.2.1 物种丰富度数据本研究所使用的物种丰富度数据(包括鸟类和哺乳动物)来自于中国物种信息服务(CSIS)数据库, 其数据库建立于1996年并更新至今, 包含575种哺乳动物、1328种鸟类[12]。物种多样性数据最初是以县级行政单元为单位进行统计, 为了消除空间尺度或面积对物种分布格局的影响, 本研究将其转换为100 km×100 km的网格, 并剔除研究区域边缘土地面积不足网格面积50%的网格(图 1)。

|
| 图 1 基于100 km×100 km网格的物种丰富度和景观水平破碎化变量空间分布 Fig. 1 Spatial distributions of species richness and landscape-level fragmentation variables in mainland China per 100 km×100 km grid cell a:哺乳动物, Mammals; b:鸟类, Birds; c:平均邻近指数, MPI; d:平均欧式最近邻接距离, MNN; e:斑块边缘密度, ED; f:斑块数量, NP |
气候和生境异质性是生物多样性地理格局的主要影响因素[13-14]。本文选用空间分辨率为1 km的19个气候变量和20个生境异质性变量来综合表征这两种主要因素。年均气候数据(1960—1990年)及高程数据下载自WorldClim网站(http://www.worldclim.org)[15], 生境异质性数据(2001—2005年)[16]下载自http://www.earthenv.org。土地利用/覆盖数据(1 km分辨率, 2000年)下载自Global Land Cover Facility(GLCF, http://www.landcover.org)。在100 km×100 km网格内分别计算各环境变量(表 1)。本文采用4种景观指数来量度景观破碎化(表 1、表 2)。计算景观指数前, 将上述1 km分辨率的GLCF土地利用/覆盖数据(包含24个类别)重新分成农田、林地、草地、裸地和水体5类[8]。基于重分类的土地覆盖数据计算网格中的景观指数(图 1), 其中包括生境类型水平(对应单个土地覆盖类型)和景观水平。空间数据在ArcGIS 10.1中进行处理, 景观指数通过Patch Analyst 5.2[17]进行计算。
| 气候变量 Climate | 生境异质性变量 Habitat heterogeneity | 景观破碎化变量 Landscape fragmentation |
| 年平均气温 | 增强型植被指数相关系数 | 景观水平-平均邻近指数 |
| 昼夜温差幅度 | 增强型植被指数幅度 | 景观水平-平均欧式最近邻接距离 |
| 等温性 | 增强型植被指数香农多样性指数 | 景观水平-斑块数量 |
| 温度季节性 | 增强型植被指数辛普森多样性指数 | 景观水平-斑块边缘密度 |
| 最热月份的最高气温 | 增强型植被指数标准误差 | 农田-平均邻近指数 |
| 最冷月份的最低气温 | 增强型植被指数均一性 | 农田-平均欧式最近邻接距离 |
| 年温差幅度 | 增强型植被指数方差 | 农田-斑块数量 |
| 最湿季度的平均气温 | 增强型植被指数对比度 | 农田-斑块边缘密度 |
| 最干季度的平均气温 | 增强型植被指数均匀度 | 森林-平均邻近指数 |
| 最热季度的平均气温 | 增强型植被指数熵 | 森林-平均欧式最近邻接距离 |
| 最冷季度的平均气温 | 增强型植被指数相异度 | 森林-斑块数量 |
| 年平均降水 | 增强型植被指数变异系数 | 森林-斑块边缘密度 |
| 湿度最大月份的降水量 | 增强型植被指数同质性 | 草地-平均邻近指数 |
| 湿度最小月份的降水量 | 增强型植被指数最大值 | 草地-平均欧式最近邻接距离 |
| 降水季节性 | 高程差 | 草地-斑块数量 |
| 湿度最大季度的降水量 | 高程标准误差 | 草地-斑块边缘密度 |
| 湿度最小季度的降水量 | 高程变异系数 | 裸地-平均邻近指数 |
| 最热季度的降水量 | 辛普森景观多样性指数 | 裸地-平均欧式最近邻接距离 |
| 最冷季度的降水量 | 香农景观多样性指数 | 裸地-斑块数量 |
| 景观类型丰富度 | 裸地-斑块边缘密度 | |
| 水体-平均邻近指数 | ||
| 水体-平均欧式最近邻接距离 | ||
| 水体-斑块数量 | ||
| 水体-斑块边缘密度 |
| 景观指数 Landscape metrics | 景观指数计算公式 Calculation formulas of landscape metrics |
| 斑块数量(NP) Number of patches | NP=Ni Ni为景观类型i的斑块数。 |
| 平均欧式最近邻接距离(MNN) Mean Euclidean nearest-neighbor index |  hij为从斑块ij到同类型的斑块的最近距离之和, N为具有最近距离的斑块总数。 |
| 平均邻近指数(MPI) Mean proximity index | 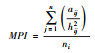 ni为景观类型i的斑块数, aij为景观类型i斑块面积, hij2为某斑块到同类型斑块的最近距离。 |
| 斑块边缘密度(ED) Edge density | 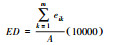 eik为景观类型i的斑块边缘总长度, A为景观总面积。 |
气候变量与生境异质性变量的意义及计算公式详见参考文献[15-16]。
1.3 统计分析为了量化气候、生境异质性和景观破碎化的作用, 本研究将解释变量分为3个相对应的变量集分别进行模型拟合(分别对应于气候模型、生境异质性模型和景观破碎化模型)。考虑到解释变量之间存在多重共线性的影响, 采用主成分分析来消除解释变量之间的多重共线性, 即分别将3组原始的解释变量进行主成分分析, 生成的相互正交的主成分变量。由于本研究关注景观破碎化因子的解释能力, 进行回归模型分析时保留了全部主成分变量以减少自变量信息的丧失。
回归模型中变量的空间自相关可能导致参数估计偏差, 增加I型误差的概率[18]。本研究使用Moran′s I系数来检测模型残差的空间自相关水平。Moran′s I指数的值域为[-1, 1], 值趋于1表明总体上空间正相关程度较高, 性质相似的单元分布较为集中;指数的值趋于-1表明空间负相关程度较高, 总体上邻近单元间差异较大, 高值与低值单元邻近, 性质相似单元趋于离散分布;Moran′s I指数的值接近0则表明总体空间自相关程度较低, 空间单元属性值均接近0或平均值, 或者总体上趋向于随机分布的格局[18]。
本研究分别采用最小二乘(OLS)模型和空间自回归(simultaneous autoregressive, SAR)模型(本文假设空间自相关存在于误差项中, 因此选择SAR error model)进行关联分析[19]。采用AIC与伪决定系数(pseudo R2)来比较不同模型的拟合优度[20]。进一步利用方差分解技术比较3组变量的相对重要性[8]。为确保数据的正态性, 模型拟合之前对物种丰富度数据进行对数变换。统计分析在R 3.3.1软件中进行, SAR模型分析用spdep包完成[21]。
2 结果相关分析结果表明(表 3), 鸟类和哺乳动物的物种丰富度对景观破碎化变量的响应表现出相同的方向性, 但不同景观破碎化变量的效应具有明显差别。考虑到景观变量之间的多重共线性, 本文利用主成分回归重点考察破碎化因子作为整体(而非单个因子)与物种丰富度的关联强度。普通最小二乘法(OLS)模型与空间自回归(SAR)模型的Moran′s I残差曲线图(图 2)表明在100 km尺度上, 空间自回归模型的残差稳定在0附近, 表明空间自回归模型对于解决空间自相关问题有较好的效果。SAR模型具有更高的拟合优度和更小的AIC值。为了更好地区分各组变量的作用的相对强弱, 通过OLS模型调整后的拟合优度和SAR模型非空间部分的伪决定系数来进行判定。对于鸟类和哺乳动物来说, 不考虑空间自相关时(OLS模型和SAR模型中的非空间部分), 景观破碎化模型的拟合优度都略低于气候模型与生境异质性模型, 其解释率达到约30%—50%;当考虑空间自相关时(SAR模型), 景观破碎化模型具有最高的拟合优度, 其解释率达到约60%—80%(表 4)。统计结果总体上表明景观破碎化对于鸟类和哺乳动物多样性的地理格局具有重要影响。
| 气候 Climate | 相关系数 Pearson correlation coefficients | 生境异质性 Habitat heterogeneity | 相关系数 Pearson correlation coefficients | 景观破碎化 Landscape fragmentation | 相关系数 Pearson correlation coefficients | |||
| 哺乳动物 Mammals | 鸟类 Birds | 哺乳动物 Mammals | 鸟类 Birds | 哺乳动物 Mammals | 鸟类 Birds | |||
| 年平均气温 | 0.503*** | 0.364*** | 增强型植被指数相关系数 | -0.330*** | -0.117*** | 景观水平-平均邻近指数 | -0.372*** | -0.291*** |
| 昼夜温差幅度 | -0.536*** | -0.365*** | 增强型植被指数幅度 | 0.048 | 0.128*** | 景观水平-平均欧式最近邻接距离 | -0.053 | -0.035 |
| 等温性 | -0.083* | 0.042 | 增强型植被指数香农多样性指数 | 0.435*** | 0.343*** | 景观水平-斑块数量 | 0.526*** | 0.353*** |
| 温度季节性 | -0.268*** | -0.261*** | 增强型植被指数辛普森多样性指数 | 0.252*** | 0.235*** | 景观水平-斑块边缘密度 | 0.309*** | 0.234*** |
| 最热月份的最 高气温 | 0.315*** | 0.178*** | 增强型植被指数标准误差 | 0.011 | 0.097** | 农田-平均邻近指数 | 0.122*** | 0.064 |
| 最冷月份的最 低气温 | 0.501*** | 0.375*** | 增强型植被指数均一性 | -0.249*** | -0.074* | 农田-平均欧式最近邻接距离 | -0.144*** | -0.120*** |
| 年温差幅度 | -0.365*** | -0.316*** | 增强型植被指数方差 | 0.244*** | 0.274 | 农田-斑块数量 | 0.404*** | 0.241*** |
| 最湿季度的平 均气温 | 0.361*** | 0.236*** | 增强型植被指数对比度 | 0.250*** | 0.273*** | 农田-斑块边缘密度 | 0.518*** | 0.382*** |
| 最干季度的平 均气温 | 0.438*** | 0.349*** | 增强型植被指数均匀度 | 0.185*** | 0.184*** | 森林-平均邻近指数 | 0.334*** | 0.240*** |
| 最热季度的平 均气温 | 0.393*** | 0.244*** | 增强型植被指数熵 | 0.470*** | 0.338*** | 森林-平均欧式最近邻接距离 | -0.119*** | -0.071* |
| 最冷季度的平 均气温 | 0.492*** | 0.381*** | 增强型植被指数相异度 | 0.404*** | 0.339*** | 森林-斑块数量 | 0.426*** | 0.300*** |
| 年平均降水 | 0.548*** | 0.374*** | 增强型植被指数变异系数 | -0.053 | 0.062* | 森林-斑块边缘密度 | 0.560*** | 0.453*** |
| 湿度最大月份的 降水量 | 0.526*** | 0.384*** | 增强型植被指数同质性 | -0.287*** | -0.128*** | 草地-平均邻近指数 | -0.227*** | -0.248*** |
| 湿度最小月份的 降水量 | 0.429*** | 0.222*** | 增强型植被指数最大值 | -0.273*** | -0.097** | 草地-平均欧式最近邻接距离 | -0.033 | -0.018 |
| 降水季节性 | -0.451*** | -0.294*** | 高程差 | 0.163*** | 0.259*** | 草地-斑块数量 | 0.512*** | 0.385*** |
| 湿度最大季度的 降水量 | 0.542*** | 0.401*** | 高程标准误差 | 0.142*** | 0.236*** | 草地-斑块边缘密度 | 0.018 | -0.003 |
| 湿度最小季度的 降水量 | 0.414*** | 0.211*** | 高程变异系数 | 0.314*** | 0.294*** | 裸地-平均邻近指数 | -0.365*** | -0.275*** |
| 最热季度的降 水量 | 0.536*** | 0.414*** | 辛普森景观多样性指数 | 0.396*** | 0.316*** | 裸地-平均欧式最近邻接距离 | 0.152*** | 0.048 |
| 最冷季度的降 水量 | 0.395*** | 0.195*** | 香农景观多样性指数 | 0.338*** | 0.284*** | 裸地-斑块数量 | -0.287*** | -0.259*** |
| 景观类型丰富度 | 0.296*** | 0.207*** | 裸地-斑块边缘密度 | -0.472*** | -0.375*** | |||
| 水体-平均邻近指数 | 0.004 | 0.017 | ||||||
| 水体-平均欧式最近邻接距离 | -0.011 | 0.005 | ||||||
| 水体-斑块数量 | 0.169*** | 0.125*** | ||||||
| 水体-斑块边缘密度 | 0.088** | 0.090** | ||||||
| *** P < 0.001, ** P < 0.01, * P < 0.05 | ||||||||
| 模型 Models | 哺乳动物Mammals | 鸟类Birds | |||||||||||
| OLS | SAR | OLS | SAR | ||||||||||
| R2 | AIC | Pseudo R2 | 非空间部分 pseudo R2 | AIC | R2 | AIC | Pseudo R2 | 非空间部分 pseudo R2 | AIC | ||||
| 气候Climate | 0.520 | 2145 | 0.793 | 0.412 | 1584 | 0.417 | 2469 | 0.583 | 0.395 | 2271 | |||
| 生境异质性 Habitat heterogeneity | 0.485 | 2209 | 0.804 | 0.421 | 1547 | 0.327 | 2600 | 0.607 | 0.279 | 2259 | |||
| 景观破碎化 Landscape fragmentation | 0.476 | 2229 | 0.804 | 0.413 | 1575 | 0.300 | 2639 | 0.616 | 0.238 | 2255 | |||

|
| 图 2 SAR模型和OLS模型的残差Moran′s I系数 Fig. 2 Moran′s I coefficients of the residuals of the SAR models and OLS models |
方差分解的结果显示, 单纯由景观破碎化因子解释的物种丰富度变异都较小, 分别为2.6%(哺乳类)和5.7%(鸟类)(图 3), 但是与单纯的气候与生境异质性因子的解释率相比较而言, 并不能完全忽视。而且景观破碎化与其他两类环境变量有着较高的交叉解释率, 对于哺乳动物, 交叉解释率达到了38.7%, 而对于鸟类, 其交叉解释率也达到了18.1%, 明显高于景观破碎化变量的单独解释率。

|
| 图 3 基于SAR模型非空间部分的方差分解 Fig. 3 Variation partitioning based on the non-spatial trends of the simultaneous autoregressive (SAR) models A:气候因子, Climate; B:生境异质性因子, Habitat heterogeneity; C:景观破碎化因子, Landscape fragmentation |
生物多样性的地理格局及其形成机制是宏生态学与生物地理学的研究主题之一[14, 22]。过去几十年来, 大量研究关注于理解生物多样性地理格局的驱动因子, 并由此产生了大量理论和假说(如能量假说、水热假说、生境异质性假说、进化历史假说等[20])。随着全球尺度环境数据的普及和精度的提高, 研究人员开始关注气候和地形等主导因素以外可能发生作用但在以往研究中被忽视的驱动因素[23]。对于宏观生态格局, 研究者往往先验地假设这些相对较小尺度的过程不会对更大尺度的格局产生影响, 然而最近一系列相关研究发现该假设在很大程度上存在不确定性[24]。例如传统研究通常认为生物间相互作用(如竞争、捕食等)只可能影响群落尺度上的生态格局和过程, 然而最新的研究表明这种群落尺度的相互作用可能产生跨等级的作用, 从而影响到大陆尺度上的生物多样性格局[25]。涉及到生境破碎化的效应, 虽然研究者早已认识到该因素在景观尺度对物种多样性分布具有重要影响, 但一直以来并未注意到其在宏观尺度上也可能发挥显著作用。近年来越来越多的证据表明, 景观格局因子同样也可能产生跨尺度作用, 在国家乃至大陆尺度上的物种分布与多样性的地理格局形成中扮演重要角色[6-7]。
研究结果进一步证实, 景观破碎化对中国大陆的鸟类和哺乳动物物种丰富度地理格局的确具有明显的关联关系。基于多元回归模型单独观察景观破碎化变量的效应, 可发现其对物种丰富度的方差贡献率最高可达近50%(基于非空间模型)或80%(基于空间模型), 仅略低于气候和生境异质性的效应。方差分解的结果表明景观破碎化的单独解释率虽然较小, 但并不足以完全忽视其效应;而且其很大程度上与气候和生境异质性变量的贡献率相互重叠, 这与Reino等[6]的研究结果一致。由于变量之间存在的多重共线性, 本文仅依靠统计分析无法对重叠部分中景观破碎化的效应进行分解, 有待于进一步开展深入研究以阐释景观破碎化相较于其他宏观环境变量的相对重要性。
景观破碎化变量对于哺乳动物物种丰富度格局的总体解释率高于鸟类, 这可能归因于其生活史特征的差异。鸟类由于具备飞翔能力, 其生活史中的各项活动(例如迁徙、觅食等)通常发生在相对更大的空间尺度上[26], 相对哺乳动物而言具有相对更强的扩散能力[27], 而表现出对景观破碎化相对较低的敏感程度;相反, 地表生境的破碎化使得哺乳动物更易受到生境隔离等效应的负面影响, 从而表现出与破碎化因子更强的关联程度。
研究在对原始解释变量进行主成分分析的基础上进行统计建模, 这可以有效解决解释变量之间多重共线性的影响;另外由于单个景观变量的效应通常较弱[6-7], 这种方法也可以避免逐步回归分析中由于变量筛选导致的无法对景观破碎化变量的整体效应进行评估的问题。当前关于景观破碎化对物种丰富度影响的研究主要关注于其负面效应, 本研究中的单变量分析结果的确发现, 在宏观水平上诸多景观破碎化变量与鸟类和哺乳动物的物种丰富度具有显著的负相关关系;但同时也有相当比例的破碎化变量(例如景观水平上的斑块数量和斑块边缘密度)与物种丰富度呈正相关关系。由于景观格局对种群大小与物种多样性维持同时存在多种(正面与负面)复杂效应, 景观格局指数间的共线性可能导致其他格局因子的效应掩盖破碎化的负面效应, 分离不同景观格局因子的混合效应还有待于后续深入研究。另外考虑到生物种群对生境破碎化有时以非线性的方式进行响应, 提出与生态过程相符的研究假说并建立捕捉非线性关系的合适统计模型(如广义加性模型等)有助于深入理解生境破碎化对生物多样性的效应。
| [1] | Ceballos G, Ehrlich P R, Barnosky A D, García A, Pringle R M, Palmer T M. Accelerated modern human-induced species losses:entering the sixth mass extinction. Science Advances, 2015, 1(5): e1400253. DOI:10.1126/sciadv.1400253 |
| [2] | 武晶, 刘志民. 生境破碎化对生物多样性的影响研究综述. 生态学杂志, 2014, 33(7): 1946–1952. |
| [3] | Fahrig L. Effects of habitat fragmentation on biodiversity. Annual Review of Ecology, Evolution, and Systematics, 2003, 34(1): 487–515. DOI:10.1146/annurev.ecolsys.34.011802.132419 |
| [4] | Fischer J, Lindenmayer D B. Landscape modification and habitat fragmentation:a synthesis. Global Ecology and Biogeography, 2007, 16(3): 265–280. DOI:10.1111/geb.2007.16.issue-3 |
| [5] | Keller I, Largiadèr C R. Recent habitat fragmentation caused by major roads leads to reduction of gene flow and loss of genetic variability in ground beetles. Proceedings of the Royal Society B:Biological Sciences, 2003, 270(1513): 417–423. DOI:10.1098/rspb.2002.2247 |
| [6] | Reino L, Beja P, Araújo M B, Dray S, Segurado P. Does local habitat fragmentation affect large-scale distributions? The case of a specialist grassland bird. Diversity and Distributions, 2013, 19(4): 423–432. DOI:10.1111/ddi.2013.19.issue-4 |
| [7] | Xu C, Huang Z Y X, Chi T, Chen B J W, Zhang M J, Liu M S. Can local landscape attributes explain species richness patterns at macroecological scales?. Global Ecology and Biogeography, 2014, 23(4): 436–445. DOI:10.1111/geb.2014.23.issue-4 |
| [8] | Martins I S, Proença V, Pereira H M. The unusual suspect:land use is a key predictor of biodiversity patterns in the Iberian Peninsula. Acta Oecologica, 2014, 61: 41–50. DOI:10.1016/j.actao.2014.10.005 |
| [9] | Tsianou M A, Koutsias N, Mazaris A D, Kallimanis A S. Climate and landscape explain richness patterns depending on the type of species' distribution data. Acta Oecologica, 2016, 74: 19–27. DOI:10.1016/j.actao.2016.06.006 |
| [10] | Triviño M, Thuiller W, Cabeza M, Hickler T, Araújo M B. The contribution of vegetation and landscape configuration for predicting environmental change impacts on Iberian birds. PLoS One, 2011, 6(12): e29373. DOI:10.1371/journal.pone.0029373 |
| [11] | Qian H, Wang X H, Wang S L, Li Y L. Environmental determinants of amphibian and reptile species richness in China. Ecography, 2007, 30(4): 471–482. DOI:10.1111/j.0906-7590.2007.05025.x |
| [12] | 解焱, 张爽, 王伟. 中国生物多样性地理图集. 长沙: 湖南教育出版社, 2009: 2–7. |
| [13] | Fløjgaard C, Normand S, Skov F, Svenning J C. Deconstructing the mammal species richness pattern in Europe-towards an understanding of the relative importance of climate, biogeographic history, habitat heterogeneity and humans. Global Ecology and Biogeography, 2011, 20(2): 218–230. DOI:10.1111/j.1466-8238.2010.00604.x |
| [14] | Chen S B, Jiang G M, Zhang J L, Li Y G, Qian H. Species turnover of amphibians and reptiles in eastern China:disentangling the relative effects of geographic distance and environmental difference. Ecological Research, 2011, 26(5): 949–956. DOI:10.1007/s11284-011-0850-3 |
| [15] | Hijmans R J, Cameron S E, Parra J L, Jones P G, Jarvis A. Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology, 2005, 25(15): 1965–1978. DOI:10.1002/(ISSN)1097-0088 |
| [16] | Tuanmu M N, Jetz W. A global 1-km consensus land-cover product for biodiversity and ecosystem modelling. Global Ecology and Biogeography, 2014, 23(9): 1031–1045. DOI:10.1111/geb.2014.23.issue-9 |
| [17] | Rempel R S, Kaukinen D, Carr A P. Patch analyst and patch grid. Ontario ministry of natural resources. Thunder Bay, Ontario:Centre for Northern Forest Ecosystem Research. (2015-11-13)[2016-08-18]. http://www.cnfer.on.ca/SEP/patchanalyst/Patch5_2_Install.htm. |
| [18] | 谢花林, 刘黎明, 李波, 张新时. 土地利用变化的多尺度空间自相关分析——以内蒙古翁牛特旗为例. 地理学报, 2006, 61(4): 389–400. DOI:10.11821/xb200604006 |
| [19] | Beale C M, Lennon J J, Yearsley J M, Brewer M J, Elston D A. Regression analysis of spatial data. Ecology Letters, 2010, 13(2): 246–264. DOI:10.1111/ele.2010.13.issue-2 |
| [20] | Hawkins B A, Field R, Cornell H V, Currie D J, Guégan J F, Kaufman D M, Kerr J T, Mittelbach G G, Oberdorff T, O'Brien E M, Porter E E, Turner J R G. Energy, water, and broad-scale geographic patterns of species richness. Ecology, 2003, 84(12): 3105–3117. DOI:10.1890/03-8006 |
| [21] | Bivand R. Spdep:spatial dependence:weighting schemes, statistics and models. (2016-07-30)[2016-08-18]. https://cran.r-project.org/web/packages/spdep/. |
| [22] | 胡慧建, 蒋志刚, 王祖望. 宏生态学(Macroecology)及其研究. 生态学报, 2003, 23(6): 1192–1199. |
| [23] | 林鑫, 王志恒, 唐志尧, 赵淑清, 方精云. 中国陆栖哺乳动物物种丰富度的地理格局及其与环境因子的关系. 生物多样性, 2009, 17(6): 652–663. |
| [24] | 蔡波, 黄勇, 陈跃英, 胡军华, 郭宪光, 王跃招. 中国蛇类物种丰富度地理格局及其与生态因子的关系. 动物学研究, 2012, 33(4): 343–353. |
| [25] | Dehling D M, Fritz S A, Töpfer T, Päckert M, Estler P, Böhning-Gaese K, Schleuning M. Functional and phylogenetic diversity and assemblage structure of frugivorous birds along an elevational gradient in the tropical Andes. Ecography, 2014, 37(11): 1047–1055. |
| [26] | Walters J R. The ecological basis of avian sensitivity to habitat fragmentation//Marzluff J M, Sallabanks R, eds. Avian Conservation:Research and Management. Washington DC:Island Press, 1998:181-192. |
| [27] | 丁晶晶, 刘定震, 李春旺, 蒋志刚. 中国大陆鸟类和兽类物种多样性的空间变异. 生态学报, 2012, 32(2): 343–350. |