 2017, Vol. 37
2017, Vol. 37文章信息
- 王蕾, 张怡婷, 张圣新, 范德玲, 汪贞, 石利利, 刘济宁
- WANG Lei, ZHANG Yiting, ZHANG Shengxin, FAN Deling, WANG Zhen, SHI Lili, LIU Jining.
- 环境条件对伸展摇蚊实验种群繁育的影响
- Effects of environmental conditions on the reproduction and development of Chironomus tentans(Diptera)
- 生态学报. 2017, 37(21): 7152-7160
- Acta Ecologica Sinica. 2017, 37(21): 7152-7160
- http://dx.doi.org/10.5846/stxb201608101639
-
文章历史
- 收稿日期: 2016-08-10
- 网络出版日期: 2017-07-11





环境生物的生存繁育受到温度、光照、湿度等多种环境因子的影响。而对于摇蚊这类经历卵—幼虫—蛹—成虫多次变态发育、生存空间在水—沉积物层、上层水体和水面转换的昆虫而言, 温度和光照是影响其全生命周期的重要环境因素, 且成虫羽化后还会受到空气湿度的影响[1]。摇蚊的世代周期短, 一代摇蚊慢性毒性试验一般为20—65 d, 比选用鱼类作试验生物节省时间和经费, 用其作为模式生物进行生态毒理学研究具有重要的科学意义。
根据已有研究, 不同摇蚊物种对环境温度、光照等条件适应性不同, 每一代的羽化时间20—65 d不等, 种群发展规律有差异[2]。其中温度和光照对摇蚊生长繁殖影响最为显著[1]。因此虽然不同摇蚊种的繁育条件对本土种的研究有一定参考价值, 但为了建立稳定的室内繁育技术和毒性测试技术, 探索区域典型摇蚊种生理特性、研究其繁殖发育条件和种群变化规律仍十分必要。鉴于摇蚊在水生态毒性测试中的诸多优势, 且物种地理分布的差异性对化学物质的生态危害评估结果有重要影响[3], 而影响地域分布差异的主要因素包括温度、光照强度和湿度, 所以虽然OECD已建立了C. riparius、C. yoshimatsui、C. dilutus等摇蚊种的急慢性毒性试验方法[4-6], 但是关于OECD提供的温度和光照等外部条件并不一定是用于本土物种。美国EPA和OECD等发达国家或地区的化学物质生态危害评估导则也明确要求充分考虑本土物种的生态危害性[7-8]。因此建立关于本土生物稳定的室内繁育技术是构建其毒性测试方法的基础。
伸展摇蚊(Chironomus tentans)广泛分布于我国新疆, 青海, 西藏, 甘肃, 内蒙古, 山西, 陕西, 河南, 安徽, 河北, 山东, 江苏等地, 是我国常见的摇蚊种之一[9]。由于缺少伸展摇蚊繁殖发育条件的系统研究, 因此无法构建相应的毒性测试方法, 特别是全生命周期毒性测试方法和多代繁殖毒性测试方法。所以为了评估有毒有害化学物质对我国本土生物的危害性、建立我国广布摇蚊种——伸展摇蚊的室内繁育技术和急慢性毒性及全生命周期毒性测试方法, 系统研究温度、光照、湿度等多种环境因子对伸展摇蚊生存繁育影响显得非常必要。
本研究以伸展摇蚊为研究对象, 重点研究温度、光照条件对其发育率、存活率、性别比和种群动态的影响, 在此基础上提出伸展摇蚊室内繁育的标准化条件及相应的种群发展规律, 为构建本土摇蚊种的毒性测试方法奠定基础。
1 材料与方法 1.1 试验生物伸展摇蚊(Chironomus tentans)取自上海化工研究院, 已在本实验室驯养几十代。驯养条件参考Chironomus riparius的养殖条件[6]。将摇蚊幼虫放入18 L玻璃缸中, 缸中加入与试验基质相同的水—沉积物系统, 体积不超过缸容积的一半;缸口用纱网罩盖, 温和曝气;每日为摇蚊幼虫饲喂粉末状鱼食, 每隔3 d用虹吸法更换一次上覆水, 更换时避免带起缸中的幼虫及其所筑的巢。以此建立的实验种群作为供试虫源。
1.2 试验基质卵置于Elendt M7溶液中孵化, 幼虫和蛹选用Elendt M7溶液和人工沉积物组成的水—沉积物系统作为培养基质。其中Elendt M7溶液的制备方法参考OECD化学品测试导则TG233[6]。人工沉积物的配方包括5%(干重)泥炭(pH值5.9;粉末状, 磨细(颗粒大小≤1 mm)且风干)、20%(干重)高岭土、75%(干重)石英砂(粒径在50—200 μm的细砂占60%)和50%水。
具体配制方法为:首先将泥炭在烘箱内烘干后用粉碎机磨成细粉, 在使用旋转搅拌器用去离子水制备所需的泥炭悬浮液, 悬浮液的pH值为5.9;然后将悬浮液在(20±2) ℃条件下温和搅拌2 d进行驯化以稳定pH值并建立稳定的微生物群。驯化后的泥炭悬浮液pH值为6.0±0.5。最后将泥炭悬浮液与Elendt M7溶液混合, 得到均一的水—沉积物系统。采用1 L烧杯盛放水—沉积物系统, 并确保水层和沉积物层体积比为4:1。使用人工配方沉积物具有以下优势:(1)构建了可重复的“标准化基质”, 无需再找未污染和清洁沉积物, 降低实验结果的变异性;(2)不受沉积物季节性变动的影响, 且不需预处理沉积物以去除本土动物群系。
1.3 试验方法将湿度和光周期分别设置为85%和16:8(光:暗), 设置4个温度(15、25、30、35 ℃)和两种光照强度(800 lx、2000 lx), 考察温度和光照强度对伸展摇蚊繁育的影响。试验在人工气候箱(MMM Climacell)中开展, 同一测试条件的测试容器随机放置。将Elendt M7溶液置于培养皿中在上述环境条件下恒温2 h后, 投入不足24 h的新卵开始试验。每种测试条件下放置3个培养皿, 每皿投入1个卵串。试验开始时每12 h在S8APO体式显微镜(德国Leica)下统计每个卵串的孵化情况(每个卵串中大概200—350粒卵)。待卵孵化为一龄幼虫后, 在同一时刻随机选取100条一龄幼虫投入盛有水—沉积物系统并已事先恒温的烧杯中, 同时对上覆水进行轻微曝气并用纱网封口。每日喂食磨细的鱼食, 每3 d换一次水, 每个处理设置3个重复。试验刚开始10 d内日喂食量控制在0.25—0.5 mg/只, 后期幼虫长大后将日饲喂量增至0.5—1 mg/只。每天在体式显微镜底下观察摇蚊的发育情况, 记录每种形态(卵、幼虫、蛹、成虫)的发育时间(发育历期即发育时间, 发育速率即发育时间的倒数), 死亡数和羽化数。并将所有羽化的摇蚊取出并置于新的盛有Elendt M7溶液的烧杯中, 在同一测试条件下的人工气候箱中开始产卵, 记录雌雄成虫的个数和死亡时间, 每天将新产出的卵移至培养皿中, 统计产卵历期间、卵数及其孵化情况, 以方便统计成虫的繁殖力(所有卵的粒数/雌成虫的个数)。
按照试验结果剔除2个极端的温度梯度并按照正交试验设置2个因素, 即温度和光照强度, 以此筛选出对伸展摇蚊种群繁育的最佳试验条件。正交试验的每个因素有2个水平, 采用正交表L4(22), 见表 1。
| 水平 Level |
因素Factors | |
| A 温度 Temperature/ ℃ |
B 光照强度 Light intensity/lx |
|
| 1 | 25 | 800 |
| 2 | 30 | 2000 |
鉴于成虫羽化后的繁育除温度和光照外, 还可能受到空气湿度的影响, 因此在正交试验的最佳水平下选取相应的温度和光照条件, 在上述条件的人工气候箱中分别放入刚羽化的5对成虫, 设置4种空气湿度(45%、65%、85%和95%), 以此进一步考察湿度对成虫繁殖力的影响。
1.4 数据处理(1) 利用SPSS 19.0(IBM, USA)对相关试验结果进行统计分析:不同温度相同光照条件下对伸展摇蚊的卵、幼虫和蛹的存活率计算所得回归方程和回归系数进行F检验;对不同温度、不同光照条件下正交试验所得的发育历期和摇蚊总存活率以及雌雄比的结果采用双因素方差分析;采用单因素方差分析(one-way ANOVA)分析4种情况(25 ℃:800 lx、2000 lx;30 ℃:800 lx、2000 lx)下的伸展摇蚊雌雄比和繁殖力的变化。以上数据分析的显著性水平均为0.05。
(2) 参照相关文献[10-11]中的方法, 计算得到相应温度和光照条件下的种群净增殖率(R0)、平均世代周期(T)、内禀增长率(rm)、周限增长率(λ)和种群翻倍时间(t), 组建不同温度、不同光照的伸展摇蚊实验种群繁殖特征生命表。
2 结果 2.1 单因素试验表 2, 表 3分别表示在相同光照强度(800 lx或2000 lx)下摇蚊各形态存活率和4个温度水平(15、25、30、35 ℃)的关系。通过模拟出的温度与摇蚊各形态的存活率的方程计算可知, 在光照强度为800 lx(表 2)情况下, 温度在15 ℃和35 ℃时, 幼虫和蛹的存活率均受到抑制, 35 ℃条件下, 所有测试组卵孵化为一龄幼虫后即死亡;15 ℃条件下的6个测试组中, 5组幼虫停止发育, 另一组幼虫虽启动发育, 但发育速率缓慢。试验进行到第45 d时, 该组幼虫仅发育至三龄期, 所以高温(35 ℃)和低温(15 ℃)条件均无法维持幼虫正常发育。幼虫和蛹的存活率在25 ℃达到最高值, 分别为92.5%和98.87%, 当温度高于25 ℃时, 各存活率明显下降。幼虫和蛹的存活率的回归方程的相关系数均大于0.99, 显著性差异均小于0.05, 说明所得模型在15—35 ℃范围内, 摇蚊幼虫和蛹两种形态存活率与温度之间均呈显著相关。
| 发育阶段 Various stages |
回归方程 Regression equation |
相关系数 R2 |
F | P |
| 卵期Embryo | Y=-0.0237X2+1.3831X+79.298 | 0.972 | 17.454 | 0.167 |
| 幼虫期Larvae | Y=-0.9191X2+45.841X-478 | 0.998 | 921.472* | 0.023 |
| 蛹期Pupa | Y=-1.0096X2+50.564X-531.71 | 0.993 | 212.033* | 0.049 |
| *表示差异显著(P<0.05) | ||||
| 发育阶段 Various stages |
回归方程 Regression equation |
相关系数 R2 |
F | P |
| 卵期Embryo | Y=-0.0094X2+0.3906X+95.257 | 0.732 | 1.362 | 0.510 |
| 幼虫期Larvae | Y=-0.7168X2+35.074X-350.29 | 0.995 | 310.412* | 0.040 |
| 蛹期Pupa | Y=-0.9842X2+49.291X-518.33 | 0.997 | 211.866* | 0.048 |
| *表示差异显著(P<0.05) | ||||
在光照强度为2000 lx(表 3)情况下, 15 ℃时, 幼虫的存活率高于光照强度为800 lx时, 但是在25—35 ℃范围内, 卵、幼虫和蛹的存活率均低于摇蚊暴露于800 lx光照强度下。而在2000 lx光照下, 15 ℃更适合于摇蚊卵的孵化, 卵的存活率达到98.93%, 幼虫和蛹的存活率同样在25 ℃达到最高值, 且回归方程的相关系数均大于0.99, 显著性差异均小于0.05, 说明所得模型在试验温度范围内, 摇蚊幼虫和蛹两种形态存活率与温度之间均呈显著相关, 而与暴露于800 lx光照强度下相同的是, 当光照强度为2000 lx时, 幼虫期摇蚊的存活率比蛹期更易受温度影响, 这也反映了幼虫对环境条件变化响应更加敏感的特性。
通过图 1可知, 在15—35 ℃范围内, 卵都能保持较高的存活率。在15 ℃、800 lx条件下, 卵死亡率最高, 但也仅为5.36%。而在25 ℃时, 800 lx和2000 lx光照强度下的存活率分别为99.4%和99.2%, 所以在这4种温度中25 ℃属于存活率最高的温度条件;而在低温(15 ℃)和高温(35 ℃)条件下, 卵的存活率均有所下降。由此可见, 在本试验条件下, 随着温度的升高, 卵的存活率逐渐升高;当超过某一临界温度时, 卵的存活率又逐渐下降。结合表 2和表 3分析可以看出, 摇蚊卵的存活率与温度变化并没有显著相关, 所以本试验所设置的4种温度和两种光强对伸展摇蚊卵的存活率影响不大。

|
| 图 1 温度和光照强度对伸展摇蚊卵的死亡率的影响 Fig. 1 Effects of egg mortality of C. tentans at different temperature and light intensity |
由于较高和较低的温度(15 ℃和35 ℃)条件下, 幼虫不能完成世代发育, 所以为了确定在多因素条件下伸展摇蚊种群繁育的最佳条件, 在单因素的基础试验上, 剔除了两个极端温度后, 选取适合的参数范围, 对温度(A)、光照强度(B), 采用L4(22)进行正交试验, 优化最佳条件见表 4。表 4中列出了不同温度和光照组合的摇蚊存活率和发育时间的的测定结果, 直观的分析可以看出, 71.43%的存活率最高, 最佳试验条件为A1B1(25 ℃, 800 lx), 且由于RA>RB, 因此因素重要性为A>B(温度比光照重要)。
| 水平 Level |
A | B | 存活率 Survival rate/% |
水平 Level |
A | B | 发育速率 Development rate/d-1 |
|
| 1 | 1 | 1 | 85.43 | 1 | 1 | 1 | 0.029 | |
| 2 | 1 | 2 | 64.22 | 2 | 1 | 2 | 0.031 | |
| 3 | 2 | 1 | 49.83 | 3 | 2 | 1 | 0.041 | |
| 4 | 2 | 2 | 32.68 | 4 | 2 | 2 | 0.049 | |
| K1 | 149.65 | 135.26 | K1 | 0.060 | 0.070 | |||
| K2 | 82.51 | 96.90 | K2 | 0.090 | 0.080 | |||
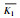 |
74.83 | 67.63 | |
0.030 | 0.035 | |||
 |
41.26 | 48.45 | |
0.045 | 0.040 | |||
| (极差Range)R | 33.57 | 19.18 | (极差Range)R | 0.015 | 0.005 | |||
| 优水平Optimization level | A1, B1 | 优水平Optimization level | A2, B2 | |||||
| 主次顺序Order | A>B | 主次顺序Order | A>B | |||||
注:Ki表示第i列上对应水平1(2, 3, 4)的数据和;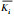 表示为i(1, 2, 3, 4)水平数据的综合平均;左边的表是关于摇蚊存活率的极差分析;右边是关于摇蚊发育速率的极差分析 表示为i(1, 2, 3, 4)水平数据的综合平均;左边的表是关于摇蚊存活率的极差分析;右边是关于摇蚊发育速率的极差分析 |
||||||||
在25 ℃条件下, 卵的平均孵化时间为1—2 d, 幼虫期为26.1—26.6 d, 全世代历期为32—35 d;30 ℃时, 卵的平均孵化时间缩短为1—1.3 d, 幼虫期缩短为14.7—17.2 d, 全世代历期缩短为20.4—24.5 d。且从表 4上看, 可知发育速率最快的最佳试验条件为A2B2(30 ℃, 2000 lx), 因素重要性为A>B(温度比光照重要)。可见, 温度和光照的增加有利于缩短发育历期, 提高发育速率。
表 5是对试验数据进行双因素方差分析的结果, 从表中可以直观的看出, 2个因素对摇蚊存活率和发育速率的影响中, 温度对其的影响均大于光照(PA<PB:0.038<0.065或0.127<0.330), 且温度对摇蚊存活率的影响极其显著(PA=0.038<0.05)。
| 变异来源 Source of variation |
平方和 Quadratic sum |
df | 均方 Mean square |
F | P | |
| 存活率 | 区间 | 1494.817 | 2 | 747.409 | ||
| Survival rate | A | 1126.945 | 1 | 1126.945 | 273.471* | 0.038 |
| B | 367.872 | 1 | 367.872 | 89.270 | 0.067 | |
| 误差 | 4.121 | 1 | 4.121 | |||
| 总和 | 14973.505 | 4 | ||||
| 变异来源 Source of variation |
平方和 Quadratic sum |
df | 均方 Mean square |
F | P | |
| 发育速率 | 区间 | 0.000 | 2 | 0.000 | ||
| Development rate | A | 0.000 | 1 | 0.000 | 24.452 | 0.127 |
| B | 0.000 | 1 | 0.000 | 3.064 | 0.330 | |
| 误差 | 0.000 | 1 | 0.000 | |||
| 总和 | 0.006 | 4 | ||||
| *表示显著(P<0.05) | ||||||
所以综上所述, 极差分析(表 4)和方差分析(表 5)得出的结论一致, 在影响摇蚊存活率和发育速率的因素中, 温度为最为重要的因素。在2种适当的条件下, 伸展摇蚊能顺利完成世代过程。在该温度(25—30 ℃)范围内, 升高温度将会可小幅度缩短摇蚊各阶段(卵、幼虫、蛹及成虫)发育历期, 提高发育速率, 但是这种影响并不显著, 而温度能显著影响摇蚊的存活率, 因此, 在2种因素条件下伸展摇蚊种群繁育的最佳条件是A1B1(25 ℃, 800 lx)。
图 2是25 ℃和30 ℃条件下温度和光强对伸展摇蚊性别比的影响结果。800 lx时, 雌雄比均接近于1。而对4种情况就雌雄比进行单因素方差分析的结果, 在25 ℃、2000 lx时雌性显著偏少(雌雄比为0.65), 而在30 ℃、2000 lx试验条件下, 摇蚊的雌性显著偏多, 性别比严重偏离1(雌雄比为2.4)。

|
| 图 2 温度和光照强度对伸展摇蚊性别比的影响 Fig. 2 Sex ratio of C. tentans at different temperature and light intensity *表示差异显著(P<0.05) |
根据正交试验关于雌雄比的方差分析(表 6)结果, 可以看出温度和光照对于摇蚊成虫的雌雄比影响并不显著, 但是从两者的显著性水平上看, 温度的重要性远大于光照强度(P:0.430<0.735), 所以, 温度对摇蚊性别比的影响远大于光照对其的影响。结合图 2也可以看出, 在相同的光照条件下, 不同的温度水平对雌雄比的影响差异极大;而在相同温度下, 两种光照强度对雌雄比的影响远小于前一种情况。所以, 综合图 2和表 6, 可知温度和光照强度对摇蚊雌雄比均有影响, 但其影响均不显著, 但温度的影响更大, 所以控制好温度范围对试验结果的正确性和合理性更为重要。
| 变异来源 Source of variation |
平方和 Quadratic sum |
df | 均方 Mean square |
F | P | |
| 雌雄比 | 区间 | 1.118 | 2 | 0.559 | ||
| Female male ratio | A | 0.994 | 1 | 0.994 | 1.558 | 0.430 |
| B | 0.124 | 1 | 0.124 | 0.195 | 0.735 | |
| 误差 | 0.638 | 1 | 0.638 | |||
| 总和 | 9.090 | 4 | ||||
| *表示差异显著(P<0.05) | ||||||
两种温度和两种光强对伸展摇蚊繁殖力的影响结果如表 7所示。产卵历期指羽化后开始产卵直至产卵结束所持续的时间。由结果可知, 25 ℃和30 ℃条件下, 产卵历期随温度升高而缩短、随光强增加而缩短。但根据4种情况的分析结果表明, 仅30 ℃、2000 lx光强条件下产卵历期显著低于其他测试组, 其他测试组之间并无显著差异。而30 ℃时每雌产卵量略低于25 ℃时, 但在本实验条件下差异并不显著。
| 指标 Index |
25℃ | 30℃ | |||
| 800 lx | 2000 lx | 800 lx | 2000 lx | ||
| 每雌产卵量Every female fecundity/(粒/雌) | 321±24 | 325±29 | 281±27 | 285±22 | |
| 产卵历期Oviposition duration/d | 8.3±0.6 | 6.3±0.6 | 6.7±0.6 | 4.3±0.6* | |
| *表示显著(P<0.05) | |||||
鉴于成虫羽化后的繁育除温度和光照外, 还可能受到空气湿度的影响, 因此图 3表明了在25 ℃、800 lx条件下的4种空气湿度(45%、65%、85%和95%)和成虫繁殖力的关系。如图所示, 85%和95%湿度时每雌产卵量显著高于其他两个湿度条件, 其中85%湿度时每雌产卵量最大。所以可以初步判断4个湿度中, 85%的空气湿度更适合于摇蚊产卵繁育。

|
| 图 3 湿度对伸展摇蚊成虫繁殖力的影响 Fig. 3 Sex ratio of C. tentans at different relative humidity *表示差异显著(P<0.05) |
根据25 ℃和30 ℃不同光强条件下所观察的种群生殖力资料, 计算出种群净增殖率(R0)、平均世代周期(T)、内禀增长率(rm)、周限增长率(λ)和种群翻倍时间(t)(表 8)。结果表明, 种群净增殖率(R0)与内禀增长率(rm)表现出相似的变化趋势, 在800 lx时明显高于2000 lx时, 30 ℃、2000 lx时最小。周限增长率(λ)均大于1, 表明种群在25 ℃和30 ℃不同光强条件下均作几何级数增加。30 ℃时种群的平均时代周期明显短于25 ℃时, 30 ℃、800 lx时种群翻倍时间(t)最短。虽然30 ℃时每雌产卵量略低于25 ℃时, 但30 ℃时雌性子代比例增加, 说明种群通过调整雌雄比来抵御外在环境的变化。
| 指标 Index |
25℃ | 30℃ | |||
| 800 lx | 2000 lx | 800 lx | 2000 lx | ||
| 净增殖率Net reproductive rate (R0) | 56.16 | 37.04 | 44.26 | 7.77 | |
| 平均世代周期Mean generation time (T) | 29.58 | 29.15 | 19.65 | 17.67 | |
| 内禀增长率Intrinsic rate of natural increase (rm) | 0.136 | 0.128 | 0.193 | 0.116 | |
| 周限增长率Finite rate of increase (λ) | 1.145 | 1.136 | 1.213 | 1.213 | |
| 种群翻倍时间Population doubling time (t) | 5.10 | 5.42 | 3.59 | 5.98 | |
| 子代雌雄比Female male ratio | 1.05:1 | 0.65:1 | 1.29:1 | 2.40:1 | |
根据上面的试验分析结果可知温度和光强对伸展摇蚊各生命阶段存活率和发育率、成虫性别比和繁殖力均有影响, 其中温度是最主要的因素。而本试验所设置的4种温度和两种光强对伸展摇蚊卵的存活率影响不大, 摇蚊卵的存活率与温度变化并没有显著相关, 出现上述现象的原因可能是由于卵外包被的卵壳在一定程度隔绝了外界不良环境条件的影响有关[12]。25—30 ℃是确保伸展摇蚊顺利完成世代发育过程的适宜温度范围, 在此温度范围内升温有助于缩短其发育历期。25 ℃、800 lx光强可以维持伸展摇蚊雌雄比接近于1, 从而确保种群稳定发展。高温(30—35 ℃)和强光(2000 lx)是导致伸展摇蚊死亡率提高的重要原因, 25 ℃、800 lx时卵、幼虫、蛹的死亡率均最低, 可以作为摇蚊种繁育的最佳条件。25—30 ℃、800—2000 lx条件未对伸展摇蚊成虫单雌繁殖量产生显著影响, 但高温(30 ℃)、强光(2000 lx)的综合作用显著缩短了成虫产卵历期。而对于羽化成虫而言, 85%—95%的相对湿度可以保证较高的产卵水平。
种群参数的变化是种群动态的重要体现, 而生命表则是获取种群参数的重要工具。根据不同温度和光强条件下伸展摇蚊实验种群生命表的研究结果, 800 lx光强、25—30 ℃时实验种群均维持较高的净增殖率(44.26—56.16), 且在此范围内升温可将平均时代周期从29.58 d缩短至19.65 d, 并由此导致内禀增长率由0.136升高至0.193.这说明伸展摇蚊在高温下采取了缩短寿命的生态对策, 从而抵消了高温下生殖力降低的不利影响。在此条件下, 伸展摇蚊种群数量每日增长1.145—1.213倍, 并在3.59—5.10 d实现种群翻倍。
综合考虑温度和光强对伸展摇蚊各生命阶段存活率和发育速率、成虫性别比和繁殖力以及种群动态的影响, 800 lx光强、25—30 ℃是保证摇蚊维持良好生命活力、顺利完成繁殖发育过程并维持稳定种群动态的适宜环境条件。为了保证羽化成虫较高的产卵水平, 还应维持85%—95%的相对湿度。值得注意的是, 本研究也证实伸展摇蚊幼虫是对外界胁迫响应最为敏感的生命阶段。因此保持适宜环境条件确保其存活率、发育率的基础上采用幼虫作为伸展摇蚊急性毒性测试的材料是比较合适的。本研究所得到的温度、光强及湿度对伸展摇蚊生长繁育的影响规律与前人所得到的环境条件对其他摇蚊种(Chironomus riparius、Chironomus kiiensis、Chironomus dorsalis)的影响规律相似, 但最适宜的繁育条件参数有所不同[1, 13-16]。
4 结论以我国广布摇蚊种——伸展摇蚊为研究对象, 本研究重点考察了温度、光强对各生命阶段存活率和发育速率、成虫性别比和繁殖力以及种群动态的综合影响, 结果表明:
(1) 在相同的光强(800 lx或2000 lx)条件下, 15—35 ℃温度范围内, 摇蚊幼虫和蛹两种形态存活率与温度之间均呈显著相关, 而幼虫期摇蚊的存活率比蛹期更易受温度影响, 这也反映了幼虫对环境条件变化响应更加敏感的特性。但是在试验条件范围内, 温度的变化对伸展摇蚊卵的存活率影响不大, 卵的存活率与温度变化并没有显著的相关性。且15 ℃和35 ℃两个极端温度不适合摇蚊的生存和繁育。
(2) 通过双因素作用下摇蚊存活率和发育速率的分析, 我们得出温度的重要性远大于光照强度, 伸展摇蚊种群繁育的最佳条件是25 ℃, 800 lx的结论。且温度和光照对于摇蚊成虫的雌雄比影响并不显著, 而在最佳条件下(25 ℃, 800 lx), 摇蚊的雌雄比为1.05, 更接近于1。
(3) 800 lx光强、25—30 ℃是保证伸展摇蚊维持良好生命活力、顺利完成繁殖发育过程, 而85%—95%的相对湿度是保证羽化成虫较高的产卵水平的必要条件。
(4) 伸展摇蚊幼虫是对外界胁迫响应最为敏感的生命阶段。在保持适宜环境条件确保其存活率、发育率的基础上采用幼虫作为伸展摇蚊急性毒性测试的材料是比较合适的。
| [1] | 方俊峰, 雷晓玲, 曹天洪, 宗祖胜. 环境因子对摇蚊幼虫存活的影响及杀灭试验研究. 净水技术, 2007, 26(1): 1–3. |
| [2] | 王雪龙. 摇蚊幼虫种类鉴定、龄期划分及各虫态发育历期的研究[D]. 上海: 上海海洋大学, 2015. http://cdmd.cnki.com.cn/Article/CDMD-10264-1015974702.htm |
| [3] | Wu F C, Meng W, Zhao X L, Li H X, Zhang R Q, Cao Y J, Liao H Q. China embarking on development of its own national water quality criteria system. Environmental Science & Technology, 2010, 44(21): 7992–7993. |
| [4] | Organisation for Economic Co-operation and Development. Guidelines for the Testing of Chemicals. Test No. 218: Sediment-Water Chironomid Toxicity Using Spiked Sediment. Paris: OECD, 2004. |
| [5] | Organisation for Economic Co-operation and Development. Guidelines for the Testing of Chemicals. Test No. 219: Sediment-Water Chironomid Toxicity Using Spiked Water. Paris: OECD, 2004. |
| [6] | Organisation for Economic Co-operation and Development. Guidelines for the Testing of Chemicals. Test No. 233: Sediment-Water Chironomid Life-Cycle Toxicity Test Using Spiked Water or Spiked Sediment. Paris: OECD, 2010. |
| [7] | United States Environmental Protection Agency. Using toxicity tests in ecological risk assessment. Intermittent Bulletin. EPA, 1994. |
| [8] | OECD. Guidance Document for Aquatic Effects Assessment. OECD Environment Monographs No. 92. Paris: OCED, 1995. |
| [9] | 姜永伟. 中国摇蚊属系统学研究[D]. 天津: 南开大学, 2011. http://d.wanfangdata.com.cn/Thesis/Y2003098 |
| [10] | 吴坤君, 陈玉平, 李明辉. 温度对棉铃虫实验种群生长的影响. 昆虫学报, 1980, 23(4): 358–368. |
| [11] | 孙兴滨. 摇蚊幼虫生态学特征及其在水处理过程中去除技术研究[D]. 哈尔滨: 哈尔滨工业大学, 2006. http://cdmd.cnki.com.cn/Article/CDMD-10213-2006136201.htm |
| [12] | Wang W B, Leopold R A, Nelson D R, Freeman T P. Cryopreservation of Musca domestica (Diptera: Muscidae) embryos. Cryobiology, 2000, 41(2): 153–166. DOI:10.1006/cryo.2000.2278 |
| [13] | Park K, Kwak I S. The effect of temperature gradients on endocrine signaling and antioxidant gene expression during Chironomus riparius development. Science of the Total Environment, 2014, 470-471: 1003–1011. DOI:10.1016/j.scitotenv.2013.10.052 |
| [14] | 方海平, 陈再忠. 光照诱导对摇蚊产卵的影响. 江苏农业科学, 2013, 41(10): 327–330. DOI:10.3969/j.issn.1002-1302.2013.10.121 |
| [15] | Sun X B, Lu Y, Sun L, Cui F Y. Effect of temperature on laboratory population of Chironomus kiiensis. Journal of Harbin University of Commerce: Natural Sciences Edition, 2013, 29(3): 292–297. |
| [16] | 孙兴滨, 王树涛, 崔福义. 二氧化氯对水源水中摇蚊幼虫灭活效果的研究. 中国环境科学, 2009, 29(6): 617–621. |