 2017, Vol. 37
2017, Vol. 37文章信息
- 邓扬悟, 唐纯, 袁红朝, 宁赵, 邹观发, 王久荣, 葛体达, 吴金水
- DENG Yangwu, TANG Chun, YUAN Hongchao, NING Zhao, ZOU Guanfa, WANG Jiurong, GE Tida, WU Jinshui.
- 13C脉冲标记法:不同生育期水稻光合碳在植物-土壤系统中的分配
- The 13C-CO2 pulsing labeling method: distribution of rice photosynthetic carbon in plant-soil systems during different rice growth stages
- 生态学报. 2017, 37(19): 6466-6471
- Acta Ecologica Sinica. 2017, 37(19): 6466-6471
- http://dx.doi.org/10.5846/stxb201607241506
-
文章历史
- 收稿日期: 2016-07-24
- 网络出版日期: 2017-05-27





2. 中国科学院亚热带农业生态研究所, 亚热带农业生态过程重点实验室, 长沙 410125;
3. 国家离子型稀土高效开发利用工程技术研究中心, 赣州 341000
2. Key Laboratory of Agro-ecological Processes in Subtropical Region, Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha 410125, China;
3. National Engineering Research Center for Ionic Rare Earth, Ganzhou 341000, China
植物通过光合碳同化途径将大气CO2转化成有机物质, 生成的光合碳经植物韧皮部运输分配到各组织部分。其中, 一部分光合碳输入地下部用于根系生长, 同时以根系沉积物(根系脱落物和根系分泌物)形式向土壤输入碳, 每年约有10%的大气CO2通过植被流向土壤[1]。植物光合碳是陆地生态系统“大气-植物-土壤”碳循环的重要组成部分, 也是土壤有机碳的重要来源[2]。全球大约10%的土地用于农业种植生产, 农田生态系统由于人类的耕种活动对碳循环产生了很大的干预和影响, 是陆地生态系统土壤碳循环研究的重要内容[3-4]。稻田是我国典型的农田生态系统;我国现有稻田面积340多万hm2, 约占全国耕地总面积的27%;我国水稻产量世界第一, 占全国粮食总产量约50%(《中国统计年鉴》, 2009—2014年均值)。稻田生态系统除了具有提供粮食等农产品的生产功能, 还在缓解全球气候变化和维持生物多样性等方面发挥着重要作用。因此, 定量研究作物光合碳在水稻各组织及地下部的分配, 对于全面认识植物-土壤间的互作关系、全球碳循环及土壤可持续利用有重要意义。
20世纪80年代以来, 随着稳定同位素技术的发展和应用, 碳、氮、氧、氢等轻元素由于在自然环境中的循环和周转过程中其同位素比值间的差异较大, 同位素分馏效应比较明显。利用13C/12C比值的变异具有原位标记特性[5], 通过测定土壤或者植物中碳同位素-δ13C自然丰度值, 可以研究植物光合碳的分配和去向。而且, 通过稳定碳同位素示踪(13C标记物), 可以指示和追踪相应C元素或其化合物在生物体及其环境介质中迁移、转化和积累, 有效地追踪植物光合碳在“植物-土壤”碳库中的动态变化和土壤碳储量的微小迁移与转换, 明确植物-土壤生态系统碳来源及其周转规律[6-7], 以及定量评价植物光合碳对土壤碳储量的相对贡献等[7-9]。目前, 常用的稳定同位素检测方法有:质谱法、核磁共振法和光腔衰荡光谱法等, 其中质谱法是稳定碳同位素分析中最通用、最精确的方法[10-11]。本文通过对不同生育期的水稻进行13C-CO2脉冲标记, 并采用稳定同位质谱仪—元素分析仪联用, 量化水稻光合碳在植株各组织及土壤中的分配, 剖析不同生育期水稻光合碳的分配规律, 进一步探明农田土壤有机碳循环的过程和特征, 以期为农田土壤有机碳的调控和土壤可持续管理提供理论依据。
1 材料和方法 1.1 材料供试土壤为亚热带地区典型红壤, 采自中国科学院长沙农业环境观测研究站, 采样时间2015年4月。土壤的基本理化性状:土壤容重1.31 g/cm3, 有机碳18.1 g/kg, 全氮1.8 g/kg, 全磷0.43 g/kg, pH 5.6。供试植物为籼性常规水稻(中早39)。
标记材料为Na2CO3-13C:99.0 atom %(Sigma;CAS 93673-48-4)。
1.2 方法 1.2.1 水稻13C-CO2脉冲标记按照田间氮肥施入量, 预先向土壤中加入200 mg N / kg土的(NH4)2SO4为氮源。将5 kg(干土重计)水稻土装于PVC盆(直径17.2 cm, 高16.7 cm)中, 添加蒸馏水至淹水2—3 cm。选择水稻三叶期幼苗移栽至PVC盆中, 每盆3株, 待水稻生长稳定后, 分别在水稻分蘖期、拔节期、抽穗期、扬花期、成熟期等5个生育期分别进行标记, 每次标记3盆。水稻13C-CO2脉冲标记在密闭植物生长箱中进行, 标记箱置于光照充足的户外, 每次标记6 h(9:00—15:00)。标记生长箱和方法参考肖和艾等[12]建立, 并作适当修改[13]。培养箱内CO2浓度控制范围为350—370 mg/kg(CO2检测系统;Shen-QZD), 13C-CO2由1 mol/L的13C-Na2CO3(丰度为99%的13C-Na2CO3)缓慢匀速滴入1 mol/L HCl反应产生(反应完全), 温度控制在24—32℃(温度控制器, SNT-96S)。与此同时, 准备3盆上述水稻放置于离标记培养箱10 m以上的区域作为对照, 用来测定植株及土壤13C的自然丰度。
1.2.2 样品的制备在每次标记结束后1 h内, 同时对标记和非标记的植物和土壤进行采集。根和秧苗洗净沥干后, 置于烘箱中, 105℃下杀青30 min, 杀青后的植物样品放置于60—70℃的干燥箱中干燥24 h, 称重并计算根、茎叶生物量。烘干的植物样品用行星式球磨仪进行磨碎, 供测定其有机碳含量和δ13C值使用。土壤样品充分混匀并挑出残余植物根系后立即进行冷冻干燥24 h, 经过干燥后的样品用球磨仪粉碎, 过60目筛, 供测定其有机碳含量和δ13C值使用。
1.2.3 分析方法用Flash HT-IRMS测定植物和土壤样品的有机碳、氮含量及其δ13C值。其分析的基本原理和测定过程是:样品经高温燃烧后(氧化/还原炉温度为950℃), 通过TCD (Thermal Conductivity Detector)检测器测定有机碳、氮含量, 剩余气体进入ConFlo IV导入稳定同位素质谱仪, 在质谱仪上测定δ13C值。
植株(根系和地上部)和土壤中13C含量的计算:
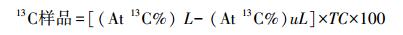
|
式中, 13C样品表示样品中13C含量, At 13C%表示样品中含13C的丰度; L和uL分别表示标记和非标记;TC样品表示样品中总有机碳的含量。
1.2.4 分析与统计数据统计分析采用SPSS 13.0 for Windows和Microsoft Excel 2010软件进行。差异显著性用Oneway ANOVA(一维方差)分析, 多重比较采用Duncan法。
2 结果与讨论 2.1 水稻地上部和根系生物量水稻地上部和根系干物质量分别为4.9—40.9 g/盆和2.0—8.1 g/盆, 并随水稻生育期的增加而呈现递增趋势(图 1), 分蘖期>拔节期>抽穗期>扬花期>成熟期。水稻地上部植株由于水稻生育期不同增长趋势差异显著(P < 0.05), 从分蘖期到拔节期增幅最大, 植株生长率增加了2—3倍, 而扬花期至成熟期植株干物质重仅增加了20%左右。比较不同生育期水稻根系的生物量, 发现分蘖期到拔节期根系出现跳跃式生长(P < 0.05), 增长幅度达到150%以上, 而到抽穗期之后, 根系生物量未现明显增加(P>0.05)。

|
| 图 1 不同生育期水稻地上部和根部干物质重 Fig. 1 The rice biomass of above ground and below ground during different growth stages |
水稻根冠比为0.2—0.4, 其中分蘖期根最高, 随着水稻生育期的增加而递减, 到抽穗期以后比值稳定在0.2左右。分析其原因可能在分蘖期水稻光合碳主要运移到地下部用于根系生长, 到拔节期后水稻光合碳向地下部运移用于根系生长的比例下降, 并逐渐向水稻茎叶及稻穗倾斜。GE等[14]的研究也指出植物光合碳的地下部输入量与根系生长速度密切相关。植物在营养生长期, 根系碳库活力高, 需转运更多的碳, 而成熟期用于根系组织形成, 根系呼吸及根际沉积的光合碳比例下降。
2.2 光合碳δ13C在水稻各组织中的分馏效应非标记的不同生育时期水稻植株各部分δ13C值见表 1。水稻植株地上部和地下部(根系)的δ13C值在-25.52‰—-28.33‰, 不同器官的δ13C值存在明显分馏效应, 且趋势基本一致, 即茎杆(麦穗)>叶片(根系)(表 1)。
| 样品 Sample/δ13Cvs PDB‰ |
分蘖期 The tillering stage |
拔节期 The jointing stage |
抽穗期 The heading stage |
扬花期 The flowering stage |
成熟期 The maturity stage |
| 叶片Leave | -27.75±0.27 a | -28.02±0.22 a | -27.83±0.20 a | -27.68±0.16 b | -27.82±0.24 b |
| 茎杆Stem | -25.52±0.31 b | -25.83±0.15 b | -25.71±0.21 c | -25.92±0.18 c | -26.01±0.33 c |
| 稻穗Grain | - | - | -26.35±0.20 b | -26.15±0.12 c | -26.41±0.22 c |
| 根系Root system | -27.83±0.30 a | -28.33±0.41 a | -28.22±0.27 a | -28.30±0.32 a | -28.31±0.26 a |
| 每列不同字母表示5%的显著差异 | |||||
这种分馏现象主要是由于13CO2的质量大于12CO2, 大气中的13CO2进入植物叶片的扩散速率低于12CO2, 而且表现出相对较低的与核酮糖1, 5二磷酸羧化氧化酶(Rubisco)的结合能力, 因此水稻植物碳水化合物的13CO2 与12CO2的比值远低于大气中13CO2与12CO2 的比值[15], 为-25.52‰—-28.33‰。水稻叶片通过光合碳作用生成的光合碳在传输过程中同样会出现同位素分馏, 表现为叶片(根系)和茎秆的同位素分馏效应明显, 差值可达2.0‰。这种水稻生育期特性导致水稻植株各器官碳同位素分馏的现象[16], 可用于指示不同生育期下水稻光合碳的分配和去向。在水稻营养生长期, 由于根系活力强, 会有较多的贫化型光合碳向根系输送, 表现出较低的根系δ13C值, 并在分蘖期到拔节期出现根系δ13C值跳跃式下降, 差值可达0.6‰。而成熟期, 即抽穗期之后, 根系δ13C值未明显变化, 说明水稻根系组织已形成, 无需向根系转运更多的碳。
2.3 不同生育期光合碳13C在水稻-土壤系统中的分配水稻植株通过光合作用合成的有机碳通过茎中的韧皮部传递至根系, 通过根系沉积作用进入土壤。采用稳定碳同位素示踪(13C-CO2标记物), 追踪了植物光合碳在“植物-土壤”碳库中的动态变化。水稻植株茎叶、根系及土壤中的13C含量分别为:16.6—112.6 mg/盆, 3.8—23.7 mg/盆和2.6—5.1 mg/盆(表 2)。水稻-土壤系统中13C光合碳随着水稻生育期而增加, 其中从分蘖期到拔节期递增比例最大, 达3倍以上, 其次为抽穗期。这段时期不仅水稻生物量增加比例较大, 光合碳的地下部分配量的也显著增加, 而水稻根系光合碳输入有利于土壤有机碳累积。
| 生育期 Growth stages |
13C-茎叶 13C-shoots/ (mg/盆) |
13C-根系 13C-root system/(mg/盆) |
13C-土壤有机碳 13C-SOC/(mg/盆) |
| 分蘖期The tillering stage | 16.59 ± 2.77 a | 3.81 ± 0.65 a | 2.56 ± 0.12 a |
| 拔节期The jointing stage | 64.44 ± 5.96 b | 5.57 ± 1.73 b | 2.79 ± 0.14 a |
| 抽穗期The heading stage | 94.39 ± 3.02 c | 7.20 ± 0.88 c | 3.61 ± 0.17 b |
| 扬花期The flowering stage | 111.64 ± 4.71 d | 11.10 ± 1.30 d | 5.03 ± 0.11 c |
| 成熟期The maturity stage | 112.57 ± 8.64 d | 23.70 ± 1.16d | 5.11 ± 0.28 c |
不同生育期13C光合碳在水稻-土壤系统中的分配比例差异明显(图 2), 其中植物茎叶所占比例最大为72%—81%, 并随着生育期增加而增加, 到扬花期达到最大值并稳定在81%左右;水稻13C-光合碳地下部根系所占比例为10%—18%, 而土壤中13C-光合碳仅占3.8%—10%, 且两者比例随着水稻生育期增加而减少。这和Lu等[17]的水稻生育期内标记的多数的同化13C保留在地上部分, 小部分的同化13C出现在地下的结果一致。水稻生长前期向地下部分配的碳量多于成熟期, 这是由于水稻不同生育期的生理特性决定的, 生长前期水稻根系碳库活力高需转运更多碳。在本研究中水稻分蘖期有近30%光合碳用于根系建成并部分通过根系分泌物进入土壤有机碳库(10%), 而到成熟期则向籽粒中分配较多, 而且光合碳在土壤中的分配比例也随生育期呈下降趋势。Werth等[18]也指出在水稻生长前期, 光合碳向根系及土壤中分配的比例高, 光合碳在根系及土壤中的分配比例随生育期呈下降趋势, 但积累量不断增加。

|
| 图 2 不同生育期水稻13C-光合碳在植物-土壤系统中的分配 Fig. 2 The allocation of photosynthesized 13C into the rice plant-soil system during different growth stages |
通过稳定同位素质谱分析技术准确量化植物光合碳及其周转过程对于深入认识陆地生态系统碳循环过程和准确估算全球碳平衡有着重要意义[19]。本实验采样碳稳定同位素脉冲标记研究了水稻不同生育期的光合碳在植物-土壤系统中的分配信息, 发现水稻植株各器官碳同位素分馏现象的客观存在。水稻-土壤系统中13C光合碳量随生育期增加, 且地下部分进入土壤有机碳库光合碳量也随之增加, 水稻根系光合碳的输入有利于土壤有机碳累积。不同生育期光合碳在植物-土壤系统的分配规律不同, 生长前期光合碳向根系及土壤中分配的比例高, 具有较强的C汇能力;而随生育期光合碳在根系及土壤中的分配比例呈下降趋势, 但积累量不断增加。
| [1] | Uchida Y, Hunt J E, Barbour M M, Clough T J, Kelliher F M, Sherlock R R. Soil properties and presence of plants affect the temperature sensitivity of carbon dioxide production by soils. Plant and Soil, 2010, 337(1/2): 375–387. |
| [2] | Schlesinger W H, Andrews J A. Soil respiration and the global carbon cycle. Biogeochemistry, 2000, 48(1): 7–20. DOI:10.1023/A:1006247623877 |
| [3] | 于贵瑞, 高扬, 王秋凤, 刘世荣, 申卫军. 陆地生态系统碳氮水循环的关键耦合过程及其生物调控机制探讨. 中国生态农业学报, 2013, 21(1): 1–13. |
| [4] | 姜勇, 庄秋丽, 梁文举. 农田生态系统土壤有机碳库及其影响因子. 生态学杂志, 2007, 26(2): 278–285. |
| [5] | Dignac M F, Bahri H, Rumpel C, Rasse D P, Bardoux G, Balesdent J, Girardin C, Chenu C, Mariotti A. Carbon-13 natural abundance as a tool to study the dynamics of lignin monomers in soil: an appraisal at the Closeaux experimental field (France). Geoderma, 2005, 128(1/2): 3–17. |
| [6] | Deng L, Wang K B, Tang Z S, Shangguan Z P. Soil organic carbon dynamics following natural vegetation restoration: Evidence from stable carbon isotopes (δ13C). Agriculture Ecosystems & Environment, 2016, 221: 235–244. |
| [7] | Kuzyakov Y, Domanski G. Carbon input by plants into the soil.Review. Journal of Plant Nutrition and Soil Science, 2000, 163(4): 421–431. DOI:10.1002/(ISSN)1522-2624 |
| [8] | Guillaume T, Damris M, Kuzyakov Y. Losses of soil carbon by converting tropical forest to plantations: erosion and decomposition estimated by δ 13C. Global Change Biology, 2015, 21(9): 3548–3560. DOI:10.1111/gcb.2015.21.issue-9 |
| [9] | 田秋香, 张威, 闫颖, 何红波, 张旭东, 郑立臣. 稳定性同位素技术在土壤重要有机组分循环转化研究中的应用. 土壤, 2011, 43(6): 862–869. |
| [10] | Paul D, Skrzypek G, Fórizs I. Normalization of measured stable isotopic compositions to isotope reference scales-a review. Rapid Communications in Mass Spectrometry, 2007, 21(18): 3006–3014. DOI:10.1002/(ISSN)1097-0231 |
| [11] | Eiler J M, Clog M, Magyar P, Piasecki A, Sessions A, Stolper D, Deerberg M, Schlueter H J, Schwieters J. A high-resolution gas-source isotope ratio mass spectrometer. International Journal of Mass Spectrometry, 2013, 335: 45–56. DOI:10.1016/j.ijms.2012.10.014 |
| [12] | 肖和艾, 吴金水, 李玲, 唐国勇, 刘畅. 采用14C同位素标记植物的装置与方法. 核农学报, 2007, 21(6): 630-632, 629-629. |
| [13] | Ge T D, Yuan H Z, Zhu H H, Wu X H, Nie S A, Liu C, Tong C L, Wu J S, Brookes P. Biological carbon assimilation and dynamics in a flooded rice-soil system. Soil Biology and Biochemistry, 2012, 48: 39–46. DOI:10.1016/j.soilbio.2012.01.009 |
| [14] | Ge T D, Liu C, Yuan H Z, Zhao Z W, Wu X H, Zhu Z K, Brookes P, Wu J S. Tracking the photosynthesized carbon input into soil organic carbon pools in a rice soil fertilized with nitrogen. Plant and Soil, 2015, 392(1/2): 17–25. |
| [15] | Schubert B A, Jahren A H. The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants. Geochimica et Cosmochimica Acta, 2012, 96: 29–43. DOI:10.1016/j.gca.2012.08.003 |
| [16] | O'Leary M H, Madhavan S, Paneth P. Physical and chemical basis of carbon isotope fractionation in plants. Plant, Cell & Environment, 2006, 15(9): 1099–1104. |
| [17] | Lu Y H, Murase J, Watanabe A, Sugimoto A, Kimura M. Linking microbial community dynamics to rhizosphere carbon flow in a wetland rice soil. FEMS Microbiology Ecology, 2004, 48(2): 179–186. DOI:10.1016/j.femsec.2004.01.004 |
| [18] | Werth M, Kuzyakov Y. 13C fractionation at the root-microorganisms-soil interface: a review and outlook for partitioning studies. Soil Biology and Biochemistry, 2010, 42(9): 1372–1384. DOI:10.1016/j.soilbio.2010.04.009 |
| [19] | Blaser M, Conrad R. Stable carbon isotope fractionation as tracer of carbon cycling in anoxic soil ecosystems. Current Opinion in Biotechnology, 2016, 41: 122–129. DOI:10.1016/j.copbio.2016.07.001 |