 2017, Vol. 37
2017, Vol. 37文章信息
- 刘海, 杜如万, 王勇, 陈玉蓝, 吴叶宽, 袁玲
- LIU Hai, DU Ruwan, WANG Yong, CHEN Yulan, WU Yekuan, YUAN Ling.
- 紫茎泽兰对四川省凉山州共生植物种间联结性及稳定性的影响
- Effects of Eupatorium adenophorum on interspecific association and the stability of companion species in Liangshan Prefecture of Sichuan Province
- 生态学报. 2017, 37(15): 5031-5038
- Acta Ecologica Sinica. 2017, 37(15): 5031-5038
- http://dx.doi.org/10.5846/stxb201605070876
-
文章历史
- 收稿日期: 2016-05-07
- 网络出版日期: 2017-03-22





2. 贵州省农业科技信息研究所, 贵阳 550006;
3. 四川省凉山州烟草公司, 西昌 615000
2. Guizhou Institute of Agricultural Science and Technology Information, Guiyang 550006, China;
3. Sichuan Liangshanzhou Tobacco Company, Xichang 615000, China
外来生物入侵是导致全球生物多样性降低的重要因素, 严重破坏入侵地生态系统的完整性[1], 随着全球化进程的推进, 入侵速度越来越快[2], 造成当地生态系统失衡, 本地物种生存受到威胁, 并因此造成巨大的经济损失, 已引起社会各界的广泛关注[3]。由此衍生的新学科——入侵生态学, 随着入侵生物的增加和入侵程度的加深逐渐成为新的研究热点[4]。除研究入侵生物本身的入侵特性外, 入侵生物对本土物种的影响及本土物种对入侵生物入侵行为的响应, 也是入侵生态学的研究重点。探讨生物入侵后群落中各物种间的联结性和稳定性是研究入侵生物和本土物种间相互作用的重要切入点[5-6]。
种间联结性是群落中不同物种间相互关系的静态反映, 这种关系是在群落不断演替过程中不同物种间相互作用形成的, 其不仅是群落形成和演化的基础, 也是人们区分群落差异和描述群落中不同物种间相互关系的重要依据[6]。分析不同物种间关联程度对探索物种间关系及其对不同环境的响应、分析群落中物种组成及分布、种群进化过程、群落分类及演化规律具有重要参考意义[7]。种间联结性研究以样方中物种出现与否的二元数据构成2×2联列表进行计算, 常用指标包括多物种间联结性、种间联结系数、χ2统计量、共同出现百分率、Ochiai系数和Dice系数等。郭志华等[8]发现, 以χ2统计量为基础, 结合AC和PC等指标研究得出植物种群联结性的效果较好。近年来, 众学者对陕西子午岭森林植物群落[9]、野生黄花蒿群落[10]和长白山森林群落[11]等种间联结性进行了分析, 但通过分析不同物种间的种间联结性探索入侵生物与入侵地物种关系的研究相对较少[12]。
外来入侵植物紫茎泽兰(Eupatorium adenophorum)为菊科泽兰属多年生恶性毒草, 自20世纪40年代由中美洲经缅甸和越南传入中国以来, 至今已大面积侵入云南、贵州、四川、重庆、广西和西藏等地的农田、森林和草地等, 海拔1000—1900 m均有分布, 不仅有进一步向高海拔地区(2500 m)入侵的趋势, 更是以每年30—60 km的速度向东、向北蔓延[13]。据调查, 2012年四川省凉山州超过10%的面积已被紫茎泽兰侵占[14], 严重破坏了当地生态平衡, 同时也给农业生产造成巨大的损失。有关紫茎泽兰群落中不同物种的种间联结性和稳定性的研究尚未见报道。笔者以四川省凉山州危害最严重的入侵植物紫茎泽兰群落为研究对象, 探讨了紫茎泽兰种群与其伴生种的种间关系, 旨在为及早预测和评估紫茎泽兰入侵广度与深度提供依据和参考。
1 材料与方法 1.1 研究区概况研究地点为四川省凉山州, 该州处于100°15′—103°53′E和26°03′—29°27′N之间, 北靠大渡河, 南临金沙江, 东接四川盆地, 西连横断山脉, 境内地貌复杂多样, 土壤类型繁多。幅员面积6万km2。属高原亚热带印度洋季风气候, 冬暖夏凉, 干温季明显, 年降雨量1000—1100 mm, 年均温14—17℃, 日照时数为2000—2400 h, 年日照辐射总量为120—150 kCal/cm2, 无霜期为230—306 d。其独特的地理条件和气候条件造就了该地区生物种类丰富多样。20世纪80年代末, 紫茎泽兰仅在凉山州西南部金沙江沿线有分布, 2007年已遍布全州15个县市, 至2012年, 紫茎泽兰的危害面积已达61.2万hm2[14]。目前, 紫茎泽兰入侵面积仍在不断扩大中, 凉山州的生态安全遭受极大威胁[15-18]。
1.2 样方设置与统计方法2015年8月3日至18日, 在凉山州紫茎泽兰危害较严重的西昌市、会理县、会东县、德昌县和普格县, 每个县市在紫茎泽兰分布片区沿等高线设置10块样地, 共计50块样地, 为科学设置样地, 保证样地的一致性, 根据紫茎泽兰长势, 并结合当地农民的描述, 所选样地紫茎泽兰生长年限均为4 a左右。每块样地以10 m×10 m为样方单位, 重复3次, 记录样方内出现的植物种类(科、属、种)和出现频度等指标, 并剔除3次重复中仅出现1次的种, 采用Excel 2007进行数据整理, OriginPro 9.0绘图软件作图。
1.3 总体联结性采用方差比率法测定紫茎泽兰群落的总体关联性, 并采用统计量W检验关联显著性[19]。计算公式如下:

|
式中, S为总物种数, N为总样方数, Tj为样方j内出现的物种数, ni为物种i出现的样方数, t为样方中种的平均数。
VR大于1表示物种间总体上表现为正关联, 小于1则表示负关联, 用统计量W=N×VR检验VR值相对1的偏移程度, W值落入χ0.952(N)<W<χ0.052(N), 则表示物种间无显著关联性。
1.4 种间联结性 1.4.1 χ2统计量鉴于取样的非连续性, 用2×2联列表统计各种对的具体值, χ2值采用Yates的连续校正公式[20-21]计算:

|
式中, n为总样方数。当χ2<3.841时, 种对间联结独立;当χ2>6.635时, 种对间存在显著生态联结;当3.841<χ2<6.635时, 种对间有一定的生态联结。ad>bc为正联结, 反之为负联结。
1.4.2 联结系数AC联结系数AC可进一步检验χ2值和种对间的联结程度。当ad≥bc时, AC=(ad-bc)/[(a+b)(b+d)];当bc>ad且d≥a时, AC=(ad-bc)/[(a+b)(a+c)];当bc>ad且d<a时, AC=(ad-bc)/[(b+d)(d+c)]。
AC值域为[-1, 1], AC值越接近1, 说明物种间的正联结性越强;越接近-1, 说明负联结性越强;AC值等于0, 说明物种间完全独立[21]。
1.4.3 共同出现百分率PC该指标用以测度种对间的正联结程度。计算公式为PC=a/(a+b+c)。PC值为0时表示无关联, 越接近1, 表示正联结越强[21-22]。
1.4.4 其他测度系数Ochiai指数(简称OI指数)表示种对同时出现的几率和联结程度, 可以克服点联结系数AC受d值影响较大造成的偏差。计算公式如下:
以上计算公式中, a为两物种同时存在的样方数, b为仅B物种存在而A物种不存在的样方数, c为A物种存在而B物种不存在的样方数, d为两物种均不存在的样方数。由于紫茎泽兰出现频度为100%, 其与任何种配对时的b和d值均为0, 为避免分母为0无法计算的情况出现, 将b和d值加权为1, 以获得较佳的客观效果[7]。
1.5 紫茎泽兰群落稳定性参考Gordon M等[24]的方法, 首先将10种优势植物种按频度由大到小依次排列, 换算为相对频度后逐步累积;再将样地内出现的植物种类总数取倒数, 对应逐步累积;两组数据一一对应, 以植物种类倒数百分率为横坐标, 累积频度为纵坐标, 建立模糊散点曲线模型。然后将横纵坐标100处直线连接, 其与曲线交点即群落稳定性比值。该值越接近20/80, 说明群落越稳定, 反之则越不稳定[25]。
平滑曲线模拟模型方程为: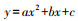
直线方程为:
将直线方程代入平滑曲线模拟模型方程:

|
交点坐标为:

|
式中, x为群落物种总数的倒数累积值, y为物种累积相对频度, a、b和c为回归曲线模型参数。该方程有2个解, 取0—100的解(另一个解远大于100, 对本文而言为无效值)。x/y值与20/80的接近程度可表征群落的稳定程度, 越接近则越稳定, 反之则越不稳定。
2 结果与分析 2.1 种群间的总体联结性根据50个样方统计资料, 紫茎泽兰群落共有155种植物。其中, 草本植物86种, 灌木34种, 乔木22种, 藤本植物10种, 蕨类植物2种, 苔藓植物1种。隶属于62科37亚科135属。种类较多的科有菊科(Asteraceae, 15属17种)、禾本科(Poaceae, 13属13种)、蔷薇科(Rosaceae, 10属12种)、豆科(Leguminosae, 7属7种)和山茶科(Theaceae, 4属6种)等。对紫茎泽兰群落中出现频率最高的10个物种进行关联性分析, 除紫茎泽兰外的9个物种分别为鬼针草(30次)、野艾蒿(28次)、马唐(26次)、灯笼草(25次)、黄花稔(18次)、爵床(15次)、地果(15次)、毛叶悬钩子(13次)和狗尾草(12次)。
根据10个优势种群的存在与不存在矩阵, 代入1.3各公式计算VR值(VR=ST2/δT2=0.9677<1), 结果表明, 10个物种间表现出负联接性。计算统计量W来检测VR值偏离1的程度, W=N×VR=48.38, 查表χ0.952(50) =34.76, χ0.052(50) =67.50, W值落入二者之间, 紫茎泽兰群落主要物种间总体呈不显著负关联。说明, 紫茎泽兰群落优势种群对环境的适应性相似甚至相同, 但对资源利用存在竞争, 总体呈不显著负关联。紫茎泽兰群落处于非稳定状态, 种间会出现一定程度的分离, 以降低竞争强度, 从而逐步趋于稳定。
2.2 主要种对间的关联性 2.2.1 χ2统计量探讨不同种对间的关联性对合理解释种对间的相互关系, 并进一步了解其规律具有重要意义。由表 1可以看出, 种群间χ2统计量均小于3.841, 仅2—9(鬼针草-毛叶悬钩子)和3—9(野艾蒿-毛叶悬钩子)的χ2统计量略大, 分别为3.16和3.26。说明主要种对间均无显著关联性。根据χ2检验原理可知, 主要种群的种对间联结性较低, 甚至趋于独立。这可能与物种本身的生态学特征和群落发展程度有关。
| 1 | |||||||||
| 0.07 | 2 | ||||||||
| 0.27 | 0.03 | 3 | |||||||
| 0.39 | 0.00 | 0.37 | 4 | ||||||
| 0.46 | 0.08 | 0.08 | 0.08 | 5 | |||||
| 0.10 | 0.18 | 0.06 | 0.26 | 0.09 | 6 | ||||
| 0.02 | 0.10 | 0.47 | 0.03 | 3.43 | 1.49 | 7 | |||
| 0.02 | 0.89 | 0.31 | 0.03 | 0.38 | 0.33 | 0.00 | 8 | ||
| 0.00 | 3.16 | 3.26 | 0.03 | 0.00 | 0.01 | 0.18 | 0.08 | 9 | |
| 0.00 | 0.77 | 0.66 | 0.03 | 0.99 | 1.58 | 0.01 | 0.01 | 0.08 | 10 |
| 1—10分别指紫茎泽兰、鬼针草、野艾蒿、马唐、灯笼草、黄花稔、爵床、地果、毛叶悬钩子和狗尾草 | |||||||||
由于χ2检验的种对间均无显著关联, 需借助AC值和PC值来反映种间联结性相对强弱。由图 1可知, AC值在[0.6, 1]、[0.3, 0.6)、(0, 0.3)、0、[-0.3, 0) 和[-0.6, -0.3) 的种对数量分别占总数的2.22%、8.89%、33.33%、6.67%、37.78%和11.11%。其中, 种对1—5(紫茎泽兰-灯笼草)、2—5(鬼针草-灯笼草)和2—7(鬼针草-爵床)AC值为0, 处于完全独立的状态;20个种对表现出正关联, 22个种对表现出负关联。种对2—9(鬼针草-毛叶悬钩子)的AC值(0.62) 大于0.6, 二者间存在明显的正关联性。种对6—10(黄花稔-狗尾草, -0.54)、3—9(野艾蒿-—毛叶悬钩子, -0.45)、6—7(黄花稔—爵床, -0.44)、1—10(紫茎泽兰-狗尾草, -0.33) 和1—9(紫茎泽兰-毛叶悬钩子, -0.32) 的负关联性较高。其余大部分种对AC值在0上下波动, 关联程度较弱。

|
| 图 1 紫茎泽兰群落联结系数、共同出现百分率、OI指数和DI指数的半矩阵图 Fig. 1 Semi-matrixes of association coefficients, co-occurrence rate, Ochiai (OI) index and Dice (DI) index of E. adenophorum community OI指数即Ochiai指数, DI指数即Dice指数, 二者均表示种对同时出现的几率和联结程度, 越接近1, 关联程度越强 |
共同出现百分率PC值均未超过0.6, PC值在[0.4, 0.6) 的种对有5个, 分别为1—2(紫茎泽兰-鬼针草)、1—3(紫茎泽兰-野艾蒿)、2—3(鬼针草-野艾蒿)、1—4(紫茎泽兰-马唐)和1—5(紫茎泽兰-灯笼草), 占总数的11.11%, 表现出较高的关联度;[0.2, 0.4) 的种对有27个, 占总数的60.00%, 有一定的关联性;[0, 0.2) 的种对有13个, 占总数的28.89%, 存在微弱关联性。
2.2.3 OI指数和DI指数OI指数可以克服点联结系数AC值受d值影响误差偏大的问题。由图 1可见, 种对1—2(紫茎泽兰-鬼针草)、1—3(紫茎泽兰-野艾蒿)、1—4(紫茎泽兰-马唐)和1—5(紫茎泽兰-灯笼草)的OI值大于0.6。说明紫茎泽兰与鬼针草、野艾蒿、马唐和灯笼草种对间有非常紧密的关联性, 相伴出现几率最大;种对6—7(黄花稔—爵床)和6—10(黄花稔-狗尾草)的OI值非常小, 分别为0.18和0.14。说明黄花稔与爵床、狗尾草的关联程度较低, 基本独立出现;其余绝大多数种对OI值集中在0.2—0.6, 关联程度一般。
DI指数与OI指数基本一致, 仅种对4—8、1—10和4—10的值略有出入。
2.3 群落稳定性由图 2可知, 平滑曲线模拟模型方程与直线方程的交点为51/49, 距离稳定交点坐标20/80较远, 说明群落处于一定的不稳定状态, 不同种群间对土壤养分、光照等的竞争剧烈。

|
| 图 2 紫茎泽兰群落的稳定性图解 Fig. 2 Stability illustration of E. adenophorum community |
种间联结程度衡量了不同物种间的相互关系和植物对环境因子的适应程度, 正联结体现了物种利用自然资源的相似性和生态位的重叠, 说明其具有相似的生境需求[26]。比较χ2检验结果与AC、PC、OI和DI指数, 反映的紫茎泽兰群落种间联结关系基本一致, 说明以上方法用于判断紫茎泽兰群落内不同种群间的联结性是可行的, 尤其OI指数与DI指数拟合度极高。χ2检验以损失种间相关多度信息为代价, 以不同物种在样方内出现与否的二元数据作为判断联结性的标准, 虽只能作定性判断, 但胜在分析方法简便, 且有明确的判断指标(P<0.01和P<0.05), 能客观、准确地反映种对间的联结性。当χ2检验不显著时, AC、PC、OI和DI指数均能反映出种对间的联结程度, 但对于联结性强弱至今仍无统一的评判标准。当样方中某一物种出现频率较低, 特别当a=0时, 均会夸大不同种对间的联结程度, 甚至得出相反的结论。当物种较多、取样数量较大的情况下, PC、OI和DI指数均能避免d值较大导致的AC偏高的缺点, 但三者仅能反映不同种对间正联结性的强弱, 当a值较小时, 忽略d值的存在, 夸大了a, b, c的作用, 从而导致负联结指数偏高。三者反映不同种对相伴出现的机率和联结程度, 值越大, 同时出现的机率越大, 但并不意味着一定是正联结, 反之亦然, 值小, 只能说明相伴出现的机率较小, 并非一定是负联结。对于正联结, 值越大, 正联结性越强;对于负联结, 则值越小, 负联结性越强。因此, 以χ2检验为基础, 定性测定物种间联结性的显著性, 辅以PC、OI、DI和AC指数等方法, 共同分析紫茎泽兰群落中不同种对的种间关系, 效果较好, 能得出相对全面、准确的结论。
综合各项分析指标, 紫茎泽兰群落中不同种群间总体呈负相关, 但不显著。20个种对间表现出正相关性, 22个种对间则表现出负相关性, 或处于完全独立的状态。说明紫茎泽兰群落中的大部分物种间存在对水肥、光照等资源的生态位竞争, 并且这种竞争将随着资源的耗竭愈演愈烈。其中, 6个主要种对的负关联性较高, 这与紫茎泽兰的入侵特性一致。研究发现, 紫茎泽兰根系发达, 大肆掠夺土壤中水肥资源, 并通过分泌化感物质, 抑制侵入地其他种群的生长。且植株高大, 枝叶茂盛, 大大降低了其他种群得到光照的机率, 从而最终形成单一群落[27]。
种间联结性揭示的是种群竞争的结果或现状, 不能揭示竞争过程[28]。研究发现, 物种间的正联结性体现了生态学特征和生存环境的差异和互补性, 从而使各物种相互兼容[29]。紫茎泽兰与鬼针草、野艾蒿、马唐和灯笼草PC、OI和DI指数均较高, 一方面, 说明紫茎泽兰在一定时期内, 与鬼针草、野艾蒿、马唐和灯笼草具有相同或相似的生态位需求和对环境的适应性。另一方面, 其他原因也会导致此现象的产生。如在紫茎泽兰侵入之前, 鬼针草、野艾蒿、马唐和灯笼草等物种形成的群落存在生态缺陷, 可入侵性强, 导致紫茎泽兰得以顺利侵入此地, 并逐渐发展壮大。一定程度上也说明鬼针草、野艾蒿、马唐和灯笼草等物种对紫茎泽兰抑制其他物种生长特性的抵抗能力较强。
群落稳定性是群落中不同种群通过竞争、共生等方式自动调节, 从而达到动态平衡的集中反映[30]。研究表明, 在新物种入侵前, 原有群落处于相对稳定的平衡状态, 入侵后, 原有平衡迅速被打破, 群落稳定性降低[31]。紫茎泽兰入侵后, 由于其具有生命力强、根、茎、种子均可再生繁殖、根系分泌物显著抑制其他种群生长、养分吸收能力强等特点, 会在短时间内形成单优群落。因此, 紫茎泽兰入侵对本土植物群落有致命危害, 严重影响群落的稳定。本文调查结果进一步验证了这一点, 其入侵过程也是本土植物群落稳定性和生物多样性不断降低甚至丧失的过程。
种间联结性分析结果受取样数目和面积影响较大。取样数量越大, 测定结果越准确, 但所需人力物力投入也相应增加;数量过少则会导致调查结果失真。取样面积过大或过小会导致样方中出现的物种数偏多或偏少, 使得具有负联结的物种出现在同一样方中, 或具有一定正联结性的物种不能出现在同一样方中, 从而导致误判。因此, 选择合适的样方面积对准确测定种间联结性十分关键。笔者根据紫茎泽兰的实际生长状况, 人为规定了取样数目和面积, 可能得出的结论会有一定误差。
| [1] | Finch D M. Climate Change in Grasslands, Shrublands, and Deserts of the Interior American West:A Review and Needs Assessment. General Technical Report RMRS-GTR-285. Fort Collins, CO:USDA Forest Service:Rocky Mountain Research Station, 2012:48-59. |
| [2] | Weber E, Li B. Plant invasions in China:what is to be expected in the wake of economic development?. Bioscience, 2008, 58(5): 437–444. DOI:10.1641/B580511 |
| [3] | 万方浩, 郭建英, 王德辉. 中国外来入侵生物的危害与管理对策. 生物多样性, 2002, 10(1): 119–125. |
| [4] | 吴昊, 丁建清. 入侵生态学最新研究动态. 科学通报, 2014, 59(6): 438–448. |
| [5] | 吴虹玥, 包维楷, 王安. 外来种入侵对乡土生物多样性的影响机制. 长江流域资源与环境, 2004, 13(1): 40–46. |
| [6] | 于兴军. 紫茎泽兰入侵生态学研究[D]. 武汉: 武汉大学, 2005. http://cdmd.cnki.com.cn/Article/CDMD-10486-2006035333.htm |
| [7] | Haukisalmi V, Henttonen H. Analysing interspecific associations in parasites:alternative methods and effects of sampling heterogeneity. Oecologia, 1998, 116(4): 565–574. DOI:10.1007/s004420050622 |
| [8] | 郭志华, 卓正大, 陈洁, 吴梅凤. 庐山常绿阔叶、落叶阔叶混交林乔木种群种间联结性研究. 植物生态学报, 1997, 21(5): 424–432. |
| [9] | 王乃江, 张文辉, 陆元昌, 范少辉, 王勇. 陕西子午岭森林植物群落种间联结性. 生态学报, 2010, 30(1): 67–78. |
| [10] | 吴叶宽, 刘翔, 李隆云, 彭锐, 崔广林, 袁玲. 西南地区野生黄花蒿群落种间联结性分析. 中国中药杂志, 2013, 38(5): 670–673. |
| [11] | 张悦, 郭利平, 易雪梅, 曹伟, 王远遐, 吴培莉, 姬兰柱. 长白山北坡3个森林群落主要树种种间联结性. 生态学报, 2015, 35(1): 106–115. |
| [12] | 郭连金, 王涛. 空心莲子草入侵对乡土植物群落种间联结性及稳定性的影响. 中国生态农业学报, 2009, 17(5): 851–856. |
| [13] | 徐海根, 强胜. 中国外来入侵生物. 北京: 科学出版社, 2011. |
| [14] | 王亚麒, 焦玉洁, 陈丹梅, 袁玲, 黄玥, 吴叶宽, 杜如万. 紫茎泽兰浸提液对牧草种子发芽和幼苗生长的影响. 草业学报, 2016, 25(2): 150–159. DOI:10.11686/cyxb2015393 |
| [15] | 王洪炯, 何萍, 马家林. 紫茎泽兰传入凉山州草地调查研究报告. 中国草地, 1994, 16(1): 62–64. |
| [16] | 佘陆均, 覃志洪, 王同军. 凉山州山羊生产面临的问题及发展对策. 四川草原, 2000, 21(4): 50–52. |
| [17] | 杨紫美. 紫茎泽兰的危害及防除. 草业与畜牧, 2011(1): 32-32, 34-34. |
| [18] | 刘月莉. 四川甘洛铅锌矿区植被生态调查及重金属超富集植物的筛选[D]. 雅安: 四川农业大学, 2008. http://cdmd.cnki.com.cn/Article/CDMD-10626-2008198334.htm |
| [19] | Schluter D. A variance test for detecting species associations, with some example applications. Ecology, 1984, 65(3): 998–1005. DOI:10.2307/1938071 |
| [20] | 王伯荪. 植物种群学. 广州: 中山大学出版社, 1989: 115–120. |
| [21] | 王伯荪, 彭少麟. 南亚热带常绿阔叶林种间联结测定技术研究Ⅰ.种间联结测式的探讨与修正. 植物生态学与地植物学丛刊, 1985, 9(4): 274–285. |
| [22] | Whittaker R H. 植物群落排序. 王伯荪, 译. 北京: 科学出版社, 1986: 83-105. |
| [23] | Lu Z J, Ma K P. Spread of the exotic croftonweed (Eupatorium adenophorum) across southwest China along roads and streams. Weed Science, 2006, 54(6): 1068–1072. DOI:10.1614/WS-06-040R1.1 |
| [24] | Godron M, Daget P, Poissonet J, Poissonet P. Some aspects of heterogeneity in grasslands of Cantal//Patil G P, Pielou E C, Waters W E. Statistical Ecology. Volume 3:Many Species Populations, Ecosystems, and Systems Analysis. University Park, PA:Pennsylvania State University Press, 1971:397-415. |
| [25] | 黄祥童, 王绍先, 黄炳军, 尹航, 崔凯峰, 赵伟, 范宇光, 顾德峰. 珍稀植物对开蕨与其伴生物种的联结性及群落稳定性. 生态学报, 2015, 35(1): 80–90. |
| [26] | 刘金福, 洪伟, 樊后保, 林荣福. 天然格氏栲林乔木层种群种间关联性研究. 林业科学, 2001, 37(4): 117–123. DOI:10.11707/j.1001-7488.20010419 |
| [27] | 李会娜, 刘万学, 戴莲, 万方浩, 曹远银. 紫茎泽兰入侵对土壤微生物、酶活性及肥力的影响. 中国农业科学, 2009, 42(11): 3964–3971. DOI:10.3864/j.issn.0578-1752.2009.11.025 |
| [28] | 康冰, 刘世荣, 蔡道雄, 温远光, 史作民, 郭文福, 朱宏光, 张广军, 刘磊. 南亚热带人工杉木林灌木层物种组成及主要木本种间联结性. 生态学报, 2005, 25(9): 2173–2179. |
| [29] | YU X J, MA K P. Variation in reproductive characteristics of Eupatorium adenophorum populations in different habitats. Weed Research, 2006, 46(4): 319–326. DOI:10.1111/wre.2006.46.issue-4 |
| [30] | 张岗岗, 王得祥, 张明霞, 刘文桢, 郭小龙. 秦岭南坡松栎林群落演替过程中种间联结性和相关性研究. 西北植物学报, 2015, 35(8): 1657–1668. |
| [31] | 郭屹立, 卢训令, 丁圣彦. 伊洛河河岸带生态系统草本植物功能群划分. 生态学报, 2012, 32(14): 4434–4442. |