 2017, Vol. 37
2017, Vol. 37文章信息
- 刘莉, 韩美, 刘玉斌, 潘彬
- LIU Li, HAN Mei, LIU Yubin, PAN Bin.
- 黄河三角洲自然保护区湿地植被生物量空间分布及其影响因素
- Spatial distribution of wetland vegetation biomass and its influencing factors in the Yellow River Delta Nature Reserve
- 生态学报. 2017, 37(13): 4346-4355
- Acta Ecologica Sinica. 2017, 37(13): 4346-4355
- http://dx.doi.org/10.5846/stxb201508241763
-
文章历史
- 收稿日期: 2015-08-24
- 网络出版日期: 2017-02-23





湿地与森林、海洋并称为全球最具生产力的三大生态系统, 虽然与后两者相比湿地在全球景观生态系统中占有较小的比例, 但却起着无可替代的生态作用, 有“地球之肾”、“物种基因库”之称。湿地植被生物量是指单位面积内实存生活植被的有机质总量, 是衡量植被生产力水平的重要指标, 更是描述湿地生态系统物质循环和能量流动过程、评价湿地生态功能价值和湿地生态系统健康状况的重要基础数据。
利用遥感技术反演植被生物量并分析其空间分异规律, 不仅能快速直观地反映较大区域范围内植被的长势及其生存环境, 而且对于湿地生态系统的结构、功能、演替规律等都具有很好的指示作用[1-2]。因此, 近年来一直是生态学、地理科学、遥感科学等相关领域专家关注的热点问题。如Güneralp等利用多光谱遥感和非参数模型估算了河漫滩地区的地上生物量[3];李仁东等利用ETM数据与抽样数据进行Albers等面积投影及线性拟合, 估算了整个鄱阳湖的生物量[4];黎夏等根据雷达后向散射系数建立了红树林湿地植被生物量的估算模型[5];谭清梅等利用实测数据、ETM+图像数据, 建立一元线性、曲线回归模型、多元逐步回归模型, 研究了盐城湿地自然保护区核心区地上生物量干重、湿重[6];叶春, 吴桂平基于MODIS植被指数产品和同期植被生物量调查资料, 建立了遥感估算模型, 研究了鄱阳湖自然保护区湿地植被生物量的空间分布规律与模式[7-8]。上述学者构建的生物量模型为大区域生物量研究提供了便捷、高效的途径, 但由于采用的遥感数据源及建模的参数不尽相同, 所以其精度及广适性存在较大差异。另外, 关于生物量空间分异与环境因子关系方面的研究尚显薄弱, 有待加强。
黄河三角洲自然保护区地处黄河下游入海口处, 是以保护新生湿地生态系统和珍稀濒危鸟类为主的湿地类型自然保护区。保护区于1992年10月经国务院批准建立;1993年, 被中国人与生物圈国家委员会批准加入“中国人与生物圈自然保护区网络”;1996年, 被湿地国际亚太组织批准加入“东亚-澳大利西亚涉禽迁徙保护区网络”;2013年, 被国际湿地公约秘书处列入“国际重要湿地名录”[9]。该保护区处于河、海、陆交互地带, 长期受黄河水沙变化、尾闾摆动及海水入侵、波浪侵蚀等复杂多变因素的交互影响, 使其呈现生境类型多样、空间分异明显、新生湿地不断增长、生态系统脆弱等特点, 形成了其特定的植被生物量空间分布规律, 具有极高的科研价值[10]。但过去关于黄河三角洲湿地的研究大多集中在湿地动态变化[11-13]、湿地生态评价[14-18]、湿地生物多样性调查[19-20]及湿地生态修复[21-22]等方面。关于湿地植被生物量空间分布与环境因子关系的研究相对较少[23-24], 且多侧重于遥感反演模型的构建[25-28], 在模型构建中选择的遥感影像及参数都不尽相同, 反演对象多为单一物种[23-24, 27], 反演精度尚有待提高[29]。到目前为止尚未见关于黄河三角洲自然保护区湿地植被生物量空间分布及其与环境因子关系的相关论文发表。基于此, 本文以野外实测湿地植被地上生物量数据、Landsat-8影像数据和土壤各因子检测数据为基础, 通过分析各遥感因子与实测植被生物量之间的关系, 构建生物量反演模型, 计算湿地植被的地上生物量并生成相应的生物量空间分布图, 分析植被生物量的空间分布规律, 探究其与环境因子间的相互关系。该研究对于深化湿地形成、演化、发展研究, 更好地监控自然保护区湿地生态系统的变化, 保护新生湿地生态系统和珍稀濒危鸟类具有重要的理论和现实意义[30-31]。
1 数据获取与方法 1.1 研究区域概况黄河三角洲湿地保护区位于新、老黄河入海口两侧, 地理坐标为118°32.981′—119°20.450′E, 37°34.768′—38°12.310′N, 总面积15.3万hm2, 其中核心区面积59419 hm2, 缓冲区面积11233 hm2, 实验区面积82348 hm2。整个保护区分两部分, 分别为1996年到现行黄河入海口部分(即图 1中的北区)和1976到1996年黄河刁口河流路入海口部分(图 1中的南区)。

|
| 图 1 研究区位置、范围及样点分布图 Fig. 1 Location range and sample distribution map of the study area |
自然保护区独特的地理位置使其兼受河、海、陆交互作用, 造就了独特的生态系统特点, 成为研究河口湿地生态系统形成、演化、发展规律的重要基地。其特点表现在:(1) 新生性。在黄河径流泥沙和海洋动力共同作用下, 河口尾闾不断淤积延伸、摆动改道、循环演变, 新生湿地不断出现;(2) 脆弱性。湿地生态系统发育层次低, 处于物质和能量、结构和功能的非均衡状态, 缺乏自我调节能力, 抵抗外界干扰能力差;(3) 重要性。该自然保护区是东北亚内陆—环西太平洋和东亚—澳大利西亚两大鸟类迁徙路线的重要中转站、越冬地和繁殖地;(4) 多样性。自然保护区湿地生境类型多样, 有大面积的深水芦苇、浅滩水塘、岸边草丛、林地、农田、水产养殖塘、浅海水域、滩涂等, 为许多动植物, 特别是鸟类提供了适宜的繁殖地。保护区内物种资源丰富, 有淡水浮游植物8门、41科、97属, 291种;海洋浮游植物4门、116种;维管束植物46科、128属、195种。鱼类19目、58科、191种;两栖动物3科、6种;爬行动物6科, 10种;哺乳动物7目、15科、25种;鸟类19目、64科、367种[32]。(5) 空间分异性。多样的生境类型, 使湿地植被具有明显的空间分异性。在靠近黄河河岸的生境中, 由于淡水供应充足, 土壤盐分较低, 聚集了大量的中生和湿生植被, 植被盖度大。而在靠近海滩的区域, 由于地势低洼, 受海水影响强烈, 土壤盐度较高, 仅有少量耐盐植物存活。以下将通过植被生物量的空间差异来探讨湿地植被与环境因子的关系。
1.2 数据获取 1.2.1 遥感影像数据本文使用的遥感影像为2014年7月20日的Landsat-8 OLI数据, OLI陆地成像仪有9个波段, 空间分辨率为30米, 不仅包括了ETM+传感器所有的波段而且对原波段进行了调整, 新增了两个波段:第1波段蓝色波段(0.433—0.453 μm), 用于海岸带的观测, 第9波段短波红外波段(1.360—1.390 μm), 用于云检测。同时, OLI第5波段(0.845—0.885 μm)排除了0.825 μm处水汽吸收的影响, 第8波段的范围变窄, 植被和非植被在全色图像上反差加大, 能更好的对植被进行识别。鉴于影像具有这些特征, 满足本研究对植被指数的提取及对影像数据的要求。
1.2.2 样点布设及实验数据为获取湿地植被样方调查数据及土壤因子监测数据, 课题组于2014年8月、2015年8月进行了实地调查取样, 时值湿地植被生长旺盛的季节, 能够客观反映植被的生长情况, 并与本文所采用的遥感影像基本同期。本次共布设5条样带53个有效样点和样方, 样带分别按由海向陆、由黄河河道向其两侧布设(图 1), 目的是搞清湿地植被生物量由海向陆、由黄河河道向两侧的变化规律。草本植物样方设置为1 m×1 m, 在植被分布比较均匀的地方, 随机扔出1 m×1 m的PE胶管, 利用手持GPS定位仪记录样方中心位置的地理坐标, 统计样方内的植被类型、盖度、株高、株数等, 然后齐地收割植株地上部分, 立即称其鲜重, 取鲜重的1/3装入密封袋, 贴上标签, 带回实验室;每个样点去其表层受人为干扰的土壤, 挖取10 cm深度的土壤作为本次研究的土样, 装入塑料袋内贴上标签一并带回实验室。将带回的鲜植物放入80℃的恒温箱中持续烘8个小时后取出称重, 再次放入烘干箱中连续烘4小时, 取出后称重, 直到前后两次称重的误差小于1%时, 确定样方植被生物量干重。土壤是植物的载体, 土壤中的盐分、养分、水分条件直接影响着植物物种和生物量分布。鉴于此, 本研究对野外取回的53个土样的有机质、氮、磷、水溶性盐、pH值、土壤含水率进行检测, 检测方法均严格按照国家相关部门发布的行业标准进行, 有机质执行NY/T1121、氮执行LY/T1228-1999、磷执行HJ 632-2011、水溶性盐执行NY/T1121.16-2006、pH值执行NY/T 1377-2007、土壤含水率执行NY/T 52-1987。需要说明的是土壤含水率本是随机变化的指标, 但考虑到研究区土壤类型仅潮土和盐土两类, 受土壤结构影响, 一般情况下潮土的含水率总是大于盐土, 所以在本区该指标具有相对稳定性。
1.3 数据处理方法及流程 1.3.1 数据处理遥感解译是研究大区域植被生物量最先进、快捷的方法。本次采用2014年7月20日的Landsat-8 OLI影像, 因研究区地形平坦, 无需地形校正, 只进行辐射定标、FLAASH大气校正和图像不规则裁剪等处理, 提取的遥感信息为各波段的反射率、各植被指数。Landsat-8 OLI遥感影像共有11个波段, 其中1—6波段对湿地植被较为敏感, 因此选取了1—6波段进行各样点的反射率提取, 并选取差别较大的植被指数NDVI、EVI、MSAVI、DVI、RVI作为构建反演模型的指标[33], 各指标名称及计算公式[29]见表 1。
| 植被指数 Vegetation index | 中文全称 Full name in Chinese | 公式 Expression |
| NDVI(Normalized Difference Vegetation Index) | 归一化植被指数 |  |
| MSAVI(Modified Soil-adjusted Vegetation Index) | 修正土壤调整植被指数 | |
| EVI(Enhanced Vegetation Index) | 增强型植被指数 | |
| DVI(Difference Vegetation Index) | 差值植被指数 | |
| RVI(Ratio Vegetation Index) | 比值植被指数 | |
| ρNIR:近红外波段, ρRED:是红外波段, ρBLUE:蓝波段 | ||
多元线性回归模型是当两个或两个以上自变量与因变量之间是线性关系时所进行的回归分析[34]。本研究以多种植被指数作为自变量, 植被生物量作为因变量建立多元线性回归模型。设y为因变量(即本研究的生物量), x1, x2, …, xk为自变量(即本研究的植被指数), 并且自变量与因变量之间为线性关系时, 多元线性回归模型为:
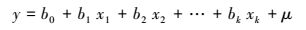
|
(1) |
式中, b0为常数项, b1, b2, …, bk为回归系数, μ为随机误差。
1.3.3 相关性分析相关性分析是研究随机变量之间相关关系的一种统计方法, 相关系数的大小表示变量之间的相关性强弱[33-34]。本研究利用相关系数对遥感因子与植被生物量进行分析。计算公式为:

|
(2) |
式中, R为相关系数, xi、yi为自变量与因变量在各样点的值, n为样点个数。
1.3.4 预测吻合度预测吻合度作为预测精度的一个衡量指标[35], 其值越大, 对应的预测模型精度越高, 可用于检验生物量模型的精度, 计算公式为:

|
(3) |
式中, G为预测吻合度, y(i)为第i个采样点实测值, 
平均残差系数是预测值与实测值相对误差系数的累积之和与观测次数的比值[36], 能很好的反映实测值与预测值的吻合度, 也可用于验证生物量模型的精度, 计算公式为:

|
(4) |
式中, y(i)为第i个采样点实测值, 
主要包括野外采样、实验室分析、遥感数据预处理、模型构建和湿地生物量反演等部分。其中野外采样包括土壤样品采集和地上植被信息采集, 经过实验室处理分析, 得到植被生物量干重及土壤盐分、养分、水分数据;遥感影像预处理包括辐射定标、大气校正、几何校正及影像裁剪, 然后提取植被指数;模型构建过程中先对各遥感因子与植被生物量干重进行相关性分析, 之后选取相关性大的遥感因子与43个样点的植被生物量建立一元线性、一元非线性和多元线性回归模型, 预留10个样点进行精度检验, 筛选出最优模型对植被生物量进行反演, 生成研究区湿地生物量空间分布图, 总体技术路线见图 2。

|
| 图 2 生物量建模流程图 Fig. 2 Biomass modeling flowchart |
在建立回归模型之前为提高反演拟合的精度将水域、道路等非植被覆盖区做了剔除处理, 之后用SPSS进行各遥感因子与植被生物量干重的相关性分析, 绘制遥感因子与植被生物量相关系数图(图 3), 从中选取相关性大的遥感因子与植被生物量建立一元线性、一元非线性、多元线性回归模型, 见表 2(一元线性、一元非线性最优模型及多元线性较优和最优模型)。

|
| 图 3 遥感因子与植被生物量相关系数图 Fig. 3 The correlation coefficient of remote sensing factors and vegetation biomass |
| 生物量反演模型 Biomass inversion model | 平均残差系数 Mean residual coefficient | 预测吻和度/%
Goodness of prediction |
| Y=207.27+1793.24h | 0.38 | 56.58 |
| Y=1/(1/u+0.004(2.38E-5)h) | 0.32 | 58.39 |
|
Y=30.57+30235.32a-29652.24b+4882.47c-5843.47d+1022.04e+1499.59f-1356.96g+
7463.64h-102.39i-1013.03k | 0.19 | 73.96 |
|
Y=-169.31+30793.11a-29758.18b+3898.13c-30995.09d+26912.74e+657.68f-2493.33h+
72.63i-1081.86k-13368.23j | 0.12 | 79.39 |
|
Y=-81.91+30538.38a-29666.04b+4199.67c-21054.01d+16722.20e+945.27f-553.00g+
1623.56h-1010.18k-8237.55j | 0.23 | 69.79 |
| Y=4.48+29535.70a-29230.29b+5292.28c-4284.89d-1260.95g+8842.50h-121.89i-885.93k-313.66j | 0.17 | 76.78 |
|
Y=56.70+29107.72a-28693.53b+5271.25c-5268.93d+1710.28f-1317.85g+9375.26h- 91.678i-897.31k-955.22j | 0.09 | 85.85 |
|
Y=-79.59+33345.82a-32536.93b+3802.82c-12204.05d+8802.64e+827.58f-1156.22g - 97.33i-1247.51k | 0.29 | 62.89 |
|
Y=117.37+28146.34a-29164.16b+5353.28c-4616.18d+1651.55e-1179.46g+10256.26h- 107.58i-3030.87j | 0.26 | 65.38 |
| 式中a=Band1, b=Band2, c=Band3, d= Band4, e= Band5, f= Band6, g=NDVI, h=DVI, i=RVI, j=MSAVI, k=EVI | ||
由表 2列出的一元线性、一元非线性最优模型及多元线性较优和最优模型可见, 在一元线性回归模型中, 以DVI为自变量建立的模型平均残差系数为0.38, 预测吻合度达56.58%, 模型最优;在一元非线性回归模型中, 以DVI为自变量建立的logistic曲线模型平均残差系数为0.32, 预测吻合度达58.39%, 模型最优;在多元线性回归模型中, 以NDVI、EVI、MSAVI、DVI、RVI、Band1、Band2、Band3、Band4、Band5作为自变量建立的模型, 其平均残差系数为0.09, 预测吻合度达85.85%, 模型精度较高, 模型最优。一元线性和一元非线性模型是基于单波段或单一植被指数对生物量进行的反演, 但在植被覆盖度不均匀时其精度和灵敏度会降低, 不能真实、准确的反映生物量信息。多元线性回归模型可将多个波段与多种植被光谱指数作为反演模型的指标, 能在一定程度上提高生物量估算的精度和可靠性[5, 37]。将表 2中的多元最优回归模型代入ENVI5.1中的Band Math进行计算, 生成植被生物量干重空间分布图并按自然间断点分类法(Natural Breaks)划分等级(表 3, 图 4,图 5)。由表 3可见, 生物量干重最低的1级区和最高的5级区面积小, 分别为82.23、72.16 km2, 各占整个湿地植被面积的13.35%、11.71%;较高的2、3、4级区所占面积较大, 为211.99、136.39、113.29 km2, 占34.41%、22.14%、18.39%;湿地生物量高值区集中于河道两侧及农田和林地周围的沟渠中, 低值区多分布于水分条件差, 土壤盐分高的滩涂、荒地区。通过ArcGIS的分类统计算出整个研究区湿地植被地上生物量干重约为454916.14 t。
| 分级 Classification | 植被生物量干重范围/(g/m2)
Vegetation dry weight | 面积/km2
Area | 占湿地植被面积比例/%
Proportion of wetland vegetation area |
| 1 | 0<B≤ 319 | 82.23 | 13.35 |
| 2 | 319<B≤ 502 | 211.99 | 34.41 |
| 3 | 502<B≤ 711 | 136.39 | 22.14 |
| 4 | 711<B≤ 1064 | 113.29 | 18.39 |
| 5 | 1064<B≤2698 | 72.16 | 11.71 |
| B:植被生物量干重 | |||

|
| 图 4 北部植被生物量干重空间分布图 Fig. 4 The space distribution of north wetland vegetation dry weight |

|
| 图 5 南部植被生物量干重空间分布图 Fig. 5 The space distribution of south wetland vegetation dry weight |
由于一元线性和一元非线性模型精度低, 所以仅对上述建立的多元线性回归模型进行验证, 结合建模时保留的10个样点, 采用预测吻合度与平均残差系数作为模型精度检验的指标, 其中平均残差系数越小, 预测吻合度越高, 表明模型精度越高, 预测结果越准确, 检验结果见表 4。
| 编号Number | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 备注Remarks |
| 实测值Measured value | 579.34 | 680.58 | 314.98 | 650.21 | 703.56 | 420.87 | 450.78 | 802.97 | 920.34 | 440.52 | 模型精度 |
| 预测值Predicted value | 611.56 | 531.35 | 379.66 | 637.92 | 687.50 | 494.84 | 416.94 | 779.35 | 1018.12 | 476.90 | 预测吻合度:85.85% |
| 残差Residual | -32.22 | 149.23 | -64.68 | 12.29 | 16.06 | -73.97 | 33.84 | 23.62 | -97.78 | -36.38 | 平均残差系数:0.09 |
从表 4可以看出, 生物量MLRM模型的平均残差系数为0.09, 预测吻合度为85.85%, 说明模型精度较高, 可用于本研究区进行生物量反演。
3 生物量分布规律及其影响因子分析 3.1 湿地植被生物量空间分布规律从图 4看出, 自然保护区刁口河入海口分区植被生物量干重呈现出南多北少, 东多西少的分布趋势。其中北部低值区分布在近海的淤泥质海滩, 南部高值区分布在远离海岸的灌丛沼泽和草本沼泽区, 东部高值区分布在生境改善恢复区内, 植被生长受到人类活动影响显著;西部低值区多分布在生态保育区内, 受人类活动影响较小。一千二管理站中的黄河故道, 植被生物量干重呈现由河道两岸向外侧递减的趋势。
从图 5看出, 黄河76a以前入海口区的生物量干重整体上呈现由海向陆增加的趋势, 除了黄河故道两侧为高值区外, 其余大面积为低值区。1976—1996年的黄河故道和96年后新生河道两侧的植被生物量都比较高, 其中前者的植被生物量略高于后者, 这与河道流路的早晚有关。综上, 植被生物量干重呈现由黄河河道向两侧、由陆向海递减的空间分布格局。
3.2 植被生物量空间分布的影响因素 3.2.1 株高对植被生物量的影响冯忠江[24]等指出, 黄河三角洲芦苇植株高度与水深成正相关, 并解释植被在进行光合作用时为了获得更多的CO2, 不断的透出水面增长高度, 因此可用芦苇高度的变化反映水深状况, 分析芦苇高度和芦苇生物量的相关性能间接反映水深与芦苇生物量的关系。
在SPSS中对芦苇、碱蓬生物量湿重、干重与植株高度的相关性进行分析, 得出芦苇湿重与植株高度的相关系数R=0.480, 芦苇干重与植株高度的相关系数R=0.782, 说明芦苇生物量与植株高度呈中度正相关关系, 水深对芦苇生物量的影响显著;碱蓬湿重与植株高度的相关系数R=0.278, 碱蓬干重与植株高度的相关系数R=0.193, 说明碱蓬生物量与植株高度呈轻度正相关, 线性关系较弱, 即碱蓬高度或者说水深对碱蓬生物量的贡献不是特别明显。
3.2.2 土壤因子对植被生物量的影响通过优势种植被芦苇和碱蓬的生物量干重与土壤各因子的相关性分析, 得到如表 5所示相关系数。
| 土壤因子 Soil factor | 有机质 Organic matter | 氮 Nitrogen | 磷 Phosphorus | 水溶性盐总量 Water soluble salts | 酸碱度 pH | 含水率 Rate of water content |
| 芦苇Phragmites australis | 0.087 | 0.127 | 0.023 | -0.159 | -0.159 | 0.387 |
| 碱蓬Suaeda glauca | 0.011 | 0.013 | 0.23 | -0.013 | 0.069 | -0.314 |
从表 5可以看出, 芦苇干重与水溶性盐总量、pH值成负相关, 与有机质、氮、磷、含水率成正相关。从相关系数上看, 芦苇生物量干重与磷的相关性最小, 与有机质、氮的相关性次之, 与土壤含水率的相关性最大。表明磷对芦苇生物量干重影响甚微, 有机质和氮影响较弱, 土壤含水率影响最大。保护区内的土壤为潮土和盐土, 潮土比盐土含水率高, 芦苇更适应在碱性弱、含水率高的潮土中生长。
碱蓬干重与土壤有机质、氮、磷和pH值成正相关, 而与水溶性盐总量和土壤含水率成负相关。碱蓬是强耐盐碱的植物, 因而碱蓬干重与pH值成正相关, 而与土壤含水率呈负相关。总体来看, 芦苇生物量干重和碱蓬生物量干重受土壤含水率影响最大, 土壤含水率对芦苇生长具有一定的促进作用, 对碱蓬生长具有一定的抑制作用。
4 结论(1) 黄河三角洲湿地保护区位于新、老黄河入海口两侧, 由于长期受黄河水沙变化、尾闾摆动及海水入侵、波浪侵蚀等复杂多变因素的交互影响, 使其呈现生境类型多样、空间分异明显、新生湿地不断增长、生态系统脆弱等特点, 形成了特定的植被生物量空间分布规律, 成为研究河口湿地生态系统形成、演化、发展规律的重要基地。
(2) 本文以野外实测湿地植被地上生物量数据、Landsat-8影像数据和土壤各因子检测数据为基础, 通过分析各遥感因子与实测植被生物量之间的关系, 构建生物量反演模型, 计算湿地植被的地上生物量并生成相应的生物量空间分布图, 分析植被生物量的空间分布规律, 探究其与环境因子间的相互关系。该研究对于深化湿地形成、演化、发展研究, 更好地监控自然保护区湿地生态系统的变化, 保护新生湿地生态系统和珍稀濒危鸟类具有重要的理论和现实意义。
(3) 运用一元线性、一元非线性和多元线性回归建模, 分析了判定系数大小、模型精度, 得出以NDVI、EVI、MSAVI、DVI、RVI、Band1、Band2、Band3、Band4、Band6为自变量, 以植被生物量干重为因变量建立的10元线性回归模型判定系数最大, 拟合精度最高。
因为一元线性和一元非线性模型是基于单波段或单一植被指数对生物量进行的反演, 此两种方法在植被覆盖度不均匀时其精度和灵敏度会降低, 不能真实、准确的反映生物量信息。多元线性回归模型是将多个波段与多种植被光谱指数作为反演模型的指标, 能在一定程度上提高生物量估算的精度和可靠性。同时, 本文在建立回归模型之前将水域、道路等非植被覆盖区做了剔除处理, 也在一定程度上提高了估算精度。
(4) 研究区湿地植被地上生物量干重约为454916.14 t, 因分布不均, 自低到高划为5级, 最低的1级区和最高的5级区面积小, 为82.23、72.16 km2, 分别占整个湿地植被面积的13.35%、11.71%;较高的2、3、4级区所占面积较大, 为211.99、136.39、113.29 km2, 占34.41%、22.14%、18.39%;高值区集中在河道、沟渠两侧, 低值区分布于水分条件差, 土壤盐分高的滩涂、荒地区。这是因为河道、沟渠两侧的生境中, 淡水供应充足, 土壤盐分较低, 聚集了大量的中生和湿生植被, 植被盖度大。而在靠近海滩的区域, 由于地势低洼, 受海水影响强烈, 土壤盐度较高, 仅有少量耐盐植物存活。
(5) 研究区湿地植被生物量干重呈现由陆向海、由黄河河道向两侧递减的趋势。植被生物量干重的空间分布格局与各环境因子有很大的关系, 水深对优势种芦苇生物量干重影响最大, 土壤含水率对优势种碱蓬生物量干重影响最大。水、盐条件是导致优势种植被生物量干重出现空间分异的主导因素。
| [1] | 王树功, 黎夏, 周永章. 湿地植被生物量测算方法研究进展. 地理与地理信息科学, 2004, 20(5): 104–109. |
| [2] | 郝文芳, 陈存根, 梁宗锁, 马丽. 植被生物量的研究进展. 西北农林科技大学学报:自然科学版, 2008, 36(2): 175–182. |
| [3] | Güneralp İ, Filippi A M, Randall J. Estimation of floodplain aboveground biomass using multispectral remote sensing and nonparametric modeling. International Journal of Applied Earth Observation and Geoinformation, 2014, 33: 119–126. DOI:10.1016/j.jag.2014.05.004 |
| [4] | 李仁东, 刘纪远. 应用Landsat ETM数据估算鄱阳湖湿生植被生物量. 地理学报, 2001, 56(5): 532–540. DOI:10.11821/xb200105004 |
| [5] | 黎夏, 叶嘉安, 王树功, 刘凯, 刘小平, 钱峻屏, 陈晓越, 何执兼, 覃朝锋. 红树林湿地植被生物量的雷达遥感估算. 遥感学报, 2006, 10(3): 387–396. |
| [6] | 谭清梅, 刘红玉, 张华兵, 王聪, 侯明行. 盐城海滨湿地植被地上生物量遥感估算研究. 自然资源学报, 2013, 28(12): 2044–2055. DOI:10.11849/zrzyxb.2013.12.003 |
| [7] | 叶春, 赵晓松, 吴桂平, 王晓龙, 刘元波. 鄱阳湖自然保护区植被生物量时空变化及水位影响. 湖泊科学, 2013, 25(5): 707–714. DOI:10.18307/2013.0512 |
| [8] | 吴桂平, 叶春, 刘元波. 鄱阳湖自然保护区湿地植被生物量空间分布规律. 生态学报, 2015, 35(2): 361–369. |
| [9] | 张德静, 东野广成, 宋丽丽, 韩朝晖. 山东黄河三角洲自然保护区现状及保护对策. 林业科技管理, 2004(3): 31–32. |
| [10] | 崔保山, 杨志峰. 湿地生态系统健康的时空尺度特征. 应用生态学报, 2003, 14(1): 121–125. |
| [11] | 李秀娟, 赵庚星, 刘洪义, 栾新诚. 黄河清水沟改道以来河口新生湿地动态监测研究. 自然资源学报, 2006, 21(2): 328–332. |
| [12] | 崔保山, 刘兴土. 黄河三角洲湿地生态特征变化及可持续性管理对策. 地理科学, 2001, 21(3): 250–256. |
| [13] | 叶庆华, 刘高焕, 姚一鸣, RussiM. 黄河三角洲新生湿地土地利用变化图谱. 地理科学进展, 2003, 22(2): 141–148. DOI:10.11820/dlkxjz.2003.02.005 |
| [14] | 崔保山, 李英华, 杨志峰. 基于管理目标的黄河三角洲湿地生态需水量. 生态学报, 2005, 25(3): 606–614. |
| [15] | 许学工, 林辉平, 付在毅, 布仁仓. 黄河三角洲湿地区域生态风险评价. 北京大学学报:自然科学版, 2001, 37(1): 111–120. |
| [16] | 韩美, 李艳红, 李海亭, 庞小平. 山东寿光沿海湿地生态系统健康诊断. 中国人口·资源与环境, 2006, 16(4): 78–83. |
| [17] | 韩美, 张晓慧. 黄河三角洲湿地主导生态服务功能价值估算. 中国人口·资源与环境, 2009, 19(6): 37–43. |
| [18] | Han M, Cui J L, Hao Z, Wang Y, Wang R Q. Eco-compensation of wetlands in Yellow River delta of Shandong Province, China. Chinese Geographical Science, 2012, 22(1): 119–126. DOI:10.1007/s11769-011-0501-1 |
| [19] | 夏江宝, 李传荣, 许景伟, 刘立杰. 黄河三角洲滩涂湿地夏季大型底栖动物多样性分析. 湿地科学, 2009, 7(4): 299–305. |
| [20] | 李峰, 谢永宏, 陈心胜, 邓正苗, 李旭. 黄河三角洲湿地水生植物组成及生态位. 生态学报, 2009, 29(11): 6257–6265. DOI:10.3321/j.issn:1000-0933.2009.11.061 |
| [21] | 单凯. 黄河三角洲自然保护区湿地生态恢复的原理、方法与实践. 湿地科学与管理, 2007, 3(4): 16–20. |
| [22] | 何智娟, 黄锦辉, 潘轶敏, 张效艳. 黄河流域生态系统特征及下游生态修复实践. 环境与可持续发展, 2010, 35(4): 9–13. |
| [23] | 牟晓杰, 孙志高, 刘兴土. 黄河口滨岸潮滩不同生境下翅碱蓬生物量空间分形特征与磷营养动态. 草业学报, 2012, 21(3): 45–53. DOI:10.11686/cyxb20120306 |
| [24] | 冯忠江, 赵欣胜. 黄河三角洲芦苇生物量空间变化环境解释. 水土保持研究, 2008, 15(3): 170–174. |
| [25] | 丁蕾, 马毅. 基于现场光谱的黄河口湿地芦苇生物量估算模型研究. 海洋环境科学, 2015, 34(5): 718-722, 728-728. |
| [26] | 吴天君, 张曦文, 赫晓慧. 基于CBERS的黄河湿地生物量反演研究. 测绘与空间地理信息, 2012, 35(5): 18-19, 27-27. |
| [27] | 傅新, 刘高焕, 黄翀, 刘庆生. 湿地翅碱蓬生物量遥感估算模型. 生态学报, 2012, 32(17): 5355–5362. |
| [28] | 王建步, 张杰, 马毅, 任广波. 基于高分一号WFV卫星影像的黄河口湿地草本植被生物量估算模型研究. 激光生物学报, 2014, 23(6): 604–608. |
| [29] | 高明亮, 赵文吉, 宫兆宁, 赫晓慧. 基于环境卫星数据的黄河湿地植被生物量反演研究. 生态学报, 2013, 33(2): 542–553. |
| [30] | Krebs C J. Ecology:The Experimental Analysis of Distribution and Abundance.5th ed. San Francisco: Benjamin Cummings, 2001: 513–536. |
| [31] | 蓝振江, 蔡红霞, 曾涛, 王硕果, 郭海燕, 曾宗永. 九寨沟主要植物群落生物量的空间分布. 应用与环境生物学报, 2004, 10(3): 299–306. |
| [32] | 宋家敬, 刘月良, 朱书玉, 张俊, 陆峰. 山东黄河三角洲国家级自然保护区详细规划(2014-2020年). 山东黄河三角洲国家级自然保护区管理局, 2014: 47–50. |
| [33] | 杨存建, 刘纪远, 黄河, 许辉熙, 党承林. 热带森林植被生物量与遥感地学数据之间的相关性分析. 地理研究, 2005, 24(3): 473–479. |
| [34] | 张景阳, 潘光友. 多元线性回归与BP神经网络预测模型对比与运用研究. 昆明理工大学学报:自然科学版, 2013, 38(6): 61–67. |
| [35] | 沈掌泉, 周斌, 孔繁胜, BaileyJ S. 应用广义回归神经网络进行土壤空间变异研究. 土壤学报, 2004, 41(3): 471–475. DOI:10.11766/trxb200303310323 |
| [36] | 王江浩, 葛咏. 遥感影像几何校正的GCP残差模拟分析. 遥感技术与应用, 2011, 26(2): 226–232. DOI:10.11873/j.issn.1004-0323.2011.2.226 |
| [37] | 邢丽玮, 李小娟, 李昂晟, 周德民. 基于高光谱与多光谱植被指数的洪河沼泽植被叶面积指数估算模型对比研究. 湿地科学, 2013, 11(3): 313–319. |