 2016, Vol. 36
2016, Vol. 36文章信息
- 张晓可, 于道平, 王慧丽, 万安, 蒋泽球, 陈寿文
- ZHANG Xiaoke, YU Daoping, WANG Huili, WAN An, JIANG Zeqiu, CHEN Shouwen
- 长江安庆段江豚主要栖息地鱼类群落结构
- Fish community structure in main habitat of the finless porpoise, the Anqing section of Yangtze River
- 生态学报, 2016, 36(7): 1832-1839
- Acta Ecologica Sinica, 2016, 36(7): 1832-1839
- http://dx.doi.org/10.5846/stxb201409251904
-
文章历史
- 收稿日期: 2014-09-25
- 网络出版日期: 2015-08-05






2. 安庆师范学院资源与环境学院, 安庆 246011;
3. 安庆市渔业局, 安庆 246011
2. College of Resource and Environment, Anqing Normal University, Anqing 246011, China;
3. Fishery Bureau of Anqing, Anqing 246011, China
长江江豚(Neophocaena asiaeorientalis asiaeorientalis)是窄鳍江豚的一种,仅分布于我国长江中下游和两个大型通江湖泊-洞庭湖和鄱阳湖[1]。然而,由于人类活动的严重干扰(如江湖阻隔、航运、非法渔业、采砂等),长江豚类栖息地正在逐渐恶化和丧失[2, 3, 4]。其种群数量已由2006年的1800头下降至2012年的1040头,并且未来10年内该物种也或将走向灭绝[4]。因此加强长江江豚的保护已经刻不容缓。在自然情况下,江豚主要捕食小型鱼类,同时也摄食一些虾类[5]。随着长江渔业资源的逐步枯竭,食物因素已经成为影响江豚生存最重要的因素[6, 7]。然而,关于江豚主要栖息地鱼类群落结构的研究较少,难以为江豚的原地和迁地保护工作提供科学依据。
在长江干流中,安庆江段是长江江豚密度最高的区域[2, 4]。在对安庆江段的多次野外调查中均发现,长江与皖河交汇处(皖河口)有较大规模江豚分布,是江豚最重要的栖息地之一[8]。为了加强江豚的保护工作,安庆市政府于2007年在安庆江段建立长江豚类自然保护区,并在皖河口这一重点水域设立江豚监测站,对江豚数量进行长期观察。因此,本研究选择皖河口及其附近水域作为研究区域,对其鱼类群落进行了周年调查,并对比了皖河口及其附近水域鱼类群落结构的差异及其动态变化,相关研究结果对鱼类资源的恢复和江豚的保护均具有重要意义。
1 材料与方法 1.1 研究区域概况皖河发源于大别山南麓,位于长江下游北岸安徽省境内。皖河全长227 km,年均径流量53.79亿m3。皖河上游有三条主要支流,由西向东分别为长河、潜水和皖水,三者汇合后形成皖河干流(图 1)。随着流域内经济的发展,皖河的航运功能越来越重要。政府部门曾对皖河进行多次河道整治,目前从皖河口向上41.2 km为三级航道,可供300吨级船舶航行。

|
| 图 1 皖河口及其附近水域鱼类采样断面 Fig.1 Fish sampling transects in estuary of the Wanhe River and its adjacent waters |
皖河口位于安庆市西,宽度约130 m。为了航运需求,曾对皖河口进行多次清淤开挖,目前河底平均高程2.21 m,年均水位9.22 m,可供500吨级船舶出入。受长江水位变化的影响,皖河口水位波动剧烈,2013—2014年调查期间最大水深14.4 m,最小水深4.6 m,平均水深8.5 m。
1.2 采样方法于2013年9月—2014年8月逐月对皖河口及其附近水域(皖河口上下1.5 km)鱼类群落进行调查。在长江北岸共设置5个采样断面(断面1和5于11月份开始调查),每个断面长度为400 m,断面之间间隔为200 m(图 1)。除断面3外,断面1、2、4和5均为缓坡,并且流速差异不大。主要采用电拖网和复合刺网相结合的方式进行采集。前者主要用于采集中下层和底层鱼类,所用电拖网电压为12 V,长3 m,宽1 m,网目为10 mm。每个断面在长江北岸顺着水流的方向电鱼400 m,然后收集所有捕获鱼类。后者主要用于采集中上层鱼类,长度为100 m,宽度为2 m;网目有3种规格,分别为10、20、40 mm。每个断面顺着水流的方向漂流10分钟,然后收集捕获的鱼类。渔获物带回实验室后立即参照相关资料进行鉴定[9, 10],并记录所有鱼类的体长和体重。体重精确到0.1 g,体长精确到1 mm。
1.3 数据分析根据已发表文献[9, 11, 12],本研究将初次性成熟小于2龄,最大体长小于24 cm的鱼类定义为小型鱼类。根据鱼类食性的不同,将其分为肉食性、植食性、杂食性、浮游生物食性和碎屑食性鱼类[11, 12]。根据垂直空间分布的不同,将其分为中上层、中下层和底层鱼类[13]。计算每个断面不同鱼类优势度,计算公式为[14]:
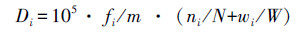
式中,Di为第i种鱼的优势度;m为取样次数;fi为第i种鱼在m次取样中出现频数;ni、wi分别为第i种鱼在m次取样中的个体数和重量;N、W分别为m次取样总个体数和总重量。
本研究中将优势度大于5000的种定为优势种[13]。不同断面渔获物的数量、重量和种类数以及中上层、中下层和底层渔获物数量、重量和种类数的差异采用单因素方差分析(ANOVA,One-way analysis of variance)来进行检验,如果有显著差异,采用Tukey多重比较来检验组之间的差异。以上所有统计分析均在SPSS 13.0中进行。
2 研究结果 2.1 种类组成共采集鱼类35种,隶属于10科26属(表 1)。鲤科鱼类占优势,共有16属20种,占总种数的57.1%;其次是鲿科和鮨科,分别为4种(11.4%)和3种(8.6%);其他各科种数都较少,共占总种数的22.9%。小型鱼类共19种,占总种数的54.3%。5个断面中,皖河口(断面3)鱼类种数最多,为27种;断面1、2、4和5分别为16、20、19和14种(表 1)。
| 物种 Species | 生态类型 Ecological guilds | 断面1 Transect 1 | 断面2 Transect 2 | 断面3 Transect 3 | 断面4 Transect 4 | 断面5 Transect 5 | |
| 银鱼科Salangidae | |||||||
| 大银鱼Protosalanx hyalocranius* | C | U | 70 | 17 | 84 | ||
| 鲤科Cyprinidae | |||||||
| 鲤Cyprinus carpio | O | De | 4500 | 18 | 1946 | ||
| 鲫Carassius auratus | O | De | 591 | 339 | 484 | 1255 | 1002 |
| 似鳊Pseudobrama simony* | P | L | 58 | 22 | 68.7 | 130 | |
| 鳊Parabramis pekinensis | H | L | 623 | ||||
| 油餐Hemiculter bleekeri* | P | U | 3301 | 5602 | 48455 | 3370 | 11885 |
| 餐Hemiculter leucisculus* | P | U | 1317 | 7045 | 36990 | 2804 | 7214 |
| 寡鳞银飘Pseudolaubuca engraulis | O | U | 24 | ||||
| 黄尾鲴Xenocypris davidi | H | L | 48 | ||||
| 银鲴Xenocypris argentea | H | De | 18 | 61 | 127 | ||
| 圆吻鲴Distoechodon tumirostris | H | L | 157 | 116 | |||
| 长蛇鮈Saurogobio dumerili* | O | De | 321 | 29 | 266 | ||
| 蛇鮈Saurogobio dabryi* | O | De | 9097 | 8883 | 1357 | 2851 | 1362 |
| 吻鮈Rhinogobio typus* | O | De | 14 | ||||
| 兴凯鱊Acheilognathus chankaensis* | D | U | 70 | ||||
| 黑鳍鳈Sarcocheilichtys nigripinnis* | O | De | 36 | ||||
| 铜鱼Coreius heterodon | O | De | 1265 | 176 | |||
| 麦穗鱼Pseudorasbora parva* | O | L | 99 | ||||
| 棒花鱼Abbottina rivularis* | O | De | 38 | ||||
| 鲢Hypophthalmichthys molitrix | P | U | 92 | ||||
| 鳙Hypophthalmichthys nobilis | P | U | 22 | ||||
| 鳅科Cobitidae | |||||||
| 泥鳅Misgurnus anguillicaudatus* | O | De | 13 | ||||
| 鲇科Siluridae | |||||||
| 鲇Silurus asotus | C | De | 3138 | 286 | 328 | 1346 | 1188 |
| 鲿科Bagridae | |||||||
| 黄颡鱼Pelteobagrus fulvidraco* | O | De | 5419 | 11461 | 4906 | 27269 | 18665 |
| 长须黄颡鱼Pelteobagrus eupogon* | O | De | 5985 | 14478 | 6930 | 17785 | 18044 |
| 光泽黄颡鱼Pelteobagrus nitidus* | O | De | 2395 | 3449 | 1733 | 606 | 87 |
| 瓦氏黄颡鱼Pelteobagrus vachelli | O | De | 1610 | 3583 | 1873 | 4330 | 69 |
| 鱵科Hemirhamphidae | |||||||
| 间下鱵Hyporhamphus intermedius* | C | U | 688 | 177 | 281 | 409 | |
| 鲱科Clupeidae | |||||||
| 短颌鲚Coilia brachygnathus | C | U | 9068 | 7903 | 12647 | 8814 | 6687 |
| 鮨科Serranidae | |||||||
| 鳜Siniperca chuatsi | C | L | 52 | 3124 | 246 | 25 | |
| 斑鳜Siniperca scherzeri | C | L | 159 | 62 | 20 | ||
| 大眼鳜Siniperca kneri | C | L | 72 | ||||
| 塘鳢科Eleotridae | |||||||
| 沙鳢Odontoburis obscura* | C | De | 12 | ||||
| 黄黝Hypseleotris swinhonis* | O | De | 70 | ||||
| 攀鲈科Anabantidae | |||||||
| 圆尾斗鱼Macropodus ocellatus* | C | L | 10 | ||||
| *小型鱼类Small fish;O杂食性 Omnivore;P浮游生物食性Planktivore;H草食性 Herbivore; De碎屑食性Detritivore;C肉食性Carnivore;U中上层Upper;L中下层Lower;D底栖性Demersal | |||||||
各断面均以小型鱼类为主,且占总种数50%以上(图 2)。从食性上来看,各断面均以杂食性鱼类种数最多,而碎屑食性鱼类种数最少,并且皖河口杂食性和浮游生物食性鱼类种数明显高于其他断面(图 2)。从垂直分布来看,各断面均以底层鱼类种数最多,而中下层鱼类种数最少,并且中上层、中下层和底层最大值均出现在皖河口(图 2)。

|
| 图 2 皖河口及其附近断面各生态类型鱼类对比 Fig.2 Comparison of different ecological guilds in estuary of the Wanhe River and its adjacent areas |
方差分析结果表明,皖河口渔获物总数量和总重量与其他断面无显著差异,但总种数却显著高于断面1和断面5(表 2)。就中上层、中下层和底层鱼类而言,5个断面的显著差异仅存在于中上层鱼类,且皖河口中上层鱼类数量和重量均显著高于其他断面,而种数仅显著高于断面1和5(表 2)。
| 变量 Variables | 断面1 Transect 1 | 断面2 Transect 2 | 断面3 Transect 3 | 断面4 Transect 4 | 断面5 Transect 5 | F | P |
| 总种数Total species number | 3.1±0.5a | 4.8±0.7ab | 5.5±0.4b | 4.7±0.6ab | 2.7±0.4a | 4.374 | 0.003 |
| 中上层鱼类种数Species number of upper fish | 0.9±0.3a | 1.6±0.2ab | 2.5±0.2b | 1.7±0.3ab | 1.0±0.2a | 6.286 | 0.000 |
| 中下层鱼类种数Species number of lower fish | 0.3±0.2 | 0.4±0.2 | 0.4±0.1 | 0.2±0.1 | 0.3±0.1 | 0.955 | 0.436 |
| 底层鱼类种数Species number of demersal fish | 1.9±0.36 | 2.8±0.5 | 2.7±0.4 | 2.8±0.4 | 1.6±0.3 | 1.610 | 0.179 |
| 渔获物总数量Total individual number of catch | 11.4±3.5 | 33.0±7.2 | 35.0±7.7 | 29.7±12.4 | 8.5±2.6 | 2.148 | 0.082 |
| 中上层渔获物数量Individual number of upper fish | 5.1±2.4a | 8.1±1.8a | 22.5±6.1b | 6.5±1.7a | 4.0±2.43a | 4.925 | 0.001 |
| 中下层渔获物数量Individual number of lower fish | 0.3±0.2 | 1.6±0.7 | 0.6±0.2 | 0.2±0.1 | 0.1±0.1 | 2.404 | 0.056 |
| 底层渔获物数量Individual number of demersal fish | 6.0±2.6 | 23.3±7.1 | 12.3±5.3 | 23.0±12.5 | 4.4±1.3 | 1.320 | 0.269 |
| 渔获物总重量Total weight of catch | 193.3±96.5 | 390.8±107.1 | 328.0±81.7 | 376.4±171.8 | 99.2±32.7 | 1.123 | 0.351 |
| 中上层渔获物重量Weight of upper fish | 38.4±18.5a | 76.8±19.7a | 255.7±79.5b | 47.4±10.1a | 42.4±27.7a | 5.092 | 0.001 |
| 中下层渔获物重量Weight of lower fish | 21.7±20.3 | 52.0±29.0 | 9.3±3.8 | 3.9±1.9 | 2.5±1.7 | 1.679 | 0.162 |
| 底层渔获物重量Weight of demersal fish | 133.3±90.5 | 262.0±86.9 | 63.0±18.5 | 325.0±171.8 | 54.4±14.3 | 1.438 | 0.228 |
| 不同的上标字母表示统计学上有显著差异(P < 0.05) | |||||||
不同断面鱼类优势度见表 1。从表中可以看出,不同断面主要优势种差异较大。断面1主要优势种为蛇鮈(Saurogobio dabryi)和短颌鲚(Coilia brachygnathus),其次是黄颡鱼(Pelteobagrus fulvidraco)和长须黄颡鱼(Pelteobagrus eupogon)。断面2、4、5中,主要优势种均为底层鱼类黄颡鱼和长须黄颡鱼,而皖河口主要优势种却为中上层鱼类餐(Hemiculter leucisculus)、油餐(Hemiculter bleekeri)和短颌鲚。
2.5 季节动态不同季节各断面渔获物的总重量和总数量如图 3所示。一般秋、冬季节这两个指标要明显高于春、夏季节,且最低值均出现在夏季。断面1和断面5于11月份才开始采集,因此会对秋季渔获物的总重量和总数量造成一定影响。

|
| 图 3 皖河口及其附近水域不同季节渔获物重量和数量(平均值) Fig.3 Weight and individual number of catch in estuary of the Wanhe River and its adjacent waters (average values) |
安庆江段上起宿松县汇口镇,下至枞阳县老洲镇,全长243 km。沿岸有许多湖泊和支流汇入长江,再加上河道弯曲,在水力条件的作用下形成了十分多样的栖息地环境,鱼类资源十分丰富。然而,由于人类活动的过度干扰,该江段鱼类种类组成已发生明显变化。本研究皖河口及其附近水域共调查到鱼类35种,与历史资料相比,明显低于20世纪90年代的63种[15]和2002—2004年的46种[16]。一些常见大型经济鱼类,如草鱼(Ctenopharyngodon idellus)、青鱼(Mylopharyngodon piceus)、鳡(Elopichthys bambusa)和赤眼鳟(Squaliobarbus curriculus)等均未采集到。一些洄游性鱼类(如长颌鲚Coilia macrognathos、鳗鲡Anguilla japonica)和河口鱼类(如三线舌鳎Cynoglossus trigrammus、鲻鱼Mugil cephalus)在2002—2004年的调查中还少量存在,本研究中已完全消失。除采样工具和努力强度不一致造成的差异外,安庆江段无节制的过度捕捞可能是导致鱼类物种多样性下降的主要原因。此外,和长江其他区域鱼类种类组成特点一致[17, 18],皖河口及其附近水域也主要是以鲤科鱼类为主,并且表现出严重的小型化趋势。与2002—2004年调查结果相比[16],小型鱼类比例已经由45.7%上升至54.3%。本研究中两种小型鱼类黄黝(Hypseleotris swinhonis)和圆尾斗鱼(Macropodus ocellatus)在2002—2004年调查中均未采集到。
3.2 不同断面鱼类群落特征5个断面中,皖河口鱼类多样性最高,这可能主要是由于皖河口鱼类栖息地更为多样。除支流外,在交汇处上方一侧存在一个水流很小甚至停滞的区域为滞留区,在交汇处下方一侧存在一个水流较缓的回水区为分离区[19]。较缓的流速使有机物在这些区域沉积,使浮游动物和底栖动物资源较为丰富,从而为鱼类提供丰富的食物来源。从鱼类食性上来看,各断面均以杂食性鱼类为主。杂食性鱼类即可摄食动物也可摄食植物,相比其他食性鱼类生态位更宽,因此对环境的适应能力更强。对黄河、西江等其他大型河流鱼类食性的分析也有类似结果[13, 20]。此外,皖河口浮游生物食性鱼类种数明显高于其他断面。这主要是由于该断面泥沙含量低、营养物质丰富、浮游生物现存量较高造成的。从垂直分布来看,皖河口中上层、中下层和底层鱼类种数均明显高于其他断面,这说明皖河口的河道开挖对鱼类多样性影响较小,但其对优势种的种类组成却影响较大。已有许多研究表明,河道挖掘会对鱼类的群落结构产生显著影响[21, 22]。本研究中皖河口的主要优势种已经由黄颡鱼和长须黄颡鱼等底层鱼类转变为餐和油餐等中上层鱼类,并且中上层鱼类的数量和重量均显著高于其他断面。此外,中上层鱼类餐和油餐主要捕食浮游生物,皖河口较为丰富的食物资源也为其优势地位的建立提供了良好条件。不同断面之间渔获物总数量、总重量以及中下层和底层鱼类的种数、数量和重量差异并不显著,这可能也是由于皖河口的挖掘造成的,因为皖河口较深的水深可能不利于中下层和底层鱼类的栖息,但对中上层鱼类影响却较小。
就不同季节而言,秋、冬季节较低的水位使鱼类活动范围缩小,且更便于采集,因此渔获物总数量和总重量都较大;相反,夏季高水位时期许多鱼类进入附近河岸带草丛中摄食或产卵,使电网和刺网捕获的难度加大,渔获物总数量和总重量也都较小。此外,为了更好的比较江豚不同活动区域鱼类群落的变化,本研究采样断面设置相对较近,这可能会对各断面鱼类群落的差异造成一定影响,在以后的研究中还需进一步验证。
3.3 鱼类群落与江豚的关系在野外环境下江豚通常选择特定水域栖息,如江河、江湖交汇处、沙洲附近等。这些水域较为丰富的鱼类资源被认为是其活动的主要原因[8, 23]。然而,本研究中不同断面渔获物的数量和重量并没有显著差异,并且不同空间鱼类的差异也仅存在于中上层鱼类,这说明皖河口江豚较大规模分布主要是由于其更为丰富的中上层鱼类资源。已有研究表明,江豚在野外环境中更容易捕食中上层鱼类,而这可能主要与江豚的形态结构有关[24]。在长期的进化过程中,白鱀豚和江豚生态位已经产生了分化,前者吻细长且着生有齿,可能更利于捕食底层鱼类;而后者吻短,更利于捕食中上层鱼类。虽然江豚也可摄食部分中下层和底层鱼类,但无疑要消耗更多的能量。江豚作为一种小型鲸类动物,能量储备并不高[5, 25]。在食物资源极其有限的情况下,用最少的能量消耗去获得尽可能多的食物对江豚是极为重要的。
本研究结果对长江江豚的保护具有重要作用。目前长江干流豚类仅剩500多头[4],然而经济发展与物种保护的尖锐矛盾使长江豚类的威胁因素一直难以消除。随着长江鱼类资源的日渐枯竭,江豚必然会花费更多的能量用于捕食。对于江豚这种濒危鲸类来说,少量个体的丧失可能会重创整个种群的维持和发展[23]。因此,迁地保护工作已在长江中下游许多长江故道中展开(如安庆西江、监利何王庙等)。在评估这种半自然水体是否适合江豚生存时,应该重点关注中上层渔获物的数量和重量。在向这种半自然水体中补充江豚食物资源时,也应该尽量增加中上层鱼类。同时,在江豚的原地保护工作中,也应该对中上层鱼类丰富的水域重点监测和保护,并在豚类保护核心区划定时作为优先考虑的区域。
| [1] | 高安利, 周开亚. 中国水域江豚外形的地理变异和江豚的三亚种. 兽类学报, 1995, 15(2): 81-92. |
| [2] | Zhao X J, Barlow J, Taylor B L, Pitman R L, Wang K X, Wei Z, Stewart B S, Turvey S T, Akamatsu T, Reeves R R, Wang D. Abundance and conservation status of the Yangtze finless porpoise in the Yangtze River, China. Biological Conservation, 2008, 141(12): 3006-3018. |
| [3] | Wang D. Population status, threats and conservation of the Yangtze finless porpoise. Chinese Science Bulletin, 2009, 54(19): 3473-3484. |
| [4] | Mei Z G, Zhang X Q, Huang S L, Zhao X J, Hao Y J, Zhang L, Qian Z Y, Zheng J S, Wang K X, Wang D. The Yangtze finless porpoise: On an accelerating path to extinction? Biological Conservation, 2014, 172: 117-123. |
| [5] | Kimura S, Akamatsu T, Li S H, Dong L J, Wang K X, Wang D, Arai N. Seasonal changes in the local distribution of Yangtze finless porpoises related to fish presence. Marine Mammal Science, 2012, 28(2): 308-324. |
| [6] | Wang Z T, Akamatsu T, Mei Z G, Dong L J, Imaizumi T, Wang K X, Wang D. Frequent and prolonged nocturnal occupation of port areas by Yangtze finless porpoises (Neophocaena asiaeorientalis): forced choice for feeding? Integrate Zoology, 2015, 10(1): 122-132. |
| [7] | Wang Z T, Akamatsu T, Wang K X, Wang D. The diel rhythms of biosonar behavior in the Yangtze finless porpoise (Neophocaena asiaeorientalis asiaeorientalis) in the port of the Yangtze River: the correlation between prey availability and boat traffic. PloS ONE, 2014, 9(5): e97907. |
| [8] | 张先锋, 刘仁俊, 赵庆中, 张国成, 魏卓, 王小强, 杨健. 长江中下游江豚种群现状评价. 兽类学报, 1993, 13(4): 260-270. |
| [9] | 湖北省水生生物研究所鱼类研究室. 长江鱼类. 北京: 科学出版社, 1976. |
| [10] | 朱松泉. 中国淡水鱼类检索表. 南京: 江苏科学技术出版社, 1995. |
| [11] | Tang J F, Ye S W, Li W, Liu J T, Zhang T L, Guo Z Q, Zhu F Y, Li Z J. Status and historical changes in the fish community in Erhai Lake. Chinese Journal of Oceanology and Limnology, 2013, 31(4): 712-723. |
| [12] | Ye S W, Li Z J, Lek-Ang S, Feng G P, Lek S, Cao W X. Community structure of small fishes in a shallow macrophytic lake (Niushan Lake) along the middle reach of the Yangtze River, China. Aquatic Living Resource, 2006, 19(4): 349-359. |
| [13] | 茹辉军, 王海军, 赵伟华, 沈亚强, 王勇, 张晓可. 黄河干流鱼类群落特征及其历史变化. 生物多样性, 2010, 18(2): 179-186. |
| [14] | 郁尧山, 张庆生, 陈卫民, 许源剑. 浙江北部岛礁周围海域鱼类优势种及其种间关系的初步研究. 水产学报, 1986, 10(2): 137-149. |
| [15] | 胡菊英, 姚闻卿. 长江下游安徽江段的鱼类. 安徽大学学报: 自然科学版, 1996, 20(1): 96-101. |
| [16] | 张敏莹, 徐东坡, 刘凯, 施炜纲. 长江安庆江段鱼类调查及物种多样性初步研究. 湖泊科学, 2006, 18(6): 670-676. |
| [17] | 邓朝阳, 朱仁, 严云志. 长江芜湖江段鱼类多样性及其群落结构的时空格局. 淡水渔业, 2013, 43(1): 28-36. |
| [18] | 严云志, 郭丽丽, 李国龙. 长江铜陵段老洲水域鱼类资源的初步调查研究. 安徽师范大学学报: 自然科学版, 2006, 29(6): 575-578. |
| [19] | 于道平, 黄敏毅, 赵凯, 陈寿文. 长江东流河道整治对长江江豚种群数量的影响. 兽类学报, 2012, 32(4): 330-334. |
| [20] | 李捷, 李新辉, 贾晓平, 李跃飞, 何美峰, 谭细畅, 王超, 蒋万祥. 西江鱼类群落多样性及其演变. 中国水产科学. 2010, 17(2): 298-311. |
| [21] | Fischer J, Paukert C, Daniels M. Fish community response to habitat alteration: impacts of sand dredging in the Kansans River. Transactions of the American Fisheries Society, 2012, 141(6): 1532-1544. |
| [22] | Paukert C, Schloesser J, Fischer J, Eitzmann J, Pitts K, Thornbrugh D. Effect of instream sand dredging on fish communities in the Kansas River USA: current and historical perspectives. Journal of Freshwater Ecology, 2008, 23(4): 623-633. |
| [23] | 张新桥. 洞庭湖及邻近水域长江江豚种群生态学研究 [D]. 武汉: 中国科学院水生生物研究所, 2011. |
| [24] | 于道平, 王江, 杨光, 章贤. 长江湖口至荻港段江豚春季对生境选择的初步分析. 兽类学报, 2005, 25(3): 302-306. |
| [25] | Koopman H N. Topographical distribution of the blubber of harbor porpoises (Phocoena phocoena). Journal of Mammalogy, 1998, 79(1): 260-270. |