 2016, Vol. 36
2016, Vol. 36文章信息
- 郑亚琼, 张肖, 梁继业, 李志军, 韩占江
- ZHENG Yaqiong, ZHANG Xiao, LIANG Jiye, LI Zhijun, HAN Zhanjiang
- 濒危物种胡杨和灰叶胡杨的克隆生长特征
- Clonal growth characteristics of the endangered species Populus euphratica oliv and Populus pruinosa schrenk
- 生态学报, 2016, 36(5): 1331-1341
- Acta Ecologica Sinica, 2016, 36(5): 1331-1341
- http://dx.doi.org/10.5846/stxb201407151443
-
文章历史
- 收稿日期: 2014-07-15
- 网络出版日期: 2015-07-22






2. 塔里木大学植物科学学院, 阿拉尔 843300
2. College of Plant Sciences, Tarim University, Alar 843300, China
克隆生长是克隆植物在自然条件下产生具有潜在独立性个体(克隆分株)并进行垂直水平空间拓展的营养生长过程[1]。关于木本植物克隆生长的研究,国内外主要从克隆构件[2, 3, 4]、基株[5, 6, 7, 8]和基株种群[9]3个水平开展了克隆生长格局、生理整合、生长构型、觅食行为及生长繁殖对策的相关研究。克隆繁殖受到克隆器官生长特性、空间拓展格局、克隆器官上芽库的形成以及环境因素的影响。克隆植物的不定芽可以视为构件独立生长的单位,每年新的不定芽的产生以及后来不定芽发育成子株,能够抵消母株的季节性死亡[10],进行死亡分摊,而只有有潜在独立生长能力的芽才能发育为地上子株,这表现为地上子株的密度是由地下芽库所决定。芽库在种群的更新与补充以及群落演替中起着非常重要的地位和作用。
胡杨(Populus euphratica Oliv.)和灰叶胡杨(Populus pruinosa Schrenk.)均隶属于杨柳科(Salicaceae)杨属(Populus),是干旱区荒漠河岸优势建群种的木本克隆植物,它们能够通过根克隆生长调节实施林窗更新、林缘扩散维持种群的稳定性和持久性,其克隆生长及其对环境的可塑性反应研究愈来愈受到关注。胡杨、灰叶胡杨的克隆繁殖源于根克隆器官——横走侧根的生长扩散、横走侧根内皮层部位不定芽的产生以及不定芽生长发育为克隆分株[3, 4, 11],同时还受到水盐阈值的影响[12, 13, 14, 15, 16]。只要能满足其最低生理的要求,胡杨繁殖根系就可以通过产生根蘖以其根系构型差异所决定的根系分布特性来消除不利条件环境带来的影响,并繁衍其后代、拓展本物种的分布[17]。有研究表明,胡杨根蘖苗通常来源于地表面20cm以上可以以水平“间隔”根的形式长到离母株40m的地方[18],但萌蘖根集中分布的深度范围会因地域的不同而不同,萌蘖根的分布深度也因母株年龄的不同而出现变化,胡杨根蘖繁殖幼苗的萌发点集中在40cm土层以内的横走侧根上[2, 11, 15, 16, 18, 19];灰叶胡根蘖繁殖幼苗的萌发点则集中在30cm土层以内的横走侧根上[20],分布深度超过此范围横走侧根上基本不能萌发出根蘖繁殖幼苗。研究还表明,胡杨克隆繁殖与克隆繁殖器官的粗度有一定关系,表现出细根产生根蘖苗多于粗根的特点[19, 21]。目前,对胡杨、灰叶胡杨克隆繁殖的研究主要集中在根蘖苗(克隆分株)出现、数量与萌蘖根分布深度、粗度关系、以及环境因子的关系探讨,但对克隆繁殖过程中最重要的环节——克隆器官上芽库形成特点、芽库大小与地上克隆分株(根蘖苗)数量变化的关系、以及环境因子在其过程的调节作用等方面尚缺乏了解。本研究拟探讨不同生态环境条件下,胡杨和灰叶胡杨横走侧根上不定芽及克隆分株的数量格局特征及其与土壤水分的关系,阐明克隆繁殖器官生长特征及土壤水分对不定芽(萌蘖芽)发生及不定芽输出转化的影响,为深入揭示胡杨和灰叶胡杨克隆繁殖的生态适应机制奠定基础。
1 研究区概况、实验材料和方法 1.1 研究区概况研究区选择在塔里木河上游农一师16团的天然灰叶胡杨林、叶尔羌河下游的阿瓦提县境内天然胡杨和灰叶胡杨混交林、以及塔里木河中游的轮台县境内的天然胡杨林。研究区地理位置及生境气象因子见表 1。
| 参数Parameter | 农一师16团天然灰叶胡杨纯林16 regiment natural P. euphratica forest | 阿瓦提县胡杨、灰叶胡杨混交林Awati P.euphratica and P. pruinosa mixed forests | 轮台县天然胡杨纯林Luntai natural P. pruinosa forest |
| 东经 East longitude | 80°50′ | 80°25′ | 83°38′ |
| 北纬 North latitude | 40°29′ | 39°40′ | 41°05′ |
| 海拔 Altitude/m | 1023 | 992.62 | 1002.62 |
| 年平均日照时数/hThe annual average sunshine time | 2890.6 | 2729 | 2725 |
| 年平均气温/℃ The annual average temperature | 11.28 | 10.4 | 10.6 |
| 最高温度/℃ The highest temperature | 41 | 39.4 | 40.5 |
| 最低温度/℃The lowest temperature | -26.6 | -25 | -25.5 |
| 年平均降水量/mm An average annual rainfall | 42 | 50.4 | 52 |
| 年平均蒸发量/mm The average annual vaporation | 1188 | 1880 | 2072 |
| 相对湿度/% Relative humidity | 52 | 56 | 50 |
16团天然灰叶胡杨林研究区地下水位1.08m,地势平坦,地表为细沙质,局部地段有盐结皮,每年洪水可漫及部分林地。研究区林型为灰叶胡杨纯林,伴生物种有柽柳、甘草、芦苇、小獐毛。灰叶胡杨分布在塔里木河源头河漫滩西北向,整个流域均可看到沿河岸延伸的灰叶胡杨林。由于该群落位于河漫滩受人畜影响较大,林缘萌蘖的幼苗地上部分绝大多数被牲畜啃食。乔木层盖度为36%。灌木层盖度为18%,草本层盖度为59%。群落类型为灰叶胡杨-甘草群落。
阿瓦提县胡杨和灰叶胡杨混交林研究区地下水位3.34m,地势平坦,地表松散的细沙,丰水年洪水可漫及,水分状况较差。研究区林型为胡杨和灰叶胡杨混交林,伴生物种有柽柳、黑果枸杞、羊角草、铃铛刺。胡杨和灰叶胡杨主要分布在高阶地,在冲积沟内林木密度相对较大,距河床稍远处密度相对较小。乔木层胡杨占60%,灰叶胡杨占40%,盖度为47%,灌木层盖度为39%,草本层盖度为23%。群落类型为胡杨和灰叶胡杨-柽柳群落。
轮台县天然胡杨林研究区地下水位2.56m,土壤类型由灰棕土、风沙土、盐碱土和灌淤土组成,以冲击、风积细沙为主,每年林地可漫及洪水。研究区林型为胡杨纯林,伴生物种有柽柳、花花柴、羊角草、芦苇、甘草。胡杨分布在一级阶地和河漫滩,距河床较近,沿河岸分布比较广泛。在进河床的林缘多为种子萌生和根蘖成林。乔木层盖度为42%,灌木层盖度为22%,草本层盖度为20%。群落类型为胡杨-柽柳群落。
1.2 实验材料与方法在农一师16团天然灰叶胡杨林内,沿垂直河道的方向选择河道边(A1)、距河道200m林内(A2)和距河道400m林内(A3)3个不同的生境为样地,样地大小100m×100m,该研究区地下水位1.08m;在阿瓦提县混交林内选择灰叶胡杨(B1)和胡杨(B2)相对集中的林内生境为样地,样地大小100m×100m,研究区地下水位3.34m;在轮台县天然胡杨林内选择河道边(C1)和林内(C2)2个不同的生境为样地,样地大小100m×100m,研究区地下水位2.56m。
在16团、阿瓦提县、轮台县的每个样地中随机选取分布离散的40株克隆分株,以克隆分株为目标进行跟踪挖掘。为了保证地上克隆分株和横走侧根的自然联系,先挖一个1 m×1 m的剖面,再向克隆分株两侧延伸2m跟踪挖取横走侧根的近端根和远端根(近端根指距离母株端近的横走侧根;远端根指远离母株端的横走侧根),以产生克隆分株部位(萌蘖处)的横走侧根为中心以50cm为间隔(近端根50—100cm、近端根0—50cm、中间根50cm、远端根0—50cm、远端根50—100cm)将横走侧根进行分段(图 1),统计和测量不同根段区域以下指标参数:①横走侧根距地面的深度、横走侧根的粗度;②未出土/出土克隆分株处横走侧根的粗度以及距离地面的深度;③横走侧根上的不定芽数量;④横走侧根上的未出土/出土克隆分株的数量、高度、基径。

|
| 图 1 产生克隆分株横走侧根不同根段示意图 Fig.1 The horizontal lateral root Producing clonal ramet of different root segment schematic diagram |
取克隆分株处0—60cm土层的土样,由上而下每20cm为一个土层,将土样充分混匀后迅速装入已知重量(W1)的铝盒中,盖上盒盖,带回室内,在天平上称重(W2)。打开盒盖(盖子放在铝盒旁边)放在(105±2)℃的恒温干燥箱中烘干至恒重(W3),计算土样的含水量。
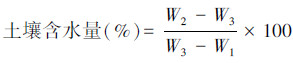
调查统计数据录入Excle表格中,计算出各指标参数的均值,以均值的大小进行作图比较,并用SPSS 13.0的单因素方差分析法对数据进行多重比较分析,试验结果以平均值±标准差表示;并用DPS软件进行相关性分析。
2 结果与分析 2.1 横走侧根空间分布特征及形态特点 2.1.1 产生克隆分株的横走侧根垂直空间分布特征表 2显示,3个研究区近端根、克隆分株所在部位(萌蘖处)的横走侧根和远端根在土壤中分布的深度均表现出远端根比近端根深,而克隆分株所在部位(萌蘖处)的横走侧根分布的深度反而比远端根和近端根浅的空间分布规律。进一步分析可以看出,16团的3个生境中灰叶胡杨克隆分株所在部位(萌蘖处)的横走侧根、近端根和远端根分布深度依次为河道边<距离河道200m林内<距离河道400m林内。3个生境中克隆分株所在部位(萌蘖处)的横走侧根均分布在10—20cm的土层内,但近端根和远端根在河道边分布在10—20cm的土层内,距离河道200m林内和距离河道400m林内则分布在20—40cm的土层内。阿瓦提混交林内,灰叶胡杨和胡杨克隆分株所在横走侧根及近端根和远端根都分布在20—40cm的土层内。轮台县胡杨天然林两个生境中,河道边胡杨近端根、克隆分株所在部位(萌蘖处)的横走侧根分布在10—20cm的表层土壤内,远端根分布在20—40cm的土层内;林内胡杨克隆分株所在部位(萌蘖处)的横走侧根、近端根和远端根均分布在20—40cm的土层内。
| 生境Habitat | 近端根深/cmProximal root depth | 萌蘖处根深/cmSprout tillers depth | 远端根深/cmDistal root depth |
| A1 | 11.60±1.28bAB | 11.45±1.46bAB | 15.85±1.61aA |
| A2 | 26.90±2.11bB | 15.05±0.45cC | 33.05±1.98aA |
| A3 | 29.85±2.33bA | 15.20±2.33cB | 37.75±2.95aA |
| B1 | 30.55±1.93abA | 28.90±2.60bA | 37.30±2.41aA |
| B2 | 32.45±1.15abA | 29.48±0.95bA | 41.70±1.62aA |
| C1 | 17.10±2.60aA | 15.95±2.63aA | 21.00±2.84aA |
| C2 | 29.15±2.12bA | 32.58±1.96abA | 40.10±2.13aA |
观察发现,有不定芽、克隆分株产生的横走侧根部位直径增粗,无不定芽、克隆分株产生的横走侧根根段区域直径较均匀,横走侧根上不定芽、未出土克隆分株和出土克隆分株密集分布部位直径的增粗比不定芽、未出土克隆分株和出土克隆分株单个分布部位直径的增粗更为明显。从表 3可以看出,不同的研究区以及同一研究区的不同生境中,产生克隆分株的横走侧根形态上均表现为出土克隆分株、未出土克隆分株和不定芽所在部位的横走侧根最粗,向两端逐渐变细,且远端根始终比近端根要粗,同一条横走侧根的不同根段根粗差异显著。
| 研究区Research area | 近端根50—100cmProximal root 50—100cm | 近端根0—50cmProximal root 0—50cm | 中间50cmMiddle root 50cm | 远端根0—50cmDistal root 0—50cm | 远端根50—100cmDistal root 50—100cm |
| A1 | 0.54±0.05cC | 0.70±0.07cBC | 1.44±0.16aA | 1.02±0.11bB | 0.71±0.07cBC |
| A2 | 0.54±0.06cC | 0.73±0.06bcBC | 1.69±0.20bcaA | 1.01±0.08bB | 0.80±0.08bcBC |
| A3 | 0.39±0.04cC | 0.65±0.07bcBC | 1.56±0.16aA | 0.89±0.09bB | 0.63±0.06bcBC |
| B1 | 0.92±0.06dD | 1.77±0.17bcBC | 3.59±0.23aA | 2.20±0.13bB | 1.43±0.12cCD |
| B2 | 1.08±0.13dD | 1.91±0.14bcBC | 3.91±0.23aA | 2.38±0.17bB | 1.52±0.16cdCD |
| C1 | 0.61±0.10bB | 0.99±0.17bAB | 1.55±0.25aA | 1.14±0.19bB | 0.76±0.12bB |
| C2 | 1.46±0.12cB | 2.33±0.10dC | 4.55±0.16aA | 2.76±0.14bB | 1.70±0.15dC |
| 检验方法为LSD,同一行不同小写字母表示在0.05水平上显著差异,同一行不同大写字母表示在0.01水平上显著差异 | |||||
不定芽、未出土克隆分株和出土克隆分株的关系决定了它们在横走侧根上的分布格局是一致的。观察发现,不定芽呈现单个或多个聚集分布,聚集在一起的不定芽、未出土克隆分株和出土克隆分株或环绕分布在横走侧根的不同位点上,或分布在横走侧根的同一个侧面(图 2—图 4)。

|
| 图 2 胡杨和灰叶胡杨横走侧根及其上不定芽空间分布的特征 Fig.2 The horizontal lateral root of P. euphratica and P. pruinosa root and adventitious bud space distribution characteristics |

|
| 图 3 胡杨和灰叶胡杨横走侧根及其上未出土克隆分株空间分布的特征 Fig.3 The horizontal lateral root ofP. pruinosa and P. euphratica and not unearthed clonal ramet space distribution characteristics |

|
| 图 4 胡杨和灰叶胡杨横走侧根及其上出土克隆分株空间分布的特征 Fig.4 The horizontal lateral root ofP. euphratica and P. pruinosa unearthed clonal ramet space distribution characteristics |
以克隆分株所在横走侧根的区段为中间50cm段向两端统计单位长度(每50cm为一个单位长度)的根段上不定芽、未出土克隆分株和出土克隆分株出现的频率。结果显示(表 4—表 6),不定芽、未出土克隆分株和出土克隆分株均表现出在横走侧根中间50cm根段出现频率最高,在0—50cm根段(不论近端根还是远端根)出现频率明显减小,在50—100cm根段出现频率更低或为零,表现出以已有克隆分株为中心向两端延伸发生分布的特征,反映了不定芽发生、不定芽转化为克隆分株数量和空间分布范围与已有克隆分株的一种联系。同一生境下同一根段相比较,不定芽、未出土克隆分株和出土克隆分株出现的频率均表现为不定芽>未出土克隆分株>出土克隆分株。不同研究区相比较,可以明显看出地下水位3.34m的阿瓦提县胡杨林,在已经有克隆分株发生的部位及其附近均没有不定芽产生,地下水位2.56m的轮台县胡杨林其不定芽、未出土克隆分株出现频率和空间分布范围居中,而地下水位1.08m的16团胡杨林其不定芽、未出土克隆分株出现频率和空间分布范最大,说明地下水位对不定芽发生及向克隆分株转化有一定的影响。
| 研究区Research area | 近端根50—100cm/%Proximal root 50—100cm | 近端根0—50cm/%Proximal root 0—50cm | 中间50cm/%Middle root 50cm | 远端根0—50cm/%Distal root 0—50cm | 远端根50—100cm/%Distal root 50—100cm |
| A1 | — | 25 | 100 | 45 | 5 |
| A2 | — | 30 | 100 | 20 | 15 |
| A3 | 10 | 20 | 100 | 25 | — |
| B1 | — | — | — | — | — |
| B2 | — | — | — | — | — |
| C1 | 10 | 35 | 65 | 20 | — |
| C2 | — | — | — | — | — |
| 研究区Research area | 近端根50—100cm/%Proximal root 50—100cm | 近端根0—50cm/%Proximal root 0—50cm | 中间50cm/%Middle root 50cm | 远端根0—50cm/%Distal root 0—50cm | 远端根50—100cm/%Distal root 50—100cm |
| A1 | — | 10 | 65 | 30 | — |
| A2 | — | 20 | 45 | 10 | — |
| A3 | 15 | — | 55 | 10 | — |
| B1 | — | — | — | — | — |
| B2 | — | — | — | — | — |
| C1 | — | 60 | 50 | — | — |
| C2 | — | — | 10 | — | — |
| 研究区Research area | 近端根50—100cm/%Proximal root 50—100cm | 近端根0—50cm/%Proximal root 0—50cm | 中间50cm/%Middle root 50cm | 远端根0—50cm/%Distal root 0—50cm | 远端根50—100cm/%Distal root 50—100cm |
| A1 | — | — | 100 | 15 | — |
| A2 | — | 5 | 100 | 5 | — |
| A3 | — | 5 | 100 | 5 | — |
| B1 | — | — | 100 | — | — |
| B2 | — | — | 100 | — | — |
| C1 | — | 60 | 100 | — | — |
| C2 | — | — | 100 | — | — |
从图 5可以看出,3个研究区中只有16团3个生境(A1、A2、A3)的灰叶胡杨和轮台河道边生境(C1)的胡杨横走侧根上能产生不定芽,共同表现出中间50cm根段不定芽、未出土和出土克隆分株数量较多,向两端急剧减少到几乎没有。16团3个生境相比较,不定芽、未出土和出土克隆分株数量均表现出河道边(A1)>距离河道200m林内(A2)>距离河道400m林内(A3),即灰叶胡杨横走侧根上不定芽、未出土克隆分株和出土克隆分株的数量随着距离河道越远数量越少。轮台县胡杨林两个生境相比较,仅河道边的胡杨其横走侧根有不定芽的产生,未出土和出土克隆分株的数量河道边远远高于林内。在阿瓦提县林内所调查的胡杨和灰叶胡杨横走侧根有出土克隆分株,但在其周围没有不定芽的产生和未出土克隆分株,表明地下水位对不定芽的发生有影响。

|
| 图 5 横走侧根不同根段不定芽、未出土克隆分株和出土克隆分株的数量特征 Fig.5 The adventitious bud quantitative characteristic on the horizontal lateral root |
不定芽发生、不定芽生长为未出土克隆分株、到形成出土克隆分株是克隆生长的3个连续阶段。表 7显示,横走侧根上不定芽数与出土克隆分株数呈极显著正相关,未出土克隆分株数与出土克隆分株数呈显著正相关,说明不定芽、未出土克隆分株的数量都直接影响出土克隆分株的数量。
| 相关系数Correlation coefficient | 不定芽数Number of adventitious bud | 出土克隆分株数Number of not unearthed clone ramets | 未出土克隆分株数Number of unearthed clone ramets |
| 不定芽数Number of adventitious bud | 1.00 | ||
| 出土克隆分株数Number of not unearthed clone ramets | 0.99 ** | 1.00 | |
| 未出土克隆分株数Number of unearthed clone ramets | 0.79 | 0.85 * | 1.00 |
| * P < 0.05,** P < 0.01 | |||
从图 6中0—60cm土壤含水量的平均值对克隆生长的影响可以看出,不同研究区以及同一研究区不同生境的土壤含水量不同,胡杨和灰叶胡杨克隆生长特征也不同。16团3个生境随着距离河道越远土壤含水量越低,横走侧根上的不定芽、未出土克隆分株和出土克隆分株的数量越少;阿瓦提林内土壤含水量较低(10%左右)胡杨、灰叶胡杨横走侧根上均没有新生不定芽和未出土克隆分株的产生,横走侧根上产生的出土克隆分株的数量相等;轮台河道边土壤含水量高于林内土壤含水量,河道边胡杨横走侧根上不定芽、未出土克隆分株和出土克隆分株数量高于林内胡杨。由表 8可以看出,不定芽和出土克隆分株数量与土壤水分含量呈极显著正相关,即随着土壤含水量的升高,横走侧根上不定芽数量和出土克隆分株数量越多。分析表明,不定芽能否转化为出土克隆分株及其数量的多少与横走侧根所在环境土壤水资源有密切关系。

|
| 图 6 土壤含水量对不定芽、出土克隆分株和未出土克隆分株的影响 Fig.6 The influence of soil moisture content for adventitious bud, not unearthed clone ramets and unearthed clone ramets |
| 相关系数Correlation coefficient | 不定芽数Number of adventitious bud | 出土克隆分株数Number of not unearthed clone ramets | 未出土克隆分株数Number of unearthed clone ramets |
| 土壤含水量Soil moisture content/% | 0.99 ** | 0.99 ** | 0.70 |
| *P < 0.05,**P < 0.01 | |||
胡杨根系在0—4.6m的土层皆有分布,而萌蘖根分布深度一般在1m以内,多集中分布在5—60cm的土层范围内,土层40cm以内是胡杨根系萌生不定芽的有效深度,分布深度大于40cm的根系上不能萌生不定芽[2, 11, 15, 18, 19];灰叶胡杨一级横走侧根集中分布在20—40cm范围内,不同生境条件下能够产生克隆分株的横走侧根垂直空间分布几近一致,主要分布在距地面5—30cm土层内[20]。本研究结果显示,3个生境中克隆分株所在部位(萌蘖处)的横走侧根均分布在10—20cm的土层内,但近端根和远端根在河道边分布在10—20cm的土层内,距离河道200m林内和距离河道400m林内则分布在20—40cm的土层内;不论是在地下水位1.08m的16团灰叶胡杨林还是地下水位3.34m的阿瓦提胡杨和灰叶胡杨混交林,产生不定芽和克隆分株的横走侧根主要分布在10—20cm的土层中,这与前人的研究结果是一致。有研究指出,随着土壤深度的增加,土壤通气状况的变差,减弱了根上萌生不定芽的能力,到达一定深度的土壤以后,根上就不再萌生不定芽;根上萌生出的不定芽在突破土层时会遇到土壤的阻力,相对来说较深层次的土壤更加紧实,会增加不定芽突破土层向上生长的阻力,不利于根蘖繁殖幼苗的出土[22]。胡杨和灰叶胡杨横走侧根集中分布在10—30cm土层,会使其接受更多的光热资源,有利于不定芽的产生以及不定芽转化为克隆分株,这可能也是胡杨和灰叶胡杨根系适应克隆生长的空间分布策略之一。
3.2 胡杨和灰叶胡杨克隆分株横走侧根形态特征与不定芽发生的关系尽管胡杨具有发达的侧根根系,但多数侧根较细,粗根数量占有的比例较少,并且具有繁殖能力的萌蘖根根系也表现出细根多于粗根的特点[21]。当生境条件相对适宜时,直径在0.1—4.0cm范围的水平根均可产生出根蘖苗,且以直径在0.1—1.6cm范围根系的出苗率最高,达到83.95%,而﹥4.0cm的根系几乎无根蘖苗出现,萌蘖能力基本丧失,这可能与直径较大的根系具有较高的木质化程度和较低的分生组织活力有关[19]。我们的观察结果显示,不定芽在一定粗度范围内的横走侧根上才会产生,并且横走侧根越细其上不定芽数量越多,超过一定粗度胡杨和灰叶胡杨横走侧根上不会产生不定芽。研究还发现,胡杨和灰叶胡杨横走侧根上有不定芽、未出土克隆分株/出土克隆分株产生的部位直径增粗,无不定芽、未出土克隆分株/出土克隆分株产生的部位直径较均匀。分析认为,横走侧根的这种形态变化可能是已经形成的克隆分株产生光合产物并在其着生部位附近积累的结果,这一结果同时为其周围产生更多不定芽提供了物质基础,这种形态上的变化其实质上也反映了已经形成的克隆分株和新一轮不定芽发生之间的内在关系。Martin Wiehle等的研究得到相似结果,认为这是由于根蘖苗的同化吸收,远端根很快就变得比它的近端根粗,它决定了营养生长的方向[18]。
3.3 胡杨、灰叶胡杨不定芽的空间分布与繁殖成功率的关系有研究报道,萌蘖根上的根蘖节部位通常产生的不定芽不止一个,通常以环状不均匀地分布,并且部分芽点相互重叠的现象也十分普遍,只要环境条件适宜,这些芽点均发育为不定芽,这是胡杨根蘖苗多呈丛状分布的内在原因,也反映出胡杨具有较强根蘖繁殖能力的属性[3, 4, 11]。不定芽的产生大多位于根蘖苗的末级端,造成此结果的原因可能是与根的分工有关,末端部位主要负责寻找有利于生长的生境,而只要条件适宜,在横走侧根的任何一个部位都能产生不定芽[9]。本研究发现,不定芽在克隆分株所在部位的50cm中间区段最高,在0—50cm根段(不论近端根还是远端根)不定芽出现的频率明显减小,在50—100cm根段出现的频率更低或为零,不定芽发生及空间分布表现出以克隆分株为中心向两端延伸发生分布的特征。分析认为,不定芽围绕克隆分株集中发生的特征是前期形成的克隆分株(根蘖苗)将光合产物优先运输和积累在其着生部位附近的结果,这也是克隆分株所在部位的区间不定芽发生及转化为克隆分株成功率最高的重要原因之一。前期克隆分株的存在和生长发育对于后期新一轮不定芽的发生、未出土克隆分株转化为出土克隆分株具有重要的生理整合意义。有研究表明,地上分蘖的变化最终是由芽库决定的,它们之间存在一个正相关关系[23]。我们的研究得到类似的结果,横走侧根上不定芽数量与出土克隆分株的数量呈极显著正相关,未出土克隆分株数与出土克隆分株数间呈显著正相关,表明不定芽、未出土克隆分株的数量都直接影响出土克隆分株的数量。同一生境不定芽、未出土克隆分株和出土克隆分株出现的频率相表现为不定芽>未出土克隆分株>出土克隆分株,也进一步说明芽库不定芽的数量决定着地上克隆分株数量的变化,从不定芽发生到出土克隆分株形成存在较大的风险。
3.4 胡杨、灰叶胡杨不定芽发生和克隆分株生长与土壤水分条件的关系大量的研究表明,在土壤水分养分条件较好、光照强度较大的条件下,植物通过无性繁殖可以产生更多的子株后代,通过子株的聚集以增加对土壤水分养分及光照的获取,充分利用资源[24, 25]。胡杨作为依靠地下潜水生长发育的隐域性植被类型,水分条件的优劣同样会对胡杨的繁殖与更新过程造成重大影响[14, 26]。胡杨根蘖生长发育的关键性因素是受到地下水埋深和土壤条件等的影响[11, 15],且地下水埋的深度决定着胡杨根蘖繁殖能否成功,如地下水位在 1.33—2.17m范围内,林下营养繁殖的无性系小株较多;地下水位在2—4m范围内,无性系小株明显减少;在7—10m范围或者更深,无性系小株几乎为零,这是由于地下水位过低导致胡杨萌蘖根根系因土壤干燥而无法长出根蘖芽[12]。除了地下水位,土壤含水量、土壤总盐含量和草灌盖度是影响胡杨根蘖苗发生数量、生长状况以及胡杨的地下萌蘖根根系特征的主要环境因素。其中,土壤含水量与胡杨根蘖苗的发生数量、幼苗株高,以及每一胡杨根蘖节的产苗数量呈显著的正相关关系(P < 0.05)[13]。本文的调查结果显示,研究区16团天然灰叶胡杨林地下水位1.08m,每年洪水可漫及部分林地,其不定芽、未出土克隆分株出现频率和空间分布范均最大;轮台县天然胡杨林地下水位2.56m,每年林地可漫及洪水,其不定芽、未出土克隆分株出现频率和空间分布范围居中;阿瓦提县胡杨和灰叶胡杨混交林研究区地下水位3.34m,丰水年洪水可漫及,水分状况较差,在已经有克隆分株发生的部位及其附近均没有不定芽产生,表明地下水位对不定芽发生及向克隆分株转化有一定的影响。相关分析结果也进一步表明,土壤水分含量与不定芽和出土克隆分株数量均呈极显著正相关。综合分析认为,地下水位以及每年洪水是否漫及林地和漫及林地的水量均对不定芽发生及不定芽向克隆分株转化有显著的影响。
| [1] | de Kroon H, van Groenendael J. The Ecology and Evolution of Clonal Plants. Leiden, The Netherlands: Backhuys Publishers, 1997. |
| [2] | 王永斌, 努尔巴依·阿布都沙力克, Thevs N. 胡杨(Populus euphratica Oliv.)根繁殖特征. 生态学杂志, 2007, 26(12): 1937-1941. |
| [3] | 李志军, 焦培培, 周正立, 李倩, 李健强. 胡杨横走侧根及不定芽发生的形态解剖学研究. 北京林业大学学报, 2011, 33(5): 42-48. |
| [4] | 李志军, 焦培培, 周正立, 李倩, 李健强. 灰叶胡杨根蘖繁殖的形态解剖学特征. 植物学报, 2012, 47(2): 133-140. |
| [5] | 廖明隽, 王其兵, 宋明华, 董鸣. 内蒙古锡林河流域不同生境中羊草的克隆构型和分株种群特征. 植物生态学报, 2002, 26(1): 33-38. |
| [6] | 罗学刚, 董鸣. 匍匐茎草本蛇莓克隆构型对土壤养分的可塑性反应. 生态学报, 2001, 21(12): 1954-1963. |
| [7] | 袁秀英, 袁登胜, 白玉明. 固定和半固定沙丘花棒克隆生长构型的比较研究. 西北林学院学报, 2006, 21(6): 32-34. |
| [8] | 陈玉福, 董鸣. 毛乌素沙地根茎灌木羊柴的基株特征和不同生境中的分株种群特征. 植物生态学报, 2000, 24(1): 40-45. |
| [9] | 刘庆, 钟章成. 斑苦竹无性系种群克隆生长格局动态的研究. 应用生态学报, 1996, 7(3): 240-244. |
| [10] | Hendrickson J R, Briske D D. Axillary bud banks of two semiarid perennial grasses: occurrence, longevity, and contribution to population persistence. Oecologia, 1997, 110(4): 584-591. |
| [11] | 李志军, 刘建平, 于军, 周正立. 胡杨、灰叶胡杨生物生态学特性调查. 西北植物学报, 2003, 23(7): 1292-1296. |
| [12] | 乌日根夫, 战士宏, 程继全, 张晓丽, 班振国. 额济纳旗天然胡杨林生物学、生态学抗旱机理与繁殖机理研究. 内蒙古林业调查设计, 2003, 26(4): 1-5. |
| [13] | 崔德宝. 艾比湖自然保护区阿其克苏河流域胡杨根蘖繁殖特征及影响因素研究[D]. 新疆: 新疆师范大学, 2011. |
| [14] | 赵文智, 常学礼, 李秋艳. 人工调水对额济纳胡杨荒漠河岸林繁殖的影响. 生态学报, 2005, 25(8): 1987-1993. |
| [15] | 武逢平, 李俊清, 李景文, 程春龙, 王旭航. 胡杨(Populus euphratica)在额济纳绿洲三种生境内的根蘖繁殖特性. 生态学报, 2008, 28(10): 4703-4709. |
| [16] | 曹德昌, 李景文, 陈维强, 彭程, 李俊清. 额济纳绿洲不同林隙胡杨根蘖的发生特征. 生态学报, 2009, 29(4): 19554-1961. |
| [17] | 黄晶晶, 井家林, 曹德昌, 张楠, 李景文, 夏延国, 吕爽. 不同林龄胡杨克隆繁殖根系分布特征及其构型. 生态学报, 2013, 33(14): 4331-4342. |
| [18] | Wiehle M, Eusemann P, Thevs N, Schnittler M.Root suckering patterns in Populus euphratica (Euphrates poplar, Salicaceae). Trees, 2009, 23(5): 991-1001. |
| [19] | 张昊, 李俊清, 李景文, 张玉波, 孙立, 吴平, 赵健. 额济纳绿洲胡杨种群繁殖物候节律特征的研究. 内蒙古农业大学学报: 自然科学版, 2007, 28(2): 60-66. |
| [20] | 郑亚琼, 周正立, 李志军. 灰叶胡杨横走侧根空间分布与克隆繁殖的关系. 生态学杂志, 2013, 32(10): 2641-2646. |
| [21] | 冯起, 司建华, 李建林, 席海洋. 胡杨根系分布特征与根系吸水模型建立. 地球科学进展, 2008, 23(7): 765-772. |
| [22] | 朱雅娟, 董鸣, 黄振英. 种子萌发和幼苗生长对沙丘环境的适应机制. 应用生态学报, 2006, 1(1): 137-142. |
| [23] | Hartnett D C, Setshogo M P, Dalgleish H J. Bud banks of perennial savanna grasses in Botswana. African Journal of Ecology, 2006, 44(2): 256-263. |
| [24] | 罗学刚, 董鸣. 蛇莓克隆构型对光照强度的可塑性反应. 植物生态学报, 2001, 25(4): 494-497. |
| [25] | 盛海彦, 李军乔, 杨银柱, 李红英. 土壤水分对鹅绒委陵菜表型可塑性的影响. 干旱地区农业研究, 2004, 22(3): 119-122. |
| [26] | Sack L, Grubb P J, Marañón T. The functional morphology of juvenile plants tolerant of strong summer drought in shaded forest understories in southern Spain. Plant Ecology, 2003, 168(1): 139-163. |