 2016, Vol. 36
2016, Vol. 36文章信息
- 余欣超, 陈珂璐, 姚步青, 马真, 王文颖, 王慧春, 赵新全, 周华坤
- YU Xinchao, CHEN Kelu, YAO Buqing, MA Zhen, WANG Wenying, WANG Huichun, ZHAO Xinquan, ZHOU Huakun.
- 模拟增温下门源草原毛虫幼虫生长发育特征
- Effects of simulated warming on the growth and development of Gynaephora menyuanensis larvae
- 生态学报[J]. 2016, 36(24): 8002-8007
- Acta Ecologica Sinica[J]. 2016, 36(24): 8002-8007
- http://dx.doi.org/10.5846/stxb201505251046
-
文章历史
- 收稿日期: 2015-05-25
- 网络出版日期: 2016-03-31
- 修订日期: 2016-03-31





2. 中国科学院成都生物研究所, 成都 610041;
3. 中国科学院大学, 北京 100049;
4. 青海师范大学生命与地理科学学院, 西宁 810008;
5. 青海省寒区恢复生态学重点实验室, 西宁 810008
2. Chengdu Institute of Biology, Chinese Academy of Science, Chengdu 610041, China;
3. University of the Chinese Academy of Science, Beijing 100049, China;
4. Academy of Life and Geography Science, Qinghai Normal University, Xining 810008, China;
5. Key laboratory of restoration ecology of cold area in Qinghai Province, Xining 810008, China
各种观测数据表明,全球变暖已是一个不争的事实[1-2]。自工业化以来,气温已经上升了约1℃[3],尤其是最近30年来,气温每10a约上升0.2℃[4]。青藏高原由于海拔高,其温度变化更明显[5-6]。其中,数量最多、分布最广的昆虫首当其冲[7]。昆虫是变温动物,对温度的变化最为敏感[8-9]。温度是直接影响植食性昆虫的主要环境因素,直接影响其生长发育、生存繁殖、种群数量及分布范围[10],并能加快其生长发育,导致其发生期提前,适生区域扩大等[11-12]。
青海门源草原毛虫Gynaephora menyuanensis Yan et Chou属鳞翅目Lepidoptera毒蛾科Lymantriidac草原毛虫属Gynaephora,是1997年发现的新种,广泛分布于青海北部高寒草甸地区。门源草原毛虫主要在幼虫期取食禾本科和莎草科牧草[13],如爆发或大发生则会对高寒草甸造成重大危害,引起家畜食物短缺,并改变草地植物群落结构,加剧草地退化和草地生态环境恶劣,而且还会导致家畜中毒,严重阻碍了草地畜牧业的健康发展[14],是青藏高原高寒草甸地区的主要害虫之一[15]。因此,研究气温升高对其幼虫的生长发育和生存的影响有极重要的现实意义。严林等[16]在实验室研究不同温度(恒温及变温)下门源草原毛虫幼虫的反应,取得了很多重大成果。本研究在野外利用OTC进行动态的模拟增温,旨在测定不同增温梯度下青海草原毛虫幼虫期的发育特性(发育历期、相对生长率、体重和存活率),为有效预测未来温度升高时草原毛虫种群动态提供基础资料。
1 研究区域概况中国科学院海北高寒草甸生态系统定位站位于青藏高原东北隅,祁连山北支冷龙岭东段南麓的平缓滩地,地理位置为37°29′—37°45′N,101°12′—101°23′E,海拔为3200—3600 m。该地区气候为典型的高原大陆性气候,无明显四季之分,仅有冷暖季之别,冷季漫长、干燥而寒冷,暖季短暂、湿润而凉爽。温度年差较小而日差较悬殊,太阳辐射强烈。土壤为高山草甸土和高山灌丛草甸土,土壤表层和亚表层中的有机质含量丰富。植被建群种为矮嵩草(Kobresia humilis),主要优势种为异针茅(Stipa aliena)、草地早熟禾(Poa pratensis)、麻花艽(Gentiana straminea)和发草(Deschampsia caespitosa)等[17]。
2 研究方法 2.1 样地与样方布置于2014年4月上旬开始观察实验。实验平台为矮嵩草草甸模拟增温试验样地,采用国际冻原计划(International Tundra Experiment,ITEX)规定的圆台形开顶式生长小室(Open Top Chambers,OTC)模拟增温。开顶式增温室(OTC)于2012年建立。OTC设4个规格(A、B、C、D型)模拟出不同的增温梯度,OTC底径依次为2.05,1.75,1.45,1.15m,顶径依次为1.60,1.30,1.00,0.70 m,圆台高度0.4 m,底角60°。随机选取OTC附近的露天草甸作对照(CK),每个处理及对照设置5个重复。
在每个OTC和对照中放置一个细网铁丝扣笼,长35 cm,宽25 cm,高40 cm。每个扣笼内放置10只刚结束越冬且体重、大小等基本相同的门源草原毛虫低龄幼虫。本研究总共使用门源草原毛虫250只。
2.2 取样与处理方法从4月牧草返青到9月底,样地的温度采用HOBO温度仪自动记录(1次/h)。每隔7 d测量毛虫的数量、体重。由于实验的连贯性不能处死毛虫,所测的毛虫质量为鲜重,用0.0001 g天平测量,为排除误差,每次测量时间一致,均为15:00—16:00。幼虫的虫龄结合实际观测(脱皮)和头壳法确定[18],毛虫的相对生长速率(RGR)通过以下公式计算:
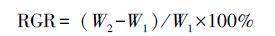
|
式中,W1为初测体重;W2为终测体重[19]。各龄期毛虫的体重依据严林[16]的研究可由体重、头壳宽等指标转化而得。
2.3 数据处理使用Microsoft Excel对各项测量数据进行描述性统计分析,采用统计软件SPSS单因素方差分析的方法对OTC与对照的各项数据进行差异性检验。文中实验数值以平均值±标准误差表示,以P<0.05作为差异显著的标准。
3 结果与分析 3.1 增温幅度基于毛虫的生活环境,本次实验测量的温度为离地5—10 cm处的气温。如图 1所示,在毛虫生长季节(4—9月),实验地的4个梯度A、B、C、D内OTC平均增温幅度分别为0.40、1.06、1.26℃和1.98℃。增温效果十分显著(P<0.05),增温幅度遵循从A、B、C、D依次增大的规律。

|
| 图 1 OTC增温效果 Fig. 1 OTC warming effect OTC为开顶式增温设备;CK为对照,A、B、C、D为增温处理;不同小写字母表示差异显著 |
门源草原毛虫的生活史分为卵期、幼虫期、蛹期和成虫期4个发育阶段,幼虫孵化出来后即进入滞育期,以1龄幼虫越冬,期间不取食,次年4月开始活动[18]。表 1中开始日期为毛虫一龄幼虫越冬后,解除滞育开始活动的日期;结束日期为毛虫幼虫全部进入蛹期的日期;发育历期为毛虫越冬后开始活动到化蛹的这段时间。如表 1所示,在CK、A、B、C和D中,毛虫一龄幼虫开始活动的日期分别为4月的24日,22日,21日,20日和18日;而全部结束幼虫期,进入蛹期的日期分别为8月16日,06日,02日,7月26日和8月1日;在CK、A、B、C和D温室中,发育历期分别为(114±3.7) d,(105±4.2) d,(98±5.6) d,(93±3.2) d,(102±4.0) d。其中A与B、B与C、B与D各温室间差异不显著(P>0.05),其他之间差异均显著(P<0.05)。
| 增温梯度 Warming gradient | 开始日期 Start date | 结束日期 The end date | 历期 Duration |
| CK | 4月24日 | 8月16日 | 114±3.7a |
| A | 4月22日 | 8月06日 | 105±4.2bc |
| B | 4月21日 | 8月02日 | 98±5.6cd |
| C | 4月20日 | 7月 26日 | 93±3.2d |
| D | 4月18日 | 8月01 日 | 102±4.0c |
| CK为对照,A、B、C、D为增温处理;不同小写字母表示差异显著 | |||
按月份比较,在5月,毛虫相对生长率从小到大依次为CK、A、B、D和C,除B和D之间外,其他彼此间均差异显著(P<0.05)(图 2);在6月,CK的相对生长速度最大,其次为C、B、A、和D,其中CK和增温处理差异均显著(P<0.05),A、C和D间彼此差异显著(P<0.05),B与A、C、D间差异均不显著(P>0.05)(图 2);在7月,各处理的生长速率差异和6月保持一致(图 2)。

|
| 图 2 模拟增温对毛虫生长速度的影响 Fig. 2 Effect of simulated warming on the Relative consumed rate of Gynaephora menyuanensis |
按处理比较,从5月至6月,CK的生长速率逐渐变大,而增温处理的A、B、C和D均逐渐变小。5月CK的相对生长速率最小,而6和7月大于增温处理(P<0.05)(图 2)。
3.4 增温对毛虫幼虫体重的影响如图 3所示,2龄期的毛虫幼虫体重基本一致,差异不显著(P>0.05);从3龄期开始,不同处理的幼虫体重出现了差异,其中CK显著大于C和D(P<0.05),A与B显著大于D(P<0.05),而CK、A、B间差异均不显著(P>0.05),A、B、C间差异不显著(P>0.05),C与D差异不显著(P>0.05);在4龄期,各处理体重与3龄期一致;在5龄期,各处理下体重出现了CK>A>B>C>D的趋势,其中CK与A、A与B、C与D之间差异不显著外(P>0.05),其他之间均差异显著(P<0.05),尤其是CK与D差异极显著(P<0.01);毛虫6龄期,雌虫与雄虫各处理下体重变化一致,为CK>A>B>C>D,其中A与B,B与C,C与D间差异不显著(P>0.05),其他之间均差异显著(P<0.05),其中CK与C、D间差异为极显著(P<0.01);7龄期,各处理体重与6龄一致。

|
| 图 3 模拟增温对体重的影响 Fig. 3 Simulated warming impact on weight |
本次实验统计了毛虫幼虫越冬后至化蛹前这一发育阶段的存活率。结果表明,在不同的增温条件下,毛虫的存活率不同。如图 4所示,在对照及增温处理A、B和C中,存活率随增温幅度的增加而升高,除了A和B处理之间外,其他处理之间均差异显著(P<0.05),其中CK与C为差异极显著(P<0.01)。增温处理D中存活率率最小,与其他处理差异显著(P<0.05),其中C与D间为差异极显著(P<0.01)。

|
| 图 4 模拟增温对存活率的影响 Fig. 4 Simulated warming effects on survival rate of Gynaephora menyuanensis larva |
门源草原毛虫以滞育的1龄幼虫越冬,翌年的4月上旬左右开始活动,其开始活动的时间与温度有直接关系[16],而昆虫的发育起始时间和发育历期的长短是昆虫生活史的重要组成部分,对世代数有直接影响[20]。
在本次实验中,与对照相比,模拟增温使A、B、C和D中1龄幼虫越冬后开始活动的时间分别提前2、3、4、6 d,即增温幅度越大的处理中的1龄幼虫越冬后开始开始活动的日期越提前;使A、B、C和D中毛虫蛹期分别提前10,14,22 d和15 d,即A—C梯度下增温越高,毛虫的蛹期提前越长,而D处理下提前的天数较C的短,说明增温过高限制了蛹期的提前;在CK、A、B、C中,发育历期随着增温梯度的增加有所变短。因此,在一定范围内,增温后1龄幼虫越冬后开始活动的日期和进入蛹期的日期提前,且毛虫幼虫期的发育历期变短。
4.2 增温对毛虫幼虫相对生长速率的影响昆虫的生长速度是生活史的重要特征[21],生长速度快的昆虫在竞争中有更大的优势,可能提前繁殖期来扩大种群数量[22]。当前全球平均温度正在上升[23],昆虫是变温动物,温度对其生长速度有直接影响[24],在发育温度区内,温度上升使昆虫的生长速度加快[25-26]。本次实验的结果也证明了这一结论,从5月到7月,增温处理中,毛虫的生长速率均为A、B、C依次变大,而D下降,这说明不是增温越高其生长速率越大,而是在一定温度范围内,温度升高会提高生长速率。
在试验中还发现:在5月,CK的生长速率小于A、B、C、D。而在6月和7月,CK的生长速率大于A、B、C、D。分析可知,CK在5、6、7月的生长速度略有增加,这是比较正常的,基本符合Books-Dyar法则,即鳞翅目昆虫幼虫生长率较恒定[27]。而增温处理下(A、B、C、D),毛虫的相对生长速度在5月达到最高,此后一直减小。这说明模拟增温对毛虫的生长影响显著,使毛虫幼虫的发育提前了,使其在5月达到了生长高峰。造成这种现象的可能原因是5月份相对6月和7月来说温度较低,而且此时毛虫处于低龄幼虫期,发育所需有效积温较老龄幼虫期少,因此增温处理下效果更明显,相对生长速度显著增大。
4.3 增温对毛虫个体体重的影响结果表明,在不同处理下,2龄期的毛虫幼虫体重基本一致。随着时间变长,毛虫发育到3龄,CK与增温处理的毛虫体重出现了差异,表现为增温幅度最大的D中毛虫体重显著小于CK(P<0.05)。当毛虫发育到5龄,开始出现CK>A>B>C>D的趋势,但趋势还不很明显,因为其中CK与A、A与B、B与C、C与D之间差异不显著(P>0.05)。毛虫发育到6龄时,CK>A>B>C>D的趋势更加明显,其中只有A与B、B与C、C与D间差异不显著(P>0.05),其他之间均差异显著(P<0.05),而7龄毛虫与6龄体重趋势相似。分析可知,和对照相比,从3龄开始,增温后毛虫幼虫体重有减轻趋势。
4.4 增温对毛虫幼虫期存活率的影响由于幼虫期较长,各处理下毛虫的个体死亡主要集中在幼虫期,因此幼虫期的生存状况是决定整个世代存活率的关键。在本次实验里,对照及增温处理A、B和C中,存活率随增温的增加而上升,到D时存活率急速下降,小于CK。因此,在适当的增温梯度下(A、B、C),温度升高可提高毛虫幼虫的存活率,但是增温过高(D)不利于其存活。
5 结论研究发现OTC模拟增温对门源草原毛虫幼虫生长发育、存活及个体大小有显著影响:随着增温变大,1龄幼虫越冬后开始活动期和蛹期提前,毛虫幼虫期的发育历期缩短;增温导致了毛虫幼虫发育模式的改变,使得其发育高峰期提前到5月份;增温后毛虫幼虫的体重减小;在适当的增温条件下(0—1.26℃),温度升高导致存活率增大,但是过高的增温(1.98℃)会导致存活率急剧下降。该研究可以丰富增温背景下草原毛虫生长发育特征研究的资料,为气候变化条件下对草原毛虫危害的应对和管理提供科学依据。
| [1] | Marcott S A, Shakun J D, Clark P U, Mix A C. A reconstruction of regional and global temperature for the past 11,300 years. Science , 2013, 339 (6124) : 1198–1201. DOI:10.1126/science.1228026 |
| [2] | Hansen J, Sato M, Ruedy R. Perception of climate change. Proceedings of the National Academy of Sciences of the United States of America , 2012, 109 (37) : E2415–E2423. DOI:10.1073/pnas.1205276109 |
| [3] | 王绍武, 罗勇, 赵宗慈, 闻新宇, 黄建斌. 工业化以来全球平均温度上升了1℃. 气候变化研究进展 , 2013, 9 (5) : 383–385. |
| [4] | Hansen J, Sato M, Ruedy R, Lo K, Lea D W, Medina-Elizade M. Global temperature change. Proceedings of the National Academy of Sciences of the United States of America , 2006, 103 (39) : 14288–14293. DOI:10.1073/pnas.0606291103 |
| [5] | 马晓波, 李栋梁. 青藏高原近代气温变化趋势及突变分析. 高原气象 , 2003, 22 (5) : 507–512. |
| [6] | 魏凤英, 曹鸿兴, 王丽萍. 20世纪80-90年代我国气候增暖进程的统计事实. 应用气象学报 , 2003, 14 (1) : 79–86. |
| [7] | Robinet C, Roques A. Direct impacts of recent climate warming on insect populations. Integrative Zoology , 2010, 5 (2) : 132–142. DOI:10.1111/inz.2010.5.issue-2 |
| [8] | 时培建, 池本孝哉, 戈峰. 温度与昆虫生长发育关系模型的发展与应用. 应用昆虫学报 , 2011, 48 (5) : 1149–1160. |
| [9] | 马春森, 马罡, 杜尧, 杨和平. 连续温度梯度下昆虫趋温性的研究现状与展望. 生态学报 , 2005, 25 (12) : 3390–3397. |
| [10] | Bale J S, Masters G J, Hodkinson I D, Awmack C, Bezemer T M, Brown V K, Butterfield J, Buse A, Coulson J C, Farrar J, Good J E G, Harrington R, Hartley S, Jones T H, Lindroth R L, Press M C, Symrnioudis I, Watt A D, Whittaker J B. Herbivory in global climate change research: direct effects of rising temperature on insect herbivores. Global Change Biology , 2002, 8 (1) : 1–16. DOI:10.1046/j.1365-2486.2002.00451.x |
| [11] | ÖgürE, TuncerC. The effects of global warming on insects. Anadolu Tarım Bilimleri Dergisi , 2011, 26 (1) : 83–90. |
| [12] | 陈瑜, 马春森. 气候变暖对昆虫影响研究进展. 生态学报 , 2010, 30 (8) : 2159–2172. |
| [13] | 严林, 刘振魁, 梅洁人, 兰景华. 野外扣笼条件下草原毛虫对食物的选择. 草地学报 , 1995, 3 (4) : 257–268. |
| [14] | 万秀莲, 张卫国. 草原毛虫幼虫的食性及其空间格局. 草地学报 , 2006, 14 (1) : 84–88. |
| [15] | Yan L, Wang G, Liu C Z. Number of instars and stadium duration of Gynaephora menyuanensis (Lepidoptera: Lymantriidae) from Qinghai-Tibetan Plateau in China. Annals of the Entomological Society of America , 2006, 99 (6) : 1012–1018. DOI:10.1603/0013-8746(2006)99[1012:NOIASD]2.0.CO;2 |
| [16] | 严林. 草原毛虫属的分类、地理分布及门源草原毛虫生活史对策的研究[D]. 兰州: 兰州大学, 2006. http://cdmd.cnki.com.cn/Article/CDMD-10730-2007012889.htm |
| [17] | Klein J A, Harte J, Zhao X Q. Experimental warming, not grazing, decreases rangeland quality on the Tibetan Plateau. Ecological Applications , 2007, 17 (2) : 541–557. DOI:10.1890/05-0685 |
| [18] | 严林, 江小蕾, 王刚. 门源草原毛虫幼虫发育特性的研究. 草业学报 , 2005, 14 (2) : 116–120. |
| [19] | 杨世勇, 宋芬芳, 谢建春. 茉莉酸诱导棉花幼苗抗虫性对棉铃虫相对生长率的影响. 西北农林科技大学学报: 自然科学版 , 2013, 41 (5) : 66–74. |
| [20] | Danks H V. Key themes in the study of seasonal adaptations in insects Ⅱ. Life-cycle patterns. Applied Entomology and Zoology , 2006, 41 (1) : 1–13. DOI:10.1303/aez.2006.1 |
| [21] | Rombough P. Development rate (communication arising): modelling developmental time and temperature. Nature , 2003, 424 (6946) : 268–269. DOI:10.1038/424268a |
| [22] | Chippindale A K, Alipaz J A, Chen H W, Rose M R. Experimental evolution of accelerated development in Drosophila. 1. Developmental speed and larval survival. Evolution , 1997, 51 (5) : 1536–1551. DOI:10.2307/2411206 |
| [23] | Semenza J C, Menne B. Climate change and infectious diseases in Europe. The Lancet Infectious Diseases , 2009, 9 (6) : 365–375. DOI:10.1016/S1473-3099(09)70104-5 |
| [24] | Irlich U M, Terblanche J S, Blackburn T M, Chown S L. Insect rate-temperature relationships: environmental variation and the metabolic theory of ecology. The American Naturalist , 2009, 174 (6) : 819–835. |
| [25] | Atkinson D. Temperature and organism size-a biological law for ectotherms?. Advances in Ecological Research , 1994, 25 : 1–58. DOI:10.1016/S0065-2504(08)60212-3 |
| [26] | Radmacher S, Strohm E. Factors affecting offspring body size in the solitary bee Osmia bicornis (Hymenoptera, Megachilidae). Apidologie , 2010, 41 (2) : 169–177. DOI:10.1051/apido/2009064 |
| [27] | Morewood W D, Ring R A. Revision of the life history of the High Arctic moth Gynaephora groenlandica (Wocke) (Lepidoptera: Lymantriidae). Canadian Journal of Zoology , 1998, 76 (7) : 1371–1381. DOI:10.1139/z98-085 |