 2016, Vol. 36
2016, Vol. 36文章信息
- 齐珂, 樊正球
- QI Ke, FAN Zhengqiu.
- 基于图论的景观连接度量化方法应用研究--以福建省闽清县自然森林为例
- Evaluation method for landscape connectivity based on graph theory: a case study of natural forests in Minqing County, Fujian Province
- 生态学报[J]. 2016, 36(23): 7580-7593
- Acta Ecologica Sinica[J]. 2016, 36(23): 7580-7593
- http://dx.doi.org/10.5846/stxb201507301599
-
文章历史
- 收稿日期: 2015-07-30
- 修订日期: 2016-05-04

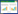




气候变化和人类活动导致的自然生境丧失和生境片断化使全球生物多样性面临威胁, 而有效缓解这类问题的手段之一是维持自然生境的景观连接度[1-5]。大量研究表明, 景观连接度水平对动物活动迁移、植物种子扩散等多项生态过程以及局域物种丰富度产生影响[6-7]。因此, 景观连接度的量化和评价被认为是生物多样性保护领域的一项重要研究课题。景观连接度(Landscape Connectivity)的定义是景观结构对景观内生态流扩散移动的促进或阻碍作用[8], 体现了生态过程对景观格局的响应。景观连接度的量化是以景观斑块空间分布为基础, 结合模式物种扩散能力进行的生境可达性计算[9]。目前景观连接度量化方法主要基于复合种群理论和空间图形理论等生态学和数学理论, 按照分析方法原理可以分为渗透理论法、距离模型法、图论分析法等[10-12]。其中图论分析法随着图论指数的不断发展[13-17]和相关软件的开发使用[18-20]得到越来越广泛的应用[21-26]。
应用图论分析法进行景观连接度研究所需的一个重要参数为距离阈值[27](Threshold Distance, 本文中用TD表示), 距离阈值是生态流的最大可达距离, 用以判断研究区域内生境斑块之间生态流的存在或强弱。最常用的距离阈值取值方法是目标物种法, 即把研究区域内的某个物种作为代表, 以该物种的最大扩散距离作为距离阈值。Szabó等人[28]和Teixeira等人[29]分别将甲虫和蝙蝠的最大移动距离作为距离阈值, Hernández [25]、Liu [24]和陈春娣[30]等人使用植物种子的最大扩散距离作为距离阈值。为了全面了解研究区域中不同扩散能力的物种受到生境片断化的不同影响[16], Xun和Laita等人[31-32]采用了距离梯度法, 即通过列举多个距离阈值将研究区域中物种的扩散能力差异考虑在内。在斑块水平研究连接度时需要使用一个特定的、能够反映景观结构特征的距离阈值[33]。Neel和Urban[33-34]等人依据生态学中“阈值”的意义, 建立“景观连接度指数-距离值”曲线并通过突变点确定距离阈值, 这种方法得到的距离阈值可以体现景观结构特征, 但不一定具有生态意义。景观连接度研究往往作为区域生物多样性保护和制定城市规划的参考, 选用的距离阈值不仅要体现研究区域物种特征, 还应适用于分析景观空间结构, 以上距离阈值取值方法均难以满足研究需求。为了提高景观连接度在区域生物多样性保护和城市规划应用中的科学性, 本研究将试图建立一个适用于多种研究目的的距离阈值综合取值方法。
图论连接度指数依据所使用的连接模型可以被分为二进制连接度指数和概率连接度指数; 按照所指示信息的对象可以分为整体连接度指数和斑块重要性指数。已有研究指出二进制指数适用于对生境斑块空间结构的宏观分析, 概率连接度指数适用于具体的亚种群动态分析[35]。整体连接度指数可以体现区域的连接度水平, 斑块重要性指数用于指示单个斑块在维持区域景观连接度中所起作用的大小[35]。在研究中同时使用多种连接度指数可以获得结构和功能、整体区域和单独斑块的丰富信息, 从而体现使用多种指数的优越性。如何联合使用多种指数, 并正确解读指数的信息也是本研究的重点之一。
本文使用图论分析法对福州市闽清县由亚热带常绿阔叶林和针叶林构成的天然混交林进行森林景观连接研究。通过使用距离阈值取值的综合方法和多种景观连接度指数对闽清县自然保护森林景观连接度进行量化和评价, 并识别研究区域中维持景观连接度的重点区域和关键森林斑块。研究结果推进图论分析法在景观连接度研究方面的应用, 对常绿阔叶林维护和生物多样性保护工作具有参考价值。
1 研究材料与方法 1.1 研究区域与数据准备闽清县位于福建省中部, 福州市西北部, 闽江下游, 距省城福州市50km, 总面积1466km2。地理坐标为北纬25°55′-26°33′, 东经118°30′-119°01′。县内地貌类型复杂多样, 山丘广布, 平原狭小。闽清县属中亚热带季风气候区, 山区植被以林为主, 有杉、松、竹和许多阔叶树种。县内有福建雄江黄楮林国家级自然保护区和自然保护小区57个。
本研究以闽清县生态公益林中的天然森林为研究对象, 基础数据来源是比例尺为1:10000的2010年福州市森林资源二类调查结果的小班数据。该数据基于样地实测和人工数字化, 精度控制在90%以上, 是包含62个小班信息的Shape格式的数据地图文件。使用软件ArcGIS 10.2从数据源中提取出闽清县林分, 将通过小班信息筛选出的起源于天然林的生态公益林小班作为研究区域内的生境斑块。使用ArcGIS 10.2插件Conefor Inputs 10(http://www.conefor.org/gisextensions.html.)提取生境斑块信息并以dbf格式存储, 然后在软件Conefor Sensinode 2.6[35]中完成图论建模和指数计算。筛选得到生境斑块1567个, 斑块总面积168.24km2, 分布情况如图 1所示。
1.2 景观连接度指数与计算按照图论分析法原理, 研究区域的生境斑块与斑块间生态流的格局被抽象为点线图形。两点连线距离小于距离阈值则判定连接存在。景观组分是任意两个斑块之间都直接或间接存在连接的斑块组, 属于不同景观组分的斑块之间不存在生态流[27, 36]。景观组分的格局成为低于景观水平、高于斑块水平的中间层次空间结构。
景观连接度指数是指示景观内生态过程进行的流畅程度的指数。本文采用的图论连接度指数有整体连接度指数NL、NC; 二进制连接度指数H、IIC; 概率连接度指数AWF、PC; 斑块重要性指数dI等4类指数, 所有指数的计算由软件Conefor Sensinode 2.6完成。为了比较不同类型指数的结果, 设定斑块间距离等于距离阈值的情况下扩散概率为0.5[14, 37]。
1.3 距离阈值的取值方法本研究使用目标物种法、距离梯度法、拟合曲线法依次对距离阈值取值。
(1)目标物种法根据林业小班调查的树种信息, 以硬阔为优势树种的小班有1318个, 占全部小班的84.11%。以马尾松(Pinus massoniana)或杉木(Cunninghamia lanceolata)为优势树种的小班次之, 有121个和75个, 分别占总数的7.72%和4.79%。根据亚洲东部常绿阔叶林和福建中亚热带阔叶林物种特点[39-40], 选取壳斗科硬叶栎类树种栓皮栎(Quercus variabilis)作为研究区域的目标种。根据Xiao[41]等人研究, 栓皮栎在自然生境下依靠啮齿动物扩散种子, 最大扩散距离为42.5m。

|
| 图 1 闽清县自然保护森林分布 Fig. 1 The conservation natural forest in Minqing County |
| 指数类型 Index type |
指数 Index |
公式 Formula |
描述 Description and interpretation |
| 整体指数 | 区域连接总数NL | 研究区域内斑块之间存在的所有连接的个数 | |
| Overall indices | 景观组分总数NC | 研究区域内斑块划分出的景观组分的个数 | |
| 二进制连接度指数 Binary indices |
Harary指数H[38] | 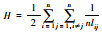
|
|
| 可能性连接度指数 Probability indices |
整体连接度指数IIC[13] | 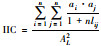
|
n:研究区域中斑块总数; nlij:斑块i和斑块j之间存在的最少连接数, 在不存在连接的斑块之间nlij=∞; pij:斑块i与斑块j之间直接存在扩散的概率; P·ij:斑块i和斑块j之间各个扩散途径的最大概率值; ai/aj:斑块i/斑块j的属性值, 一般取值为斑块面积; AL:研究区域的总属性值, 当斑块属性值取斑块面积时, AL为研究区域总面积。研究区域连接度越高, H、IIC、AWF、PC的值越大 |
| 面积权重指数AWF[27] | 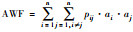
|
||
| 可能性连接度指数PC[14] | 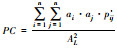
|
||
| 斑块重要性指数 Patch importance indices |
斑块重要性指数dI[14] | 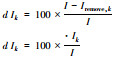
|
dIk:连接度指数I对应的斑块k的重要性指数; I:研究区域原本的连接度指数值; Iremove, k:去除斑块k后的研究区域的连接性指数值dIk指示单个斑块对区域整体连接度贡献大小, 用以对斑块进行重要性识别和排序 |
(2)距离梯度法根据Corlett[42]对亚洲森林植物种子传播方式和传播距离的研究, 可以将植物种子的传播距离分为5个级别。表 2中的前4个传播类型适用于研究区域植被, 因此选用10m、100m、1km、10km作为距离阈值以体现研究区域内不同植物物种的扩散能力。
| 传播距离 Distance |
传播方式 Dispersal method |
描述 Descriptions |
| 0-10m | 主动传播、动物传播 | 以自身机械力主动传播或依靠蚁类被动传播 |
| 10-100m | 风力传播、动物传播 | 依靠风力传播(有翅种子)或啮齿类等小型动物传播 |
| 100m-1km | 动物传播 | 依靠动物传播, 包括中小型鸟类和哺乳类(如果蝠、大多数灵长类) |
| 1-10km | 动物传播 | 依靠活动范围较广的动物(如大多数陆生草食动物、广生境鸟类)传播 |
| > 10km | 风力传播、水力传播、动物传播 | 传播方式既有风力、水力等非生物传播方式, 也有包括大型哺乳动物(如大象、犀牛)的捕食搬运和人类活动传播 |
| 表格来源:Corlett[42], 由作者翻译 | ||
(3)拟合曲线法比照Urban和Rubio等人研究[34, 43]选取10m-30km共40个距离值计算NL、NC、H、IIC、AWF和PC等连接度指数。对40个距离值和相应的H、IIC、AWF、PC指数值双向取自然对数。使用SPSS Statistics 17分析模块的曲线估计进行4条“景观连接度指数-距离值”曲线拟合, 对拟合得到的曲线求导数以找出突变点, 计算突变点的距离值。通过分析NL、NC指数计算结果和研究区域的实际情况对突变点距离值进行取舍, 以确保取值具有生态意义。
2 结果与分析 2.1 研究区域斑块特征研究区域内有1567个斑块, 斑块总面积168.24km2, 占总面积的11.45%。根据景观特征, 将斑块面积划分为5个等级, 分别为小型斑块(>0-5 hm2)、中型斑块(>5-10 hm2)、大型斑块(>10-20 hm2)、较大性斑块(>20-30 hm2)、巨型斑块(>30 hm2)。由表 3和图 2可知, 小型、中型和大型斑块的数量均大于较大型斑块和巨型斑块, 其中大型斑块数量最多。各类型斑块的总面积比例呈近似正态分布, 大型斑块具有最大的总面积。小型、中型斑块数量超过总斑块数的一半但面积仅占26.75%, 较大型、巨型斑块数量占总数的11.74%而面积占比接近30%。由此可知研究区域主要斑块类型为面积为10-20 hm2的大型斑块, 整体呈现大量小面积斑块分散分布于少量大面积斑块之间的格局。
| 斑块类型 Type of Patches |
面积范围 Area range /hm2 |
斑块数量 No of patches |
数量占比 Proportions of Patch No/% |
斑块面积 Area of patches/hm2 |
面积占比 Proportions of patch area/% |
平均斑块面积 Mean area/hm2 |
| 小型斑块 Small Patches |
0-5 | 394 | 25.14 | 1112.31 | 6.61 | 2.82 |
| 中型斑块 Medium Patches |
5-10 | 455 | 29.04 | 3388.61 | 20.14 | 7.45 |
| 大型斑块 Big Patches |
10-20 | 534 | 34.08 | 7594.67 | 45.14 | 14.22 |
| 较大型斑块 Large Patches |
20-30 | 158 | 10.08 | 3816.35 | 22.68 | 24.15 |
| 巨型斑块 Huge Patches |
> 30 | 26 | 1.66 | 912.12 | 5.42 | 35.08 |

|
| 图 2 各斑块类型的数量与面积占比 Fig. 2 Proportions of patch number and patch area of each type |
依次按照目标物种法、距离梯度法、拟合曲线法取值得到多个距离阈值。根据已有研究中栓皮栎种子最大扩散距离为42.5m, 选取50m作为目标物种法确定的距离阈值。使用距离梯度法选取10m、100m、1km、10km作为代表研究区域内不同扩散方式的植物种类的距离阈值。考虑到还有12.51%数量的小班以依靠风力传种的松科或杉科植物为优势树种, 借鉴Nathan等人[44]对异叶铁杉(Tsuga heterophylla)和花旗松(Pseudotsuga menziesii)的有翅种子依靠风力长距离传播研究中最大扩散距离为800-1600m的研究结果, 选取1500m作为目标物种法和距离梯度法的补充距离阈值。
使用拟合曲线法过程中得到的“景观连接度指数-距离值”拟合曲线如图 3, 方程见表 4。由于变量距离阈值数量级跨度较大, 本研究采取对指数值和距离值分别取自然对数后再进行曲线估计回归分析的做法。拟合曲线方程中指数值突变(即斜率最高的)点处的距离值分别为974、916、986m和781m, 基本接近于1km。根据NL和NC的计算结果(图 4)对得到的距离值进行检验和取舍。由图 4可知当距离阈值为5km时NC降至最小值1, 说明当距离阈值达到5km时, 研究区域中只存在一个景观组分, 所有斑块都可以直接或间接相互联系。当距离阈值为12km时, NL达到最大值369395, 即在距离阈值超过12km后所有的二进制指数达到最大值。为了尽可能反映景观结构特征, 距离阈值应在5km以内选择。使用曲线拟合法取到的1km距离阈值符合这个条件。

|
| 图 3 景观连接度指数H、IIC、AWF、PC的拟合曲线和突变点 Fig. 3 Curves and changing points of H, IIC, AWF and PC |
| 指数 Indices |
曲线公式 Curve fitting equations |
R2 | Sig. | F | 突变点x值 Change point |
突变点对应距离值 Distance/m |
| H | y=-0.0272x3-0.0021 x2+1.0007x+11.317 | 0.9962 | 0.000 | 3119.98 | -0.0257 | 974 |
| IIC | y=-0.0253x3-0.0067 x2+0.8701x-7.1597 | 0.9946 | 0.000 | 2200.59 | -0.0883 | 916 |
| AWF | y=-0.0258x3-0.0011 x2+0.9874x+34.342 | 0.9973 | 0.000 | 4419.35 | -0.0142 | 986 |
| PC | y=-0.0162x3-0.012 x2+0.6286x-5.8164 | 0.9985 | 0.000 | 8240.33 | -0.2469 | 781 |

|
| 图 4 指数NC和NL的计算结果 Fig. 4 Results of NL and NC |
代表目标物种和不同扩散方式树种的6个距离阈值10m、50m、100m、1km、1.5km、10km分别对应的NC值为349、302、242、56、33、1。除去将景观组分划得过于细碎的距离阈值10m和几乎没有划分出景观组分的距离阈值10km, 其余4个距离阈值的景观组分划分情况如图 5所示。其中包含斑块数量最多的前10个景观组分以彩色标出。由图可知当距离阈值小于1km时邻近的斑块被划分到不同组分, 研究区域内景观组分分布过于分散; 当距离阈值大于1km时原本相距较远的斑块被划分在了一起, 不符合实际情况, 这2种情况都不适合于挖掘景观结构特征[23]。当距离阈值取1km时, 研究区域出现了明确的主次景观组分, 适于景观结构分析。综上所述, 使用1km作为距离阈值分析景观连接度的结构特征最为合适。

|
| 图 5 不同距离阈值下景观组分划分情况平面图 Fig. 5 Component distribution and prioritization at different TD 从左上到右下分别表示距离阈值为50m、100m、1km和1.5km时的景观组分划分情况; 每幅图中具有最多斑块数量的10个景观组分被用彩色标出 |
由代表区域优势物种的50m、1.5km和代表植物不同扩散能力的10m、100m、1km、10km组成用以分析研究区域整体连接度水平的距离阈值梯度。表 5显示了在各个距离阈值下的景观连接度指数值。其中所有IIC值都低于PC值并且二者都低于0.01, 说明研究区域景观连接度水平较低。随距离阈值的增加所有连接度指数值上升, 说明扩散能力越强的物种受到景观破碎化的影响越小。几乎所有指数在1km距离阈值处有显著提高, 说明距离阈值达到1km左右时斑块之间存在连接的数量显著增多, 并出现了较清晰的景观组分结构。
| 连接度指数 Connectivity indices |
距离阈值 Threshold distance |
|||||
| 10m | 50m | 100m | 1000m | 1500m | 10000m | |
| H | 8433 | 9819 | 11745 | 85122 | 120312 | 642684 |
| IIC | 1.11×10-4 | 1.24×10-4 | 1.43×10-4 | 8.42×10-4 | 1.14×10-3 | 4.375×10-3 |
| AWF | 8.30×1013 | 9.69×1013 | 1.19×1014 | 7.66×1014 | 1.23×1015 | 6.01×1015 |
| PC | 5.56×10-4 | 6.71×10-4 | 8.07×10-4 | 3.03×10-3 | 4.01×10-3 | 9.77×10-3 |
以景观组分为结构单位探究景观结构与景观连接度的关系。将距离阈值取1km的情景下划分出的56个景观组分按照软件Conefor Sensinode 2.6输出顺序命名为C1-C56, 分别计算景观组分的斑块数量、斑块面积、斑块重要性指数dIIC和dPC的总值并排序, 各排序结果的前10组分见表 6。斑块数量和斑块面积的排序结果相近, 所有排序第1、2位的景观组分均为C3、C24。其中C3的斑块数量和斑块面积明显高于其他组分, 斑块重要性指数总值占所有斑块的90%以上。此外C3的单位面积的重要性指数值也明显高于其他景观组分。因此, 从结构和功能两方面看, C3是研究区域中最主要的景观组分, 占据整个区域的关键位置(图 5)并行使了绝大部分的连接度功能。其他的景观组分处于次要地位, 对维持整体景观连接度的贡献有限。
| 排序 Rank |
按斑块数量Rank by No of patches | 按斑块面积Rank by area of patches | 按景观组分∑dIIC值Rank by ∑dIIC | 按景观组分∑dPC值Rank by ∑dPC | |||||||||||||
| 景观组分 Component |
斑块数量 No of patch |
比例 Rate/% |
景观组分 Component |
斑块面积 Sum of patch area/hm2 |
比例 Rate/% |
景观组分 Component |
景观组分dIIC值 Sum of dIIC |
比例 Rate/% |
单位面积dIIC值 dIIC per area unit |
景观组分 Component |
景观组分dPC值 Sum of dPC |
比例 Rate/% |
单位面积dPC值 dPC per area unit |
||||
| 1 | C3 | 1014 | 64.71 | C3 | 11857 | 70.50 | C3 | 206.55 | 93.81 | 1.74 | C3 | 263.78 | 92.48 | 2.23 | |||
| 2 | C24 | 173 | 11.04 | C24 | 2044 | 12.15 | C24 | 10.97 | 4.98 | 0.54 | C24 | 13.98 | 4.90 | 0.68 | |||
| 3 | C1 | 122 | 7.79 | C1 | 606 | 3.61 | C1 | 1.24 | 0.56 | 0.20 | C50 | 1.32 | 0.46 | 0.55 | |||
| 4 | C23 | 22 | 1.40 | C23 | 252 | 1.50 | C23 | 0.31 | 0.14 | 0.12 | C1 | 1.25 | 0.44 | 0.21 | |||
| 5 | C44 | 21 | 1.34 | C50 | 239 | 1.42 | C50 | 0.28 | 0.13 | 0.12 | C39 | 1.14 | 0.40 | 1.20 | |||
| 6 | C12 | 20 | 1.28 | C5 | 166 | 0.99 | C5 | 0.14 | 0.06 | 0.08 | C41 | 1.05 | 0.37 | 0.81 | |||
| 7 | C19 | 17 | 1.08 | C12 | 150 | 0.89 | C12 | 0.11 | 0.05 | 0.07 | C44 | 0.47 | 0.17 | 0.45 | |||
| 8 | C4 | 15 | 0.96 | C19 | 146 | 0.87 | C19 | 0.11 | 0.05 | 0.07 | C5 | 0.42 | 0.15 | 0.25 | |||
| 9 | C50 | 14 | 0.89 | C41 | 129 | 0.77 | C41 | 0.09 | 0.04 | 0.07 | C23 | 0.32 | 0.11 | 0.13 | |||
| 10 | C2 | 13 | 0.83 | C4 | 112 | 0.66 | C4 | 0.06 | 0.03 | 0.06 | C40 | 0.30 | 0.11 | 0.43 | |||
为分析重要区域内的斑块重要性和空间分布特征, 对景观组分C3内的1014个斑块进行重要性指数值排序。按软件默认顺序将C3内斑块命名为P1-P1014, 所有斑块的重要性指数dIIC和dPC值见图 6。大部分斑块的dIIC和dPC值都很低, 只有极少数斑块的重要性明显高于其他斑块。dIIC和dPC的最大值分别为3.15和3.79, 与其他研究中dI值可能达到10以上的情况相比偏低, 这说明在本研究区域内没有能够特别显著影响整体连接度水平的斑块。从C3中筛选出具有最高dIIC或dPC值的10个斑块, 分布位置如图 7(C)和(D)。斑块P497、P787、P780同时出现在dIIC和dPC的前10中, 说明这3个斑块的重要性较高。具有最高dIIC或dPC值的10个斑块相对于普通斑块能够对整体连接度水平产生更大影响, 是需要重点保护和维持的区域。

|
| 图 6 C3组内所有斑块的dIIC、dPC值与对应的斑块ID(距离阈值1km) Fig. 6 Patch importance values of dIIC and dPC and corresponding patch IDs in C3 at TD of 1km |

|
| 图 7 C3组内斑块重要性分级结果分和具有最高dI值的前10名斑块空间分布图(距离阈值1km) Fig. 7 Distribution of each patch importance level and top 10 patches of dIIC or dPC in C3 at TD of 1km |
划分C3内斑块的dIIC和dPC值为包括极高重要值(dI≥1), 高重要值(0.5≤dI < 1), 中等重要值(0.1≤dI < 0.5)和低重要值(0≤dI < 0.1)等4个级别。某个低重要值斑的缺失对整体连接度水平的影响不超过0.1%, 如果缺失某个极高重要值斑块则将会导致区域整体连接度水平降低1%以上。具有最高dIIC值的10个斑块中有6个具有极高重要值, 而具有最高dPC值的10个斑块均具有极高重要值。图 7体现了斑块重要性指数值的空间分布趋势, 其中(A)和(B)显示了大部分斑块具有中等重要值, 低重要值斑块处于C3的边缘区域, 高重要值和极高重要值斑块被中等重要值斑块包围。两个指数的低重要值和中等重要值区域形状相似, 但dPC的计算结果中有更多高重要值斑块, 并且这些斑块的空间分布呈现类似廊道的形态。
为了探究斑块重要性指数值是否与斑块面积存在正比关系, 分别在按dIIC或dPC划分出的极高重要值斑块范围、景观组分C3范围和研究区域范围等3个层次进行相关性检验。由表 7可知, dIIC和dPC值在景观组分C3范围和研究区域范围内与斑块面积呈显著正相关关系, 说明斑块面积在一定程度上决定了斑块重要性指数的大小。但是在2组极高重要值斑块范围的检验结果出现了Pearson相关性指数为负和不显著的情况。说明对这些具有极高重要值的斑块来说, 决定其重要值的最主要因素不只是斑块面积, 而是包括斑块空间位置在内的其他斑块属性。景观组分C3内面积最大的前10个斑块(P497、P37、P311、P119、P142、P79、P90、P622、P139)中只有P497和P311进入了dIIC或dPC值的排名的前10位, 这也证明了斑块重要性值并不完全取决于斑块面积的结论。
| 斑块重要性指数 Patch importance indices |
研究区域全部斑块 All patches |
Sig. | 景观组分C3斑块 Patches in C3 |
Sig. | 极高重要值斑块 Key patches |
Sig. |
| dIIC | 0.540** | 0.000 | 0.587** | 0.000 | -0.126 | 0.406 |
| dPC | 0.465** | 0.000 | 0.482** | 0.000 | 0.282 | 0.056 |
| **表示在0.01水平(单侧)上显著相关 | ||||||
目前使用的距离阈值取值方法各有局限性。目标物种法的局限性在于物种的最大扩散距离不易准确测定, 在开展研究之前实地测定研究区域生境下目标物种的最大扩散距离也难以实现。虽然可以参考已有研究对物种扩散距离的测定结果, 但是即使参考同一物种的相关研究仍然存在生境条件差异造成的误差。植物种子传播最大距离受到风力、传粉者等因素影响; 动物的最大扩散距离会因为物种所处的生活阶段(如迁徙、捕食等)和观察时间周期等因素不同而产生差异[45]。本研究中选取的目标物种栓皮栎的种子扩散主要依靠啮齿动物的搬运, 受当地啮齿动物的行为特征和种群大小的影响。另一目标物种马尾松依靠风力播种, 种子扩散距离受当地的地势地貌、风向风速等因素影响[46]。另外在无法获取目标物种扩散距离的研究成果时, 可以参考具有相似种子特征和传播方式的物种在相似环境条件下研究结论, 如本研究借鉴了异叶铁杉和花旗松种子传播的研究结果。因此目标物种法得到的距离值作为距离阈值取值的参考更为合适。距离梯度法[47]避免了选取单一物种可能导致的片面性和误差, 但是不能为景观结构分析选取一个具体距离值作为最优距离阈值, 使研究只能停留在宏观层面。本研究使用距离梯度法确定了4个距离阈值, 实现了针对不同扩散能力物种的景观连接度比较研究。Neel、Urban和Shanthala[23, 33-34]等人认为距离阈值应当是使整体连接度水平发生突变的距离值, 虽然可以使用拟合曲线法得到但不能在所有研究中实现[15], 而且需要检验取值的生态意义。本研究使用H、IIC、AWF和PC等连接度指数的计算结果拟合求得4个接近于1km的突变点距离值, 结合景观组分划分平面图和整体连接性指数NL、NC值确定了最优距离阈值为1km, 并使用该距离阈值分析了研究区域的景观结构与景观连接度。
本研究采用了由目标物种法、距离梯度法、拟合曲线法3个步骤组成的距离阈值综合取值方法, 得到的距离阈值既可用于全面评价一个区域的景观连接度水平又能满足区域保护规划等研究需求。从方法的效率上看, 第三步拟合曲线法能够避免第一步目标物种法可能造成的误差, 但需要结合第一、二步的取值和研究区域实际情况取舍计算结果。第二步距离梯度法列举出的距离阈值可以用于整体连接度评价, 第三步拟合曲线时要用到的备选距离值必然包含第一、二步选取的距离值, 因此优选阈值的准备工作和整体连接度评价指数的计算可以同时完成, 节省了操作和时间。从取值结果来看, 最优距离阈值1km的生态学意义是风力传播和动物传播物种的最大扩散距离, 代表了研究区域中植物物种的中等扩散能力。以中等扩散能力物种为对象分析研究区域的景观结构与景观连接度水平之间关系具有一定合理性。综上所述, 本研究提出的距离阈值综合取值方法是一种包含信息全面、取值结果可靠、操作省时高效并且可以同时满足多种研究目的的距离阈值取值方法。
3.2 指数的联合使用与比较本研究使用景观连接度指数与斑块重要性指数、二进制连接度指数与概率连接度指数对研究区域的景观连接度进行了多尺度、多角度的分析。其中景观连接度指数IIC和PC用于评价研究区域整体景观连接度水平, 斑块重要性指数dIIC和dPC用于指示单个斑块对景观连接度的贡献量。结合使用这两类指数可以使研究层次丰富, 增加结果的应用性。从斑块之间连接的量化原理分类, 本研究使用了二进制连接度指数NL、NC、H、IIC和概率连接度指数AWF、PC等两类指数。二进制模型将斑块间连接判断为“有”和“无”, 适用于划分景观结构; 概率模型将连接赋值为扩散存在的概率, 适用于比较连接的强弱。
软件Conefor Sensinode 2.6的开发者Pascual-Hortal和Saura认为dIIC具有对景观格局变化敏感的优点[13], 但在斑块功能分析方面dPC更有优势[15]。表 8比较了分别按照dIIC值和dPC值对景观组分C3内斑块的重要性分级情况, 在表的右上角区域, 按dPC划分的级别高于按dIIC划分的级别, 而表的左下角为相反情况。从合计看, 除了极高值级别斑块的数量存在明显差异外, 按两个指数的划分结果里其他重要性等级斑块的数量差异不超过20。值得注意的是, 同时被dIIC和dPC划分为极高重要值等级的斑块仅有1个, 由dIIC值划分为极高重要值的6个斑块中有5个按照dPC值划分为低重要值或中等重要值斑块, 而按dPC划分出的33个极高重要值斑块中有24个斑块具有的dIIC值仅为中等重要值级别。这些差异说明了dIIC和dPC从不同角度量化斑块重要性。由于计算二进制连接度指数dIIC的过程损失了大部分斑块间距离的信息, 而概率连接度指数dPC的值与斑块间距离一一对应, 因此dIIC指数主要反映景观结构信息,对距离阈值变化的响应更迅速, 对有助于维持景观结构完整的斑块赋予高重要性。而dPC指数包含了更多斑块空间位置的信息,对有助于维持区域生态流流动的斑块赋予高重要性。从图 7(B)可以看到具有极高重要级别dPC值的斑块大多处于研究区域的关键位置并行使了踏脚石功能。综上, 联合使用dIIC和dPC可以得到有不同的侧重点的斑块重要性结果, 并能加深对研究区域斑块特征的理解。
| 按dPC划分重要性等级 Patch important level classified by dPC |
||||||
| 低重要等级 Low importance |
中等重要等级 Medium importance |
高重要等级 High importance |
极高重要等级 Very high importance |
合计 Total |
||
| 按dIIC划分 | 低重要等级 | 248 | 45 | 3 | 1 | 297 |
| 重要性等级 | 中等重要等级 | 26 | 598 | 25 | 24 | 673 |
| Patch important level | 高重要等级 | 0 | 12 | 19 | 7 | 38 |
| classified by dIIC | 极高重要等级 | 3 | 2 | 0 | 1 | 6 |
| 合计 | 277 | 657 | 47 | 33 | 1014 | |
(1)本研究提出的距离阈值综合取值方法是一种具有广泛适用性的距离阈值取值方法。相比目标物种法、距离梯度法、拟合曲线法等单一方法, 综合方法取值更具准确性和科学性, 并能同时满足多种研究目的。
(2)二进制连接度指数具有划分景观结构的重要功能, 但在计算斑块重要性时难以体现斑块空间位置信息。概率连接度指数的计算过程中模拟生态流流量, 对行使疏导生态流功能的斑块赋予更高重要性。联合使用两类指数可以更全面理解研究区域的景观结构和斑块重要性格局。
(3)闽清县自然森林斑块分散度较高, 景观连接度水平较低, 主要景观组分C3是整个区域生态流流动最频繁的区域。在研究区域内面积越大的斑块重要性指数值越高, 但是发挥关键性作用的极高重要值斑块的重要性指数值并不完全取决于斑块面积, 而更多的与斑块位置等因素相关。据此结论, 建议在制定区域物种保护规划时在关键位置增加高质量斑块以起到促进和维持生境景观连接度的作用, 不仅节省用地空间还能获得较好的保护成效。
本研究的局限性在于忽略了闽清县周边县份的自然森林, 可能导致低估位于研究区域边缘斑块的重要性。另外本研究沿用了其他研究中将斑块面积作为软件Conefor Sensinode 2.6中斑块属性(Node Attribute)值的做法, 但从本研究的结论可知斑块面积不能完全代表生境斑块的属性, 因此改进斑块属性的赋值方法是景观连接度研究中亟需解决的问题。
| [1] | Lindenmayer D B, Fischer J. Tackling the habitat fragmentation panchreston. Trends in Ecology & Evolution , 2007, 22 (3) : 127–132. |
| [2] | Brooks C P. Quantifying population substructure:extending the graph-theoretic approach. Ecology , 2006, 87 (4) : 864–872. DOI:10.1890/05-0860 |
| [3] | D'Eon R G, Glenn S M, Parfitt I, Fortin M J. Landscape connectivity as a function of scale and organism vagility in a real forested landscape. Conservation Ecology , 2002, 6 (2) : 10–10. |
| [4] | Martensen A C, Pimentel R G, Metzger J P. Relative effects of fragment size and connectivity on bird community in the Atlantic Rain Forest:implications for conservation. Biological Conservation , 2008, 141 (9) : 2184–2192. DOI:10.1016/j.biocon.2008.06.008 |
| [5] | 岳天祥. 生物多样性研究及其问题. 生态学报 , 2001, 21 (3) : 462–467. |
| [6] | Jesus F M, Pivello V R, Meirelles S T, Franco G A D C, Metzger J P. The importance of landscape structure for seed dispersal in rain forest fragments. Journal of Vegetation Science , 2012, 23 (6) : 1126–1136. DOI:10.1111/jvs.2012.23.issue-6 |
| [7] | Martensen A C, Ribeiro M C, Banks-Leite C, Prado P I, Metzger J P. Associations of forest cover, fragment area, and connectivity with Neotropical understory bird species richness and abundance. Conservation Biology , 2012, 26 (6) : 1100–1111. DOI:10.1111/j.1523-1739.2012.01940.x |
| [8] | Taylor P D, Fahrig L, Henein K, Merriam G. Connectivity is a vital element of landscape structure. Oikos , 1993, 68 (3) : 571–573. DOI:10.2307/3544927 |
| [9] | Tischendorf L, Fahrig L. On the usage and measurement of landscape connectivity. Oikos , 2000, 90 (1) : 7–9. DOI:10.1034/j.1600-0706.2000.900102.x |
| [10] | Kindlmann P, Burel F. Connectivity measures:a review. Landscape Ecology , 2008, 23 (8) : 879–890. |
| [11] | 富伟, 刘世梁, 崔保山, 张兆苓. 景观生态学中生态连接度研究进展. 生态学报 , 2009, 29 (11) : 6174–6182. |
| [12] | 吴昌广, 周志翔, 王鹏程, 肖文发, 滕明君. 景观连接度的概念、度量及其应用. 生态学报 , 2010, 30 (7) : 1903–1910. |
| [13] | Pascual-Hortal L, Saura S. Comparison and development of new graph-based landscape connectivity indices:towards the priorization of habitat patches and corridors for conservation. Landscape Ecology , 2006, 21 (7) : 959–967. DOI:10.1007/s10980-006-0013-z |
| [14] | Saura S, Pascual-Hortal L. A new habitat availability index to integrate connectivity in landscape conservation planning:comparison with existing indices and application to a case study. Landscape and Urban Planning , 2007, 83 (2/3) : 91–103. |
| [15] | Saura S, Rubio L. A common currency for the different ways in which patches and links can contribute to habitat availability and connectivity in the landscape. Ecography , 2010, 33 (3) : 523–537. |
| [16] | Saura S, Estreguil C, Mouton C, Rodríguez-Freire M. Network analysis to assess landscape connectivity trends:application to European forests (1990-2000). Ecological Indicators , 2011, 11 (2) : 407–416. DOI:10.1016/j.ecolind.2010.06.011 |
| [17] | Baranyi G, Saura S, Podani J, Jordán F. Contribution of habitat patches to network connectivity:redundancy and uniqueness of topological indices. Ecological Indicators , 2011, 11 (5) : 1301–1310. DOI:10.1016/j.ecolind.2011.02.003 |
| [18] | Ray N. PATHMATRIX:a geographical information system tool to compute effective distances among samples. Molecular Ecology Notes , 2005, 5 (1) : 177–180. DOI:10.1111/men.2005.5.issue-1 |
| [19] | Saura S, Torné J. Conefor Sensinode 2.2:a software package for quantifying the importance of habitat patches for landscape connectivity. Environmental Modelling & Software , 2009, 24 (1) : 135–139. |
| [20] | Foltête J C, Clauzel C, Vuidel G. A software tool dedicated to the modelling of landscape networks. Environmental Modelling & Software , 2012, 38 : 316–327. |
| [21] | Ernst B W. Quantifying connectivity using graph based connectivity response curves in complex landscapes under simulated forest management scenarios. Forest Ecology and Management , 2014, 321 : 94–104. DOI:10.1016/j.foreco.2013.07.033 |
| [22] | O'Brien D, Manseau M, Fall A, Fortin M J. Testing the importance of spatial configuration of winter habitat for woodland caribou:an application of graph theory. Biological Conservation , 2006, 130 (1) : 70–83. DOI:10.1016/j.biocon.2005.12.014 |
| [23] | Shanthala Devi B S, Murthy M S R, Debnath B, Jha C S. Forest patch connectivity diagnostics and prioritization using graph theory. Ecological Modelling , 2013, 251 : 279–287. DOI:10.1016/j.ecolmodel.2012.12.022 |
| [24] | Liu S L, Deng L, Dong S K, Zhao Q H, Yang J J, Wang C. Landscape connectivity dynamics based on network analysis in the Xishuangbanna Nature Reserve, China. Acta Oecologica , 2014, 55 : 66–77. DOI:10.1016/j.actao.2013.12.001 |
| [25] | Hernández A, Miranda M, Arellano E C, Saura S, Ovalle C. Landscape dynamics and their effect on the functional connectivity of a Mediterranean landscape in Chile. Ecological Indicators , 2015, 48 : 198–206. DOI:10.1016/j.ecolind.2014.08.010 |
| [26] | 陈杰, 梁国付, 丁圣彦. 基于景观连接度的森林景观恢复研究--以巩义市为例. 生态学报 , 2012, 32 (12) : 3773–3781. |
| [27] | Bunn A G, Urban D L, Keitt T H. Landscape connectivity:a conservation application of graph theory. Journal of Environmental Management , 2000, 59 (4) : 265–278. DOI:10.1006/jema.2000.0373 |
| [28] | Szabó S, Novák T, Elek Z. Distance models in ecological network management:a case study of patch connectivity in a grassland network. Journal for Nature Conservation , 2012, 20 (5) : 293–300. DOI:10.1016/j.jnc.2012.06.002 |
| [29] | Teixeira T S M, Weber M M, Dias D, Lorini M L, Esbérard C E L, Novaes R L M, Cerqueira R, Vale M M. Combining environmental suitability and habitat connectivity to map rare or Data Deficient species in the Tropics. Journal for Nature Conservation , 2014, 22 (4) : 384–390. DOI:10.1016/j.jnc.2014.04.001 |
| [30] | 陈春娣, ColinM D, MariaI E, GlennS H, 吴胜军. 城市生态网络功能性连接辨识方法. 生态学报 , 2015, 35 (19) : 6414–6424. |
| [31] | Laita A, Mönkkönen M, Kotiaho J S. Woodland key habitats evaluated as part of a functional reserve network. Biological Conservation , 2010, 143 (5) : 1212–1227. DOI:10.1016/j.biocon.2010.02.029 |
| [32] | Xun B, Yu D Y, Liu Y P. Habitat connectivity analysis for conservation implications in an urban area. Acta Ecologica Sinica , 2014, 34 (1) : 44–52. DOI:10.1016/j.chnaes.2013.11.006 |
| [33] | Neel M C. Patch connectivity and genetic diversity conservation in the federally endangered and narrowly endemic plant species Astragalus albens (Fabaceae). Biological Conservation , 2008, 141 (4) : 938–955. DOI:10.1016/j.biocon.2007.12.031 |
| [34] | Keitt T H, Urban D L, Milne B T. Detecting critical scales in fragmented landscapes. Ecology and Society , 1997, 1 (1) : 4. |
| [35] | Bodin Ö, Saura S. Ranking individual habitat patches as connectivity providers:integrating network analysis and patch removal experiments. Ecological Modelling , 2010, 221 (19) : 2393–2405. DOI:10.1016/j.ecolmodel.2010.06.017 |
| [36] | Urban D, Keitt T. Landscape connectivity:a graph-theoretic perspective. Ecology , 2001, 82 (5) : 1205–1218. DOI:10.1890/0012-9658(2001)082[1205:LCAGTP]2.0.CO;2 |
| [37] | 熊春妮, 魏虹, 兰明娟. 重庆市都市区绿地景观的连通性. 生态学报 , 2008, 28 (5) : 2237–2244. |
| [38] | Ricotta C, Stanisci A, Avena G C, Blasi C. Quantifying the network connectivity of landscape mosaics:a graph-theoretical approach. Community Ecology , 2000, 1 (1) : 89–94. DOI:10.1556/ComEc.1.2000.1.12 |
| [39] | 李昌华. 亚洲东部常绿阔叶林的分布. 自然资源 , 1997 (2) : 37–45. |
| [40] | 廖成章, 洪伟, 吴承祯, 王新功, 程煜, 封磊. 福建中亚热带常绿阔叶林物种多样性的空间格局. 广西植物 , 2003, 23 (6) : 517–522. |
| [41] | Xiao Z S, Jansen P A, Zhang Z B. Using seed-tagging methods for assessing post-dispersal seed fate in rodent-dispersed trees. Forest Ecology and Management , 2006, 223 (1/3) : 18–23. |
| [42] | Corlett R T. Seed dispersal distances and plant migration potential in tropical East Asia. Biotropica , 2009, 41 (5) : 592–598. DOI:10.1111/j.1744-7429.2009.00503.x |
| [43] | Rubio L, Saura S. Assessing the importance of individual habitat patches as irreplaceable connecting elements:an analysis of simulated and real landscape data. Ecological Complexity , 2012, 11 : 28–37. DOI:10.1016/j.ecocom.2012.01.003 |
| [44] | Nathan R, Katul G G, Horn H S, Thomas S M, Oren R, Avissar R, Pacala S W, Levin S A. Mechanisms of long-distance dispersal of seeds by wind. Nature , 2002, 418 (6896) : 409–413. DOI:10.1038/nature00844 |
| [45] | Zetterberg A, Mörtberg U M, Balfors B. Making graph theory operational for landscape ecological assessments, planning, and design. Landscape and Urban Planning , 2010, 95 (4) : 181–191. DOI:10.1016/j.landurbplan.2010.01.002 |
| [46] | Bullock J M, Clarke R T. Long distance seed dispersal by wind:measuring and modelling the tail of the curve. Oecologia , 2000, 124 (4) : 506–521. DOI:10.1007/PL00008876 |
| [47] | Blazquez-Cabrera S, Bodin Ö, Saura S. Indicators of the impacts of habitat loss on connectivity and related conservation priorities:do they change when habitat patches are defined at different scales. Ecological Indicators , 2014, 45 : 704–716. DOI:10.1016/j.ecolind.2014.05.028 |