 2016, Vol. 36
2016, Vol. 36文章信息
- 靳明, 叶碧碧, 庞燕, 储昭升, 杨琦
- JIN Ming, YE Bibi, PANG Yan, CHU Zhaosheng, YANG Qi
- 濒危种螺蛳对典型附着藻的摄食特性及其在洱海流域恢复的可行性
- Study on Margarya melanioides' feeding characteristics towards four attached algae and its potential restoration in Erhai basin
- 生态学报, 2016, 36(2): 518-524
- Acta Ecologica Sinica, 2016, 36(2): 518-524
- http://dx.doi.org/10.5846/stxb201402240314
-
文章历史
- 收稿日期: 2014-02-22
- 网络出版日期: 2015-06-08






2. 中国地质大学(北京), 水资源与环境学院, 北京 100083
2. China University of Geosciences, School of Water Resources & Environment, Beijing 100083, China
螺类是淡水生态系统中重要的底栖动物之一,广泛存在于沟渠、池塘、河流以及湖泊中,它们可以加速水底碎屑的分解,促进泥水界面的物质交换和水体的自净,在生态系统物质循环和能量流动中起着重要作用[1, 2];Chaparro等人研究指出腹足类软体动物能通过呼吸过程中分泌的黏液将水中漂浮颗粒物沉降并通过舌齿进行摄食[3],这种摄食活动能强烈影响藻类特别是着生藻类的分布、生物量、生产力以及种类组成[4]。Phillips等人研究认为,着生藻类对沉水植物的营养及光竞争是沉水植被在富营养化水体中退化的关键因子[5]。螺类对着生藻类的适度摄食,可以降低藻类对宿主植物的竞争与毒害作用[6]有利于宿主植物的生长,同时也可以加快营养物质的循环利用率,降低藻类之间的光竞争强度,一定程度上促进着生藻类群落的生产力的提高[7]。高等水生植物与浮游藻类存在竞争关系,它们可以通过释放化学物质抑制浮游藻类生长,同时吸收水体中氮、磷等营养物质,从而达到净化水体的效果[8]。因此构建稳定的水生植物-螺类-藻类生态系统对富营养化水体的净化以及藻类的控制有很重要的意义。水生植物可以为螺类提供食物资源(着生藻类)、提供逃避捕食者的避难所、繁殖基质,同时还能提供富氧的底栖环境[9]。螺蛳(Margaryamelanioides)是云南省高原湖泊的特有物种[10],2009年被世界自然保护联盟(IUCN)列入了极危物种(critically endangered,CR)( http://www.iucnredlist. org)。在过去的50a里此物种生物量急剧减少,20个世纪90年代初杜宝汉等对洱海进行了一次全面调查结果显示螺蛳仍为洱海软体动物的优势种[11],而现在洱海流域已经很少能发现其足迹,有预计其可能会在2015年消亡[12]。
笔者通过对洱海流域上游海菜花湿地塘藻类分布特征,水体温度、pH值、以及其他环境要素进行调研,结合螺蛳摄食特性研究,探讨在海菜花水培湿地中对螺蛳育种并进行恢复的可行性。
1 材料与方法 1.1 实验材料实验用螺蛳采自云南省洱海流域上游海菜花湿地螺蛳临时养殖塘,个体健康、大小均匀、螺壳无损,实验前清除螺蛳体表附着生物及污物并饥饿处理48 h,实验前称其湿重。实验用水为曝气处理12h的自来水,pH值为8.5±0.2;实验选取两种硅藻(菱形藻Nitzschia sp.和舟形藻Naviculaceae sp.),一种绿藻(四尾栅藻Scenedesmus quadricauda)和一种蓝藻(孟氏颤藻Planktothrixmougeotii)作为实验用藻。这4种藻均是洱海流域海菜花湿地中存在的典型的附着藻类。
模拟实验是在玻璃缸(长宽高为20cm×20cm×20cm)中进行,放置于AnkeSTX-250-A型号恒温光照培养箱中以保证实验所需条件,玻璃缸中安装曝气速率可调的微孔曝气装置。
1.2 实验方法 1.2.1 实验用藻的培养与定量所用4种藻均通过分离纯化获取。菱形藻和舟形藻采用CSI硅藻培养基扩大培养、四尾栅藻和孟氏颤藻采用BG11培养基扩大培养以实验备用,培养温度为(25±0.5)℃,光照强度为2000lx,24h曝气,光暗比为12h∶12h。
实验中藻的定量均采用血球计数板于显微镜下计数。实验结束时将实验组的螺蛳小心取出并用蒸馏水于玻璃钢中仔细清洗其表面附着藻以及污物,玻璃缸中剩余藻用鲁哥固定液固定浓缩后进行计数。
1.2.2 摄食影响因素实验方法在装有4L水的玻璃缸中放置5个个体均匀的饥饿处理的螺蛳,每个实验组设置1个空白对照以及3个平行,实验结束后将螺蛳放回原暂养塘中。通过控制不同的因素来探讨各因素对螺蛳摄食率的影响。
螺蛳对4种藻的摄食偏好实验开始时装置中4种藻的量见下表 1,实验控制在暗处、25℃和饱和溶解氧条件下进行;于暗处、饱和溶解氧条件下分别调节恒温培养箱温度为10、15、20、25℃和30℃条件探讨温度对螺蛳的摄食影响;在饱和溶解氧和25℃水温条件下调整恒温培养箱光照分别为0、100、500、1000、1500、2000、5000、10000lx探讨光照对螺蛳摄食影响;在暗处、25℃水温条件下将玻璃缸密闭封住通过调整曝气装置的曝气强度来控制适当的溶解氧探讨溶解氧对螺蛳摄食活动的影响;选取5—50g不同质量的螺蛳于25℃、饱和溶解氧条件在暗处培养探讨螺蛳重量与其摄食的关系。
| 藻种名称 Algae | 菱形藻 Nitzschia sp. | 舟形藻 Navicula sp. | 四尾栅藻 Scenedesmus quadricauda | 孟氏颤藻 Planktothrixmougeotii |
| 孟氏颤藻按照长度50μm为1个藻细胞计数 | ||||
| 数量Number/个 | 3.14×108 | 2.94×108 | 3.91×108 | 6.50×108 |
除螺蛳对4种藻摄食偏好实验外其余实验用藻均选用舟形藻Navicula sp.。
除螺蛳重量与摄食关系实验外其余实验均选用重量为25—30g的螺。
1.2.3 螺蛳摄食率计算螺蛳对藻的摄食率根据Frost公式[13]的修正公式计算:
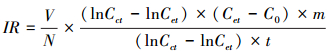
式中,IR摄食率(mg 个-1 d-1);V实验水体体积(mL);N每个实验处理中螺蛳个体数(只);Cct对照组结束时藻细胞浓度(个/mL);Cet实验组结束时藻细胞浓度(个/mL);C0实验起始藻细胞浓度(个/mL);m藻单细胞湿重(mg);t实验持续时间(d)。
1.3 海菜花湿地水体及海菜花叶表面藻类的分布特征调查2013年7月份对洱海北部流域洱源县境内无螺蛳和有螺蛳海菜花湿地塘的藻环境进行调查,各采样点分别采集水样和海菜花植物样,用血球计数板法对水体中及海菜花茎叶表面的藻进行定性定量。
调查的两类海菜花塘均为人工种植塘,水深为1.1m左右,海菜花密度基本相近约为15株/m2,泥质塘底长有少量的轮藻和金鱼藻等沉水植物;含有螺蛳的海菜花塘为螺蛳保种暂养塘(已养殖6个月)螺蛳密度为6 个/m2,除部分低洼地螺蛳分布稍有聚集外,总体分布比较均匀。两类塘中均含有少量的椭圆萝卜螺,密度在0.1个/m2以下,基本无其他大型软体动物分布。
1.4 数据分析本实验数据采用单因素方差分析方法分析差异性,采用双变量相关性分析方法分析相关性。分析作图所用工具为excel2007和SPSS19。
2 结果与分析 2.1 螺蛳对4种典型藻类的摄食偏好螺蛳对4种藻的摄食结果显示舟形藻Navicula sp.的摄食强度显著高于其他3种藻P<0.05,其摄食率为(2.17±0.13)mg 个-1 d-1,对四尾栅藻的摄食强度最低摄食率为(0.82±0.16)mg 个-1 d-1,螺蛳对菱形藻的摄食率为(1.77±1.69)mg 个-1 d-1稍高于孟氏颤藻的(1.72±1.75)mg 个-1 d-1(图 1)。

|
| 图 1 螺蛳对4种藻的摄食率 Fig.1 Ingestion rate of Margarya melanioideson the four algae |
结果显示螺蛳在20℃和25℃时对舟形藻Navicula sp.的摄食量极显著高于其他温度P<0.01(图 2),25℃的摄食率为(2.19±0.16)mg 个-1 d-1,温度小于15℃时螺蛳摄食率均小于0.3mg 个-1 d-1。在光照为0—10000lx范围内螺蛳均有明显的摄食活动;光照强度在1000—2000lx区间内螺蛳摄食活动相对较强均在3.0 mg 个-1 d-1以上,摄食率显著高于其他光照强度,随着光照强度的增加螺蛳的摄食活动呈现先增加后降低的趋势(图 3),螺蛳摄食率在光照强度为1500lx时达到最大为(4.16±0.23)mg 个-1 d-1,说明螺蛳的摄食活动需要一定光照的刺激。DO对螺蛳摄食活动影响显著,在DO小于1mg/L的条件下螺蛳基本停止摄食活动,在DO大于3.6mg/L时摄食活动活跃(图 4),DO为3.6mg/L时螺蛳摄食率达到(3.41±0.17)mg 个-1 d-1。

|
| 图 2 温度对螺蛳摄食的影响 Fig.2 Influences of temperature on Margarya melanioides Ingestion rate of Navicula sp. |

|
| 图 3 光照对螺蛳摄食的影响 Fig.3 Influences of illumination on Margarya melanioides Ingestion rate of Navicula sp. |

|
| 图 4 溶解氧对螺蛳摄食的影响 Fig.4 Influences of DO on MargaryamelanioidesIngestion rate of Navicula sp. |
螺蛳摄食率与其大小有关,体重小的螺蛳摄食率显著高于体重大的(图 5),所选螺蛳中5—10g的螺蛳摄食率最大为(14.18±0.57)mg 个-1 d-1,体重在30—40g的螺蛳摄食率最小均小于0.1 mg 个-1 d-1。

|
| 图 5 螺蛳体重与摄食的关系 Fig.5 The relationship between Margaryamelanioides weight and Ingestion rate of Navicula sp. |
调查结果显示被调查的海菜花塘pH值、DO以及温度基本一致,分别为(8.23±0.23)、(7.69±1.93)mg/L、(20.7±0.3)℃,而两类塘水体中以及海菜花茎叶表面的藻类分布却存在显著差异。
无螺蛳海菜花塘水体中藻类生物量约为8.53×105个/L优势藻均为绿藻和硅藻;有螺塘水体中藻生物量为5.02×105个/L,显著低于无螺塘P<0.05,优势藻为隐藻、甲藻和裸藻(图 6)。

|
| 图 6 海菜花塘水体中藻细胞浓度 Fig.6 Algae concentration of Ottelia acuminate ponds |
茎叶表面藻类分布调查结果知茎叶表面以硅藻为优势藻,无螺塘海菜花茎叶表面硅藻密度为6.67×104个/cm2显著高于有螺塘的5.57×104个/cm2,P<0.05(图 7);硅藻组成中均是以舟形藻为绝对优势藻种,舟形藻占硅藻生物量比例在40%以上。

|
| 图 7 海菜花茎叶表面藻密度 Fig.7 Algae density of Ottelia acuminate leaf surface |
由上述结果可知有螺的海菜花塘水体中及茎叶表面的硅藻密度均显著小于无螺塘,说明螺蛳的存在对水体中硅藻以及海菜花茎叶表面附着硅藻的生物量和分布有明显的影响。
3 讨论本研究的摄食实验证实螺蛳偏好摄食硅藻,螺蛳对舟形藻和菱形藻这两种典型硅藻的摄食率要高于四尾栅藻和颤藻,是因为底栖硅藻含有较绿藻和蓝藻更为丰富的营养物质[14, 15, 16],这也是硅藻在水产养殖业中被广泛用作饵料的原因。
温度是影响动物神经功能和行为能力的直接因素,对水生动物摄食行为也有重要影响。Loosanoff等认为幼贝在低温时由于体内酶不活跃而摄食少生长发育缓慢[17]。螺蛳在20—25℃摄食活动最为强烈,一方面是由于螺蛳在此温度范围内体内酶活性强另一方面可能是由于这个温度段其食物质量较好,硅藻饵料在此温度生长状况最佳,马志珍等[18]报道了底栖舟形藻最适生长温度是25℃,邢荣莲等研究表明在25℃培养时硅藻的蛋白质含量和胞内、外糖含量均最高;20℃培养时硅藻脂肪含量最高[16]。螺蛳在1000—2000lx时摄食率显著高于其他光照区间是由于其触角上有明显的眼点,推测其对光照敏感,一定光照的刺激会激发螺蛳其他功能性神经的应激反应使其摄食活动更加活跃。有研究也表明光照可影响水生动物消化酶能力[19, 20, 21],进而对其摄食效率、生长和代谢等活动有重要影响[22, 23, 24]。一般认为水体中溶解氧低于4mg/L时会引起鱼类窒息死亡,本次对螺蛳摄食研究表明其在DO为3.6mg/L时其摄食活动相当活跃,说明螺蛳适宜耐低溶解氧环境。相关研究也表明在富营养化湖泊水体中大型无脊椎底栖动物对水体的DO 要求较低[25, 26];徐海军、张许峰等认为在水中溶解氧不断降低时,贝类对自身耗氧率进行了调节,使其能更好的维持自身正常的生存和生长[27, 28]。本研究在溶解氧为6mg/L时螺蛳摄食率反而稍低可能是由于曝气装置产生的扰动对其摄食活动稍有影响,此推测有待进一步研究。贝类的个体大小是影响滤水率和摄食率的重要因子之一[29, 30]。Walne等认为外界因子也是决定摄食率的重要因素,牡蛎幼体对几种微型藻的摄食率随水体中藻浓度的增加而增加[31]。本研究表明幼螺摄食率显著高于成螺,说明幼螺对食物的需求更高,在螺蛳繁殖期要适量的补给硅藻,以保证有充足的食物,保障幼仔螺的成活率。
洱海流域海菜花湿地塘中良好的藻类环境可为螺蛳提供良好的食物来源,茁壮的茎叶不仅是舟形藻生长的良好附着基质也是幼螺栖息、躲避与活动的场所;海菜花群落可以改善水下光照和溶氧,并且能人为控制成无竞争者环境,为螺蛳提供安全的赖以生存的环境基础。洱海流域沿湖地区地域广阔、水源充足、气候温和可以通过构建海菜花湿地以达到螺蛳育种,并进一步实现其在洱海流域扩增的目的。
4 结论在所选4种藻中螺蛳喜好摄食舟形藻(Naviculaceae sp.);螺蛳在温度为20—25℃时摄食活动最为活跃;螺蛳生长需要一定光照的刺激,最适宜光照强度范围为1000—2000lx;在DO为3.6mg/L以上时螺蛳的摄食活动均非常强烈,表明其有一定的耐低溶解氧能力;幼螺摄食需求强于成螺。
通过构建海菜花湿地塘对螺蛳进行保种、育种并进一步实现其在洱海流域的扩增恢复是有效可行的。
致谢:感谢中国科学院水生生物研究所徐军老师和谢志才老师在螺蛳摄食以及病害防治方面给予的帮助。
| [1] | Covich A P, Palmer M A, Crowl T A. The role of benthic invertebrate species in freshwater ecosystems:zoobenthic species influence energy flows and nutrient cycling. BioScience, 1999, 49(2):119-127. |
| [2] | Lindegaard C. The role of zoobenthos in energy flow in two shallow lakes. Hydrobiologia, 1994, 275-276(1):313-322. |
| [3] | Chaparro O R, Thompson R J, Pereda S V. Feeding mechanisms in the gastropod Crepidulafecunda. Marine Ecology Progress Series, 2002, 234:171-181. |
| [4] | Stevenson R J, Bothwell M L, Lowe R L, Thorp J H. Algal Ecology:FreshwaterBenthicEcosystem. San Diego, CA:Academic Press, 1996. |
| [5] | Phillips G L, Eminson D, Moss B. A mechanism to account for macrophyte decline in progressively eutrophicated freshwaters. Aquatic Botany, 1978, 4:103-126. |
| [6] | VanVierssen W, Hootsmans M, Vermaat J. Lake Veluwe, a Macrophyte-Dominated System under Eutrophication Stress. Netherlands:Springer, 1994. |
| [7] | Lamberti G A, Gregory S V, Ashkenas L R, Steinman A D, McIntire C D. Productive capacity of periphyton as a determinant of plant-herbivore interactions in streams. Ecology, 1989,70(6):1840-1856. |
| [8] | Ervin G N, Wetzel R G. An ecological perspective of allelochemical interference in land-water interface communities. Plant and Soil, 2003, 256(1):13-28. |
| [9] | Fennessy MS, Cronk J K, Mitsch W J. Macrophyte productivity and community development in created freshwater wetlands under experimental hydrological conditions. Ecological Engineering, 1994, 3(4):469-484. |
| [10] | 陈元晓, 张迺光, 张闻, 李京昆. 阳宗海螺蛳和螺蛳的核型研究. 动物学研究, 1996, 17(1):94-96. |
| [11] | 杜宝汉. 日中洱海生态调查. 海洋与湖沼, 1994, 25(5):532-538. |
| [12] | Song Z Y, Zhang J Q, Jiang X M, Wang C M, Xie Z C. Population structure of an endemic gastropod in Chinese plateau lakes:evidence for population decline. Freshwater Science, 2013, 32(2):450-461. |
| [13] | Frost B W. Effects of size and concentration of food particles on the feeding behavior of the marine planktonic copepod Calanuspacificus. Limnology andOceanography, 1972, 17(6):805-815. |
| [14] | Lebeau T, Robert J M. Diatom cultivation and biotechnologically relevant products. Part II:current and putative products. Applied Microbiology and Biotechnology, 2003, 60(6):624-632. |
| [15] | 邢荣莲. 海洋底栖硅藻的筛选, 培养和应用研究. 大连:大连理工大学, 2007. |
| [16] | 湖北省水生生物研究所藻类研究室藻类应用组. 淡水硅藻的大量培养. 水生生物学集刊, 1975, 5(4):503-512. |
| [17] | Loosanoff V L Davis H C. Rearing of bivalve mollusks. Advances inMarine Biology, 1963, 1:1-136. |
| [18] | 马志珍, 季梅芳, 陈汇远. 一种可作鲍和海参饵料的底栖舟形藻的培养条件的研究. 海洋通报, 1985, 4(4):36-39. |
| [19] | Biswas A K, Seoka M, Inoue Y, Takii K, Kumai H. Photoperiod influences the growth, food intake, feed efficiency and digestibility of red sea bream(Pagrus major). Aquaculture, 2005, 250(3/4):666-673. |
| [20] | Cuvier-Péres A, Jourdan S, Fontaine P, Kestemont P. Effects of light intensity on animal husbandry and digestive enzyme activities in sea bassDicentrachuslabrax post-larvae. Aquaculture, 2001, 202(3/4):317-328. |
| [21] | 王芳, 宋传民, 丁森, 董双林. 光照对中国对虾稚虾3种消化酶活力的影响. 中国水产科学, 2007, 13(6):1028-1032. |
| [22] | Biswas A K, Seoka M, Tanaka Y, Takii K, Kumai H. Effect of photoperiod manipulation on the growth performance and stress response of juvenile red sea bream(Pagrus major). Aquaculture, 2006, 258(1/4):350-356. |
| [23] | García-Esquivel Z, Montes-Magallón S, González-Gómez M A. Effect of temperature and photoperiod on the growth, feed consumption, and biochemical content of juvenile green abalone, Haliotisfulgens, fed on a balanced diet. Aquaculture, 2007, 262(1):129-141. |
| [24] | Taylor J F, North B P, Porter M J R, BromageN R, Migaud H. Photoperiod can be used to enhance growth and improve feeding efficiency in farmed rainbow trout, Oncorhynchusmykiss. Aquaculture, 2006, 256(1/4):216-234. |
| [25] | Gong Z J, Xie P. Impact of eutrophication on biodiversity of the macrozoobenthos community in a Chinese shallow lake. Journal of Freshwater Ecology, 2001, 16(2):171-178. |
| [26] | 王丑明, 谢志才, 宋立荣, 肖邦定, 李根保, 李林. 滇池大型无脊椎动物的群落演变与成因分析. 动物学研究, 2011, 32(2):212-221. |
| [27] | 徐海军, 凌去非, 李倩, 林建华, 沈雯婷, 吴林坤, 曹萍. 六种淡水贝类耗氧率的初步研究. 水产养殖, 2010,31(1):1-4. |
| [28] | 张许峰, 吴小平, 欧阳珊, 甘武. 四种淡水贝的摄食率和耗氧率的实验研究. 海洋湖沼通报, 2008,(3):144-154. |
| [29] | Bayne B L, Newell R C. Physiological energetics of marine molluscs. New York:Academic Press, 1983:407-415. |
| [30] | Winter J E. A review on the knowledge of suspension-feeding in lamellibranchiate bivalves, with special reference to artificial aquaculture systems. Aquaculture, 1978, 13(1):1-33. |
| [31] | Walne P R. Observations on the food value of seven species of Algae to the larvae of Ostreaedulis I. Feeding experiments. Journal of the Marine Biological Association of the United Kingdom, 1963, 43(3):767-784. |