 2016, Vol. 36
2016, Vol. 36文章信息
- 修晨, 郑华, 欧阳志云
- XIU Chen, ZHENG Hua, OUYANG Zhiyun.
- 不同类型人类活动干扰对河岸带外来植物群落的影响——以北京永定河为例
- Community characteristics of riparian alien plants influenced by different types of human disturbance: A case study of Yongding River, Beijing
- 生态学报[J]. 2016, 36(15): 4689-4698
- Acta Ecologica Sinica[J]. 2016, 36(15): 4689-4698
- http://dx.doi.org/10.5846/stxb201501060032
-
文章历史
- 收稿日期: 2015-01-06
- 网络出版日期: 2015-11-16





河岸带介于陆地与河流之间的过渡地带,是指从水-陆交界处至河水影响消失的地带,是水陆间物质、能量和生物的通道,是重要的生态交互作用区[12]。作为一种典型的开放系统,由于频繁的自然和人为干扰,河岸带极易被外来植物所入侵[3-5]。强烈的人类活动如快速城市化(河流过度取水[6]、道路建设[7]、娱乐休憩[8])、农业活动(农田[9]、放牧踩踏[10])等所带来的人为干扰,被普遍认为在河岸带外来植物的引入、传播和产生影响方面起到重要的推动作用[11-13]。人类活动对自然生境的干扰导致河岸带植被的物种构成发生明显变化[14], 外来植物种类的增多与乡土植物种类的减少并存[15-16]。这降低了河岸带原始植被群落的多样性和功能,影响水陆生态系统的物质能量交换等生态系统功能[17-18]。因此,研究不同人为扰动类型下河岸带外来植物群落的特征,对于反映快速城市化背景下河岸带植被结构的变化,了解外来植物在不同人为干扰下进入自然生境的主要途径等,具有十分重要的意义。
尽管国内学者对河岸带植被的分布格局[19]、物种多样性[20]、功能群划分[21]和生态服务功能[22]等方面做了大量的研究工作,但是针对河岸带外来植物的研究较少[23-24],特别是在不同人类活动干扰类型下,外来植物群落的特征,入侵途径和影响程度的相关研究尚未见报道。目前,城市化背景下的河岸带的植被研究及其生态恢复已渐渐成为生态学研究的一个热点,因此分析不同人类活动干扰类型下的河岸带外来植物群落特征,并与原生河段进行对比研究,对于外来植物和河岸带研究都有重要的意义。本研究以海河流域主要支流之一,北京永定河的河岸带外来植物群落为研究对象,在系统的河岸带植被调查的基础上,对不同人类活动干扰类型下的河岸带外来草本植物群落特征进行分析,旨在反映社会经济不断发展、快速城市化等的背景下河岸带外来植物群落的特征。
1 研究地区北京永定河位于38°59′—39°14′N,115°35′—117°5′E,流经北京、河北和天津,全段包括永定河山峡段、城市段和平原段3段,河岸带海拔为10.1—520.8 m之间。该河系全长为210 km,流域面积3168 km2,属于海河流域的一部分,是华北地区最大的河流之一[25]。永定河珠窝水库至三家店段为山峡段,全长70 km,这一段坡降较大,河流较窄,水能资源丰富,受人类活动干扰的影响较弱;三家店至北京南六环路段为城市段,全长约37 km,主要为季节性河道,受快速城市化等较强人类活动干扰的影响;北京南六环路至屈家店为平原段,全长约103 km,受农业活动等人类活动干扰较多(表 1)。由于上游水库拦蓄和地下水水位的下降等原因,该段河道在平水年干涸十分严重,大部分河道已近30年无水。其中,永定河山峡段的年平均流量为808 m3/s。该流域地区气候具有明显的暖温带、半湿润大陆性季风气候特征,年降水量在400—600 mm之间,7月至9月降水量占全年降水量的70%以上[26]。
| 河段River section | 河段长度/kmLength | 海拔/mElevation | 地理位置Location | 不同干扰类型Type of disturbance of human activity |
| 山峡段Mountain section | 70 | 214.4—520.8 | 珠窝水库至三家店 | 原生河段,是人类活动干扰较弱的代表类型河段 |
| 城市段Urban section | 37 | 83.7—99.8 | 三家店至北京南六环路 | 城市干扰类型。主要受快速城市化引起人类活动干扰的影响,具体特征为人口密度大、道路密度高、机动车辆多、土地利用方式剧烈改变,生境破损化,不透水层面积增加等; 是人类活动高干扰的代表类型河段 |
| 平原段Plain secton | 103 | 10.1—64.3 | 北京南六环路至屈家店枢纽 | 农业干扰类型。主要受农田,放牧等农业活动干扰,具体特征是河岸带有农田施肥、人工除草、病虫害防治、放牧踩踏等人为干扰; 属于人类活动干扰较高的代表类型河段 |
自2009年8月至9月,对永定河河岸带外来草本植物群落进行了植被调查,采用分层取样的方法确定样方。首先,在确定研究样地之前,对全河流河岸带做了一次物种普查,明确了不同类型河段的典型外来植物群落。按照每个代表性的植物群落类型,确定一个研究样地。其次,在每个样地设置一条样带:沿着垂直于河道的方向,从河滩处开始每隔5 m设置一个1 m×1 m的草本样方[27],至河岸带边缘(距离河滩处不超过100 m)。通过每个研究样地的相应数量的样方来反映植物群落的基本特征。根据植被类型的代表性原则,本研究在永定河河岸带上共选取了11处样地,共150个样方。其中,山峡段4个样地,共34个样方;城市段3个样地,共55个样方;平原段4个样地,共61个样方(图 1)。

|
| 图 1 北京永定河河岸带研究样地分布示意图 Fig. 1 Locations of the study area, vegetation transects and three river sections of Yongding River in Beijing |
本调查统计每个样方内的草本植物群落特征指标,主要指标为样方内草本植物的种名、生活型、水分生态型、株数、平均高度和地上生物量。对于样方内草本植物的株数的计量,如果草本植物成丛生型或莲座型生长, 则分开计算株数;如果样方内为单一物种且均匀分布, 则调查代表性的25 cm×25 cm面积中的株数再按面积推算样方株数;如果样方内物种分布不均匀,则对1 m×1 m样方进行全部取样。对于地上生物量的取样,原则上将样方内所有草本植物种、全部采样,然后进行烘干、计量;如果样方内有单一性均匀分布的植物种,则采样代表性的25 cm×25 cm面积中的地上生物量再按面积推算样方总生物量。
2.2 数据分析 2.2.1 植物来源的判定方法(1) 乡土植物 乡土植物指在没有人为作用的情况下,自然地分布在特定地区、生态系统或生境的植物种,不包括乡土植物与外来植物的杂交种[28-29]。本文中主要是根据植物种在河流流经地区的自然分布情况及引种、栽培历史来判断[30-31]。
(2) 外来植物 由于人为有意或无意的作用而出现在特定地区的植物种,即为该地区的外来植物[29]。本文中将外来植物分为国内外来中与国外外来种,主要根据原产地,引种记载和在全国各地的主要分布情况来判断[30-31]。
(3) 入侵植物 在特定地区有能力自然生长并能产生可繁殖的下一代外来植物,通常在其自然生长地区之外,距离母株相当远的距离内存在大量个体,并具有大范围扩散的潜力[29]。本文中的入侵植物根据文献[32-33]来确定。
2.2.2 优势度与优势种的确定采用综合优势度指数(synthetic dominance index, SDR4)衡量各物种在河岸带草本植物群落中的优势度[34]。
优势度的计算公式:
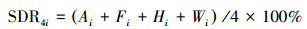
式中, SDR4i为第i种植物的优势度;Ai为第i种植物的相对多度;Fi为第i种植物的相对频度;Hi为第i种植物的相对盖度;Wi为第i种植物的相对重量(地上生物量)。
河岸带草本植物的优势种进行K-means聚类分析得到。在草本植物群落中,采用植物种的优势度(SDR4)作为参数,将各组植物分为两类,方差结构表明两类之间具有显著性的差异(P < 0.05),因此认为其中优势度较高的一类为优势种。
2.2.3 相似性分析采用Sorensen相似性指数[35]来分析不同河段间外来植物群落的物种构成相似性。
其计算公式是:
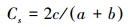
式中, a, b 为河段A和河段B的物种数;c为两个河段的共有植物种。
本文中所用到的数据分析,如方差分析,聚类分析和卡方检验等均使用PASW Statistics 18软件进行分析处理。
3 结果与分析 3.1 河岸带外来植物的物种构成及来源本调查中,永定河河岸带植被共有维管束植物27科72属101种,外来植物13科28属29种,外来草本植物占所有植物的28.7%。其中源于国内的外来种12种,隶属于8科12属;源于国外的外来种17种,隶属于8科16属;外来入侵植物11种。国外外来种主要来自美洲和亚洲,分别占64.71%和17.65%。在外来植物科的组成结构上,植物种类主要集中于少数几个大科如菊科(Compositae)、禾本科(Gramineae)、豆科(Leguminosae)和苋科(Amaranthaceae)。其中,菊科是国外外来种最主要的科,包括6个种,其次是苋科,包含3个种。国内外来种最主要的科是禾本科,包含有4个种。外来种比较多的属较少,其中国外外来种的种数以苋属(Amaranthus)最多,为2个种。
3.2 外来种比例不同河段河岸带草本植物群落的物种构成呈现出显著性差异(F=40.59, df=4, P=0.001)(图 2)。相比于原生河段,随着人类活动干扰逐渐加强,外来种比例由山峡段的20.9%,上升至平原段和城市段的30.2%和25.5%。国外外来种在山峡段的比例最低,仅为7.0%,在平原段和城市段则上升至18.6%和15.7%(图 2)。其中,典型的国外外来草本植物种有反枝苋(Amaranthus retroflexus)、牛筋草(Eleusine indica)和大麻(Cannabis sativa)等。国内外来种在山峡段较高,为14.0%,在平原段和城市段则将至11.6%和9.8%(图 2)。典型的国内外来种有假稻(Leersia japonica)和芒(Miscanthus sinensis)等。

|
| 图 2 北京永定河不同河段河岸带外来植物种比例和外来植物优势度 Fig. 2 Percentage of Alien plants and synthetic dominance index (SDR4) in the riparian zone of different river sections in Yongding River |
随着人类活动干扰强度不断增加,河岸带植物物种的优势度(SDR4)构成在不同河段也显现出显著性差异(F=18.69, df=4, P=0.001)(图 2)。外来草本植物的优势度呈逐渐上升趋势,优势度比例由山峡段的12.1%,上升至平原段和城市段的13.4%和17.5%。国外外来种优势度在山峡段最低,仅为3.3%,在平原段和城市段则增加至7.1%和13.4%,呈现逐渐上升的趋势。国内外来种优势度则成相反的趋势,在山峡段最高,为8.7%,而在平原段和城市段则降低为6.3%和3.4%(图 2)。
沿着人类活动干扰的环境梯度,不同河段的外来植物优势种有所不同。其中,山峡段的优势种为芒和假稻,均为国内外来种。平原段的优势种有4个,分别是芒、假稻、大麻和磨盘草(Abutilon indicum),其中大麻为外来入侵种,其它3种均为国内外来种。城市段的优势种为反枝苋和牛筋草,均为外来入侵种(图 2,表 3),其优势度已超过该河段81.6%的乡土种。
| 种名Species | 科名Family | 物种总株数Total species abundance | 物种来源Origin of species | 入侵种Invasive species | ||
| 山峡段Mountain section | 平原段Plain section | 城市段Urban section | ||||
| 假稻Leersia japonica | 禾本科 | 528 | 370 | 4 | 国内外来 | |
| 牛筋草Eleusine indica | 禾本科 | 0 | 6 | 949 | 国外外来 | √ |
| 圆叶牵牛Pharbitis purpurea | 旋花科 | 0 | 1 | 3 | 国外外来 | √ |
| 荻Triarrhena sacchariflora | 禾本科 | 0 | 1 | 16 | 国内外来 | |
| 芒Miscanthus sinensis | 禾本科 | 200 | 29 | 0 | 国内外来 | |
| 三叶鬼针草Bidens pilosa | 菊科 | 7 | 0 | 0 | 国外外来 | √ |
| 小蓬草Conyza canadensis | 菊科 | 1 | 0 | 0 | 国外外来 | |
| 土三七Gynura segetum | 景天科 | 180 | 0 | 0 | 国内外来 | |
| 菊芋Helianthus tuberosus | 菊科 | 2 | 0 | 0 | 国外外来 | √ |
| 黄香草木犀Melilotus officinalis | 豆科 | 2 | 0 | 0 | 国内外来 | |
| 唐松草Thalictrum aquilegifolium | 毛茛科 | 46 | 0 | 0 | 国内外来 | |
| 菅Themeda villosa | 禾本科 | 8 | 0 | 0 | 国内外来 | |
| 磨盘草Abutilon indicum | 锦葵科 | 0 | 2 | 0 | 国内外来 | |
| 繁穗苋Amaranthus paniculatus | 苋科 | 0 | 4 | 0 | 国外外来 | |
| 钻叶紫菀Aster subulatus | 菊科 | 0 | 172 | 0 | 国外外来 | |
| 大麻Cannabis sativa | 桑科 | 0 | 3 | 0 | 国外外来 | √ |
| 畦畔莎草Cyperus haspan | 莎草科 | 0 | 7 | 0 | 国内外来 | |
| 番薯Ipomoea batatas | 旋花科 | 0 | 25 | 0 | 国外外来 | |
| 紫苜蓿Medicago sativa | 豆科 | 0 | 2 | 0 | 国外外来 | √ |
| 芝麻Sesamum indicum | 胡麻科 | 0 | 2 | 0 | 国外外来 | |
| 喜旱莲子草Alternanthera philoxeroideS | 苋科 | 0 | 0 | 102 | 国外外来 | √ |
| 反枝苋Amaranthus retroflexus | 苋科 | 0 | 0 | 388 | 国外外来 | √ |
| 豚草Ambrosia artemisiifolia | 菊科 | 0 | 0 | 33 | 国外外来 | √ |
| 虎尾草Chloris virgata | 禾本科 | 0 | 0 | 79 | 国外外来 | |
| 曼陀罗Datura stramonium | 茄科 | 0 | 0 | 2 | 国外外来 | √ |
| 苦荬菜Ixeris polycephala | 菊科 | 0 | 0 | 2 | 国内外来 | |
| 鸡眼草Kummerowia striata | 豆科 | 0 | 0 | 3 | 国内外来 | |
| 砂引草Messerschmidia sibirica | 紫草科 | 0 | 0 | 2 | 国内外来 | |
| 意大利苍耳Xanthium italicum | 菊科 | 0 | 0 | 6 | 国外外来 | √ |
外来植物群落的生活型结构在不同河段具有显著性差异(F=120.9, df=10, P=0.001)(图 3)。山峡段外来植物种比例最高的生活型是多年生草本植物,为66.7%;其中又以多年生中生草本的比例最高,达到55.6%;多年生湿生草本的比例较低,为11.1%。一年生植物也呈现出较高的比例,均属于中生草本植物,达到了22.2%。平原段外来植物种的生活型结构由一年生和多年生草本组成,其中一年生中生草本比例最高,达到了53.8%;多年生中生草本和多年生湿生草本的比例位居其次,分别达到了30.8%和15.4%。城市段外来植物种比例最高的生活型是一年生草本植物,到达了69.2%;其中,一年生中生草本和一年生湿生草本比例分别为53.8%和15.4%。多年生植物包括多年生湿生草本、中生草本和旱生草本,比例分别为15.4%、7.7%和7.7%(图 3)。重要的多年生中生草本有菊芋和芒等,多年生湿生草本有假稻等。而重要的一年生中生草本有牛筋草和大麻等,一年生湿生草本有意大利苍耳(Xanthium italicum)等。

|
| 图 3 北京永定河不同河段河岸带外来植物的生活型结构 Fig. 3 Life span of Alien species in the riparian zone of different river sections in Yongding River in Beijing |
采用Sorensen相似性指数,对不同河段的外来植物物种构成进行相似性分析。结果显示,城市段与平原段相似性最高,为0.308;山峡段与平原段其次,为0.182;山峡段与城市段相似性最低,为0.091(表 2)。城市段与平原段共有的植物种有4种,分别是假稻、牛筋草、圆叶牵牛(Pharbitis purpurea)和荻(Triarrhena sacchariflora),其中牛筋草和圆叶牵牛都是入侵种;山峡段与平原段共有的物种有2种,分别为假稻和芒;相似性最低的山峡段与城市段之间,仅有假稻一个共有种。而假稻是唯一一种在3个河段都出现的植物(表 2, 表 3)。
| 河段类型River section | Sorensen相似性指数Sorensen Classic index | |
| 山峡段Mountain section | 城市段Urban section | 0.091 |
| 山峡段Mountain section | 平原段Plain section | 0.182 |
| 城市段Urban section | 平原段Plain section | 0.308 |
虽然不同河段所独有的外来植物种数量比较相近,但是随着人类活动干扰强度不断增加,物种的更替变化导致其构成呈现出很大的差异。山峡段、平原段和城市段出现的独有外来植物分别有8种、8种和9种(表 3)。但是,从山峡段到平原段,新出现的国外外来种数量与山峡段相比增长了1倍,由原来的37.5%增加至75%,新出现的物种主要有繁穗苋(Amaranthus paniculatus)、番薯(Ipomoea batatas)和钻叶紫菀(Aster subulatus)等。从平原段到城市段,新出现的国外外来种数量上虽然与平原段持平,但其中却包含5个外来入侵种,分别是豚草(Ambrosia artemisiifolia)、曼陀罗(Datura stramonium)、意大利苍耳、喜旱莲子草(Alternanthera philoxeroideS)和圆叶牵牛(表 3)。
3.6 外来入侵种随着人类活动的干扰强度不断增加,外来入侵种比例逐渐增大,从山峡段的4.7%,增加至平原段和城市段的9.3%和13.7%(图 4)。同时,外来入侵种的优势度也逐渐增加。与山峡段、平原段相比,城市段外来入侵种的优势度也由2.2%、3.7%上升至14.5%(图 4)。本次北京永定河河岸带植物调查共记录到赵娟娟[36]、刘全儒[33]、李振宇和解焱[32]及王苏铭[37]等记载的河岸带草本外来入侵植物11种(表 3)。其中,入侵植物牛筋草在本次调查中同时出现在平原段和城市段,出现频度达15次,居所有外来草本植物的第1位,优势度居第2位(外来植物共有29种),分布较为广泛。反枝苋则是城市段中最优势的入侵植物,在所有草本植物(共有51种,包括乡土和外来种)中优势度居第4位,优势度高于城市段92%的乡土植物种。

|
| 图 4 北京永定河不同河段河岸带的外来入侵种物种比例和外来入侵种优势度 Fig. 4 Percentage of invasive species and synthetic dominance index (SDR4) of invasive species in the riparian zone of different river sections in Yongding River |
北京永定河河岸带外来草本植物共有13科28属29种,外来植物种数所占比例为28.7%,表现出较高的比例,这与国内外其他河流河岸带外来植物的研究结果(24%—30%)相比,比例相似[4, 24]。在外来植物科的组成结构上,北京永定河外来植物种主要以菊科、禾本科、豆科和苋科等被子植物中的几个大科为主。这些科作为维管束植物中占据主导地位的植物科,其本身的特点有利于成功进入新的环境中[38-39],而河岸带临近水道、环境开阔,这也对风媒、水媒传播的外来植物的传播提供了便利。如,菊科自身具备有利于扩散的特征,如繁殖率高、繁殖方式多样、风媒传播等;苋科植物则具有种子小、数量多的特点,且具有较广泛的药用价值,易被人为引入;而豆科植物具备固氮能力,通过自身营养累积而获得资源上的优势[40]。
本研究中,不同河段间的河岸带外来草本植物群落的结构有显著的差异,外来种比例、优势度和生活型结构变化十分明显。总体上,外来植物群落呈现出:外来种比例和优势度在山峡段较低,在平原段和城市段较高。多年生草本植物在山峡段比例最高,平原段其次,城市段最低;一年生草本植物在城市段最高,平原段其次,山峡段最低。形成这种格局的可能原因是:山峡段处于北京永定河上游山区,受地形自然因素,如河道狭窄且密布鹅卵石、河岸土壤稀薄养分有限等;此外,受交通、经济等人类活动干扰较小,因此外来种在较少。外来物种构成上也以国内外来种为主,如芒和假稻等。生活型结构是群落特征中用于刻画植物群落对人类干扰响应的有效工具[41]。一年生植物由于其生长速度快,繁殖时间早,种子数量多等特征,因此在受干扰强烈的环境中有较多的分布[42]。城市段和平原段的外来植物群落的生活型结构中,一年生植物的比例都较高,这反映了两个河段的河岸带环境都受到了一定程度的人类活动干扰。但是不同河段受干扰的类型是有差异的。
平原段是农业干扰类型,人类影响主要来自农耕、放牧及村镇生活垃圾等。平原段处于永定河下游,多为广阔的河漫滩,因此出现了部分河岸带被周边农田、菜地等侵蚀,以及家畜放牧等现象。农业引种等人为引入是该河段外来植物进入河岸带的途径之一。在该河段河岸带出现了频度较高的农业外来植物,如番薯是当地引种较多的农田作物,而繁穗苋为一种优良的牧草植物;同时也出现了优势度较高的农业类杂草,如禾本科的假稻适生于湿地和田边,是典型的危害农田的农业杂草。此外,交通道路也是外来物种传播的重要途径,频繁的交通运输增加了植物种子或者其他类型的繁殖体扩散的机会[43-44]。本研究中,由于河岸带一侧大多建有沿河公路,这利于一些外来植物种子随人流和车辆流动而迁入。如菊科的钻叶紫菀,其种子小数量多,易沿路边河道蔓延,快速传播并侵入河岸带生态系统[23]。农业干扰类型下,外来植物主要通过人为引种和交通道路等途径进入河岸带。
城市段是城市干扰类型,主要受快速城市化等剧烈人类活动干扰的影响。在人类的各种活动中,城市化对外来植物的进入有显著的促进作用[45]。城市化过程中,人类活动(建筑工程,水利设施建设等)引起的土地利用类型改变,破坏了原有植被,这可能增加外来种迁入和定居的机会[46-47],促进了外来种的扩散和传播[48],是导致外来植物侵入的重要途径之一。在本研究中,与山峡段相比,城市段的外来植物的物种比例增加、优势度增大;而外来入侵种的比例和优势度更是达到最高,分别为13.7%和14.5%,其中,外来入侵种反枝苋和牛筋草的优势程度已经超越绝大部分乡土植物。这反映了在城市化所导致的土地利用类型改变的影响下,外来植物种呈现扩大趋势,入侵程度加深。这与国外同类研究结果一致,Meek等人发现河岸带植物群落外来植物入侵情况与河岸带生态系统周边的土地利用类型具有显著的关系,与城市接壤的河岸区域具有最高的外来植物丰富度[16]。此外,北京城市环境中巨大的人流物流、贸易往来等也增加了无意携带外来植物种子的风险,为外来物种的扩散创造了便利条件。
虽然目前外来植物的比例尚在可控范围内,但是在人类活动干扰的影响下,北京永定河河岸带依然存在较高的生物入侵风险。城市化建设所导致的生态扰动,河流断流都可能为生物入侵创造有利条件[49],在将来的防治工作中应采取更加严格的检疫制度,防止已知的入侵物种传入北京。同时,在对退化的河岸带进行生态修复的时候,应以乡土湿地植物的推广种植为主,这对于防范外来植物入侵同样具有重要意义。乡土物种在适应当地小环境方面更具优势,对本地的气候、土壤环境、水位的变化有较强的适应性;乡土植物一般多年生长,可建立稳定的植物群落,以较小的生态代价来恢复植被覆盖。此外,应加强对尚未入侵的河岸带的保护,防止新的外来植物再次入侵。
致谢: 首都师范大学胡东副教授对植物分类和植物群落调查给予帮助,张志明、赵娟娟、王芸、白杨、孟世勇、刘成立、孙成耀对文章讨论、植物群落调查和取样给予帮助,特此致谢。| [1] | 陈吉泉. 河岸植被特征及其在生态系统和景观中的作用[J]. 应用生态学报 , 1996, 7 (4) : 439–448. |
| [2] | 郭怀成, 黄凯, 刘永, 郁亚娟. 河岸带生态系统管理研究概念框架及其关键问题[J]. 地理研究 , 2007, 26 (4) : 789–798. |
| [3] | David M Richardson, Patricia M Holmes, Karen J Esler, Susan M Galatowitsch, Juliet C Stromberg, Steven P Kirkman, Petr Pyšek, Richard J Hobbs. Riparian vegetation: degradation, alien plant invasions, and restoration prospects[J]. Diversity and Distributions , 2007, 13 (1) : 126–139. DOI:10.1111/j.1366-9516.2006.00314.x |
| [4] | Anne-Marie Planty-Tabacchi, Eric Tabacchi, Robert J Naiman, Collette Deferrari, Henri Décamps. Invasibility of species-rich communities in riparian zones[J]. Conservation Biology , 1996, 10 (2) : 598–607. DOI:10.1046/j.1523-1739.1996.10020598.x |
| [5] | Kate Rowntree. An assessment of the potential impact of alien invasive vegetation on the geomorphology of river channels in South Africa[J]. Southern African Journal of Aquatic Sciences , 1991, 17 (1/2) : 28–43. |
| [6] | An S Q, Chen X L, Sun C S, Wang Y J, Li J. Composition change and vegetation degradation of riparian forests in the Altai Plain, NW China[J]. Plant Ecology , 2003, 164 (1) : 75–84. DOI:10.1023/A:1021225204808 |
| [7] | Derric N Pennington, James R Hansel, David L Gorchov. Urbanization and riparian forest woody communities: diversity, composition, and structure within a metropolitan landscape[J]. Biological Conservation , 2010, 143 (1) : 182–194. DOI:10.1016/j.biocon.2009.10.002 |
| [8] | Washitani I. Plant conservation ecology for management and restoration of riparian habitats of lowland Japan[J]. Population Ecology , 2001, 43 (3) : 189–195. DOI:10.1007/s10144-001-8182-8 |
| [9] | Kirstin Deutschewitz, Angela Lausch, Ingolf Kühn, Stefan Klotz. Native and alien plant species richness in relation to spatial heterogeneity on a regional scale in Germany[J]. Global Ecology and Biogeography , 2003, 12 (4) : 299–311. DOI:10.1046/j.1466-822X.2003.00025.x |
| [10] | N Meeson, Robertson A I, Jansen A. The effects of flooding and livestock on post-dispersal seed predation in river red gum habitats[J]. Journal of Applied Ecology , 2002, 39 (2) : 247–258. DOI:10.1046/j.1365-2664.2002.00706.x |
| [11] | Annett Wania, Ingolf Kühn, Stefan Klotz. Plant richness patterns in agricultural and urban landscapes in Central Germany-spatial gradients of species richness[J]. Landscape and Urban Planning , 2006, 75 (1/2) : 97–110. |
| [12] | Roy D B, Hill M O, Rothery P. Effects of urban land cover on the local species pool in Britain[J]. Ecography , 1999, 22 (5) : 507–517. DOI:10.1111/eco.1999.22.issue-5 |
| [13] | Karen P Alston, David M Richardson. The roles of habitat features, disturbance, and distance from putative source populations in structuring alien plant invasions at the urban/wildland interface on the Cape Peninsula, South Africa[J]. Biological Conservation , 2006, 132 (2) : 183–198. DOI:10.1016/j.biocon.2006.03.023 |
| [14] | Jane A Catford, Barbara J Downes. Using multi-scale species distribution data to infer drivers of biological invasion in riparian wetlands[J]. Diversity and Distributions , 2010, 16 (1) : 20–32. DOI:10.1111/j.1472-4642.2009.00631.x |
| [15] | Maskell L C, Firbank L G, Thompson K, Bullock J M, Smart S M. Interactions between non-native plant species and the floristic composition of common habitats[J]. Journal of Ecology , 2006, 94 (6) : 1052–1060. DOI:10.1111/jec.2006.94.issue-6 |
| [16] | Clifton S Meek, David M Richardson, Ladislav Mucina. A river runs through it: land-use and the composition of vegetation along a riparian corridor in the Cape Floristic Region, South Africa[J]. Biological Conservation , 2010, 143 (1) : 156–164. DOI:10.1016/j.biocon.2009.09.021 |
| [17] | Heather Greenwood, Dennis J O'Dowd, P S Lake. Willow (Salix x rubens) invasion of the riparian zone in south-eastern Australia: reduced abundance and altered composition of terrestrial arthropods[J]. Diversity and Distributions , 2004, 10 (5/6) : 485–492. |
| [18] | Philip E Hulme, Eleanor T Bremner. Assessing the impact of Impatiens glandulifera on riparian habitats: partitioning diversity components following species removal[J]. Journal of Applied Ecology , 2006, 43 (1) : 43–50. |
| [19] | 郭跃东, 郭晋平, 张芸香, 吉久昌. 文峪河上游河岸林群落环境梯度格局和演替过程[J]. 生态学报 , 2010, 30 (15) : 4046–4055. |
| [20] | 江明喜, 邓红兵, 唐涛, 蔡庆华. 香溪河流域河岸带植物群落物种丰富度格局[J]. 生态学报 , 2002, 22 (5) : 629–635. |
| [21] | 郭屹立, 卢训令, 丁圣彦. 伊洛河河岸带生态系统草本植物功能群划分[J]. 生态学报 , 2012, 32 (14) : 4434–4442. |
| [22] | 郭二辉, 孙然好, 陈利顶. 河岸植被缓冲带主要生态服务功能研究的现状与展望[J]. 生态学杂志 , 2011, 30 (8) : 1830–1837. |
| [23] | 郭屹立, 丁圣彦, 苏思, 卢训令, 苑晓雯, 郭静静, 李乾玺. 伊洛河流域外来草本植物分布格局[J]. 生态学报 , 2013, 33 (17) : 5438–5447. |
| [24] | 左倬, 蒋跃, 薄芳芳, 沈蓓蕾, 由文辉. 平原河网地区滨岸带外来植物入侵现状及影响研究——以上海青浦区为例[J]. 生态环境学报 , 2010, 19 (3) : 665–671. |
| [25] | Wang L, Wang Z J, Toshio Koike, Yin H, Yang D W, He S. The assessment of surface water resources for the semi-arid Yongding River Basin from 1956 to 2000 and the impact of land use change[J]. Hydrological Processes , 2010, 24 (9) : 1123–1132. DOI:10.1002/hyp.v24:9 |
| [26] | 张宗祜. 华北平原地下水环境演化. 北京: 地质出版社, 2000. |
| [27] | Wang G H, Li H, An M, Ni J, Ji S J, Wang J. A regional-scale consideration of the effects of species richness on above-ground biomass in temperate natural grasslands of China[J]. Journal of Vegetation Science , 2011, 22 (3) : 414–424. DOI:10.1111/jvs.2011.22.issue-3 |
| [28] | USNA. What is a native plant, anyway? 2006-03-07 [2012-10-10]. http://tiny.cc/il8imw. |
| [29] | Petr Pysek. Alien and native species in Central European urban floras: a quantitative comparison[J]. Journal of Biogeography , 1998, 25 (1) : 155–163. DOI:10.1046/j.1365-2699.1998.251177.x |
| [30] | 贺士元, 邢其华, 尹组棠, 江先甫. 北京植物志(第三版). 北京: 北京出版社, 1993. |
| [31] | 中国科学院植物研究所.中国数字植物标本馆(CVH). [2012-09-20]. http://www.cvh.org.cn. |
| [32] | 李振宇, 解焱. 中国外来入侵种. 北京: 中国林业出版社, 2002. |
| [33] | 刘全儒, 于明, 周云龙. 北京地区外来入侵植物的初步研究[J]. 北京师范大学学报:自然科学版 , 2002, 38 (3) : 399–404. |
| [34] | Wang Z W, Xing F, Zhu T C, Li X C. The responses of functional group composition and species diversity of Aneurolepidium chinense grassland to flooding disturbance on Songnen Plain, Northeasten China[J]. Acta Phytoecologica Sinica , 2002, 26 (6) : 708–716. |
| [35] | Anne E Magurran. Ecological Diversity and Its Measurement. Princeton, New Jersey: Princeton University Press, 1988. |
| [36] | 赵娟娟.北京市建成区城市植物的种类构成与分布格局[D].北京:中国科学院研究生院, 2010. |
| [37] | 王苏铭, 张楠, 于琳倩, 赵容慧, 郝鹏, 李景文, 姜英淑, 沙海峰, 刘义, 张志翔. 北京地区外来入侵植物分布特征及其影响因素[J]. 生态学报 , 2012, 32 (15) : 4618–4629. |
| [38] | Petr Pysek, David M Richardson, Jan Pergl, Vojtěch Jarosik, Zuzana Sixtova, Ewald Weber. Geographical and taxonomic biases in invasion ecology[J]. Trends in Ecology & Evolution , 2008, 23 (5) : 237–244. |
| [39] | V Heywood. Patterns, extents and modes of invasions by terrestrial plants//J A Drake, H A Mooney, F di Castri, R H Grovers, F J Kruger, M Rejmánek, M Williamson, eds. Biological Invasions: A Global Perspective. Chichester: Wiley, 1989: 31-60. |
| [40] | 吴征镒. 中国植被. 北京: 科学出版社, 19801–1144. |
| [41] | McIntyre S, Lavorel S, Tremont R M. Plant life-history attributes: their relationship to disturbance response in herbaceous vegetation[J]. The Journal of Ecology , 1995, 83 (1) : 31–44. DOI:10.2307/2261148 |
| [42] | J Philip Grime. Plant Strategies and Vegetation Processes. New York: Wiley, 1979. |
| [43] | Stephen C Trombulak, Christopher A Frissell. Review of ecological effects of roads on terrestrial and aquatic communities[J]. Conservation Biology , 2000, 14 (1) : 18–30. DOI:10.1046/j.1523-1739.2000.99084.x |
| [44] | Marion Hourdequin M. Introduction[J]. Conservation Biology , 2000, 14 (1) : 16–17. DOI:10.1046/j.1523-1739.2000.99358.x |
| [45] | 彭羽, 刘雪华. 城市化对植物多样性影响的研究进展[J]. 生物多样性 , 2007, 15 (5) : 558–562. |
| [46] | Melinda D Smith, Alan K Knapp. Exotic plant species in a C4-dominated grassland: invasibility, disturbance, and community structure[J]. Oecologia , 1999, 120 (4) : 605–612. DOI:10.1007/s004420050896 |
| [47] | W M Lonsdale, A M Lane. Tourist vehicles as vectors of weed seeds in Kakadu National Park, Northern Australia[J]. Biological Conservation , 1994, 69 (3) : 277–283. DOI:10.1016/0006-3207(94)90427-8 |
| [48] | Jessica J Hellmann, James E Byers, Britta G Bierwagen, Jeffrey S Dukes. Five potential consequences of climate change for invasive species[J]. Conservation Biology , 2008, 22 (3) : 534–543. DOI:10.1111/j.1523-1739.2008.00951.x |
| [49] | 徐汝梅, 叶万辉. 生物入侵:理论与实践. 北京: 科学出版社, 2003. |