 2016, Vol. 36
2016, Vol. 36文章信息
- 王猛, 汪季, 蒙仲举, 柴享贤, 吕世杰, 王德慧, 乌云嘎
- WANG Meng, WANG Ji, MENG Zhongju, CHAI Xiangxian, LÜ Shijie, WANG Dehui, WU Yunga.
- 巴丹吉林沙漠东缘天然梭梭种群空间分布异质性
- Spatial heterogeneity of natural Haloxylon ammodendron Populations at Ta-Mu-Su, Badain Jaran Desert, China
- 生态学报[J]. 2016, 36(13): 4055-4063
- Acta Ecologica Sinica[J]. 2016, 36(13): 4055-4063
- http://dx.doi.org/10.5846/stxb201411142250
-
文章历史
- 收稿日期: 2014-11-14
- 网络出版日期: 2015-10-29





2. 内蒙古农业大学理学院, 呼和浩特 010018;
3. 内蒙古农业大学农学院, 呼和浩特 010018
2. College of Science, Inner Mongolia Agricultural University, Hohhot 010018, China;
3. College of Agricultural, Inner Mongolia Agricultural University, Hohhot 010018, China
种群空间分布格局是植物种群生物学特性对环境条件长期适应和选择的结果[1],它与物种的生物学特性、种间竞争以及生境条件等密切相关[2],是衡量种群昌盛与否的重要指标,关系到种群的生存和进化[3]。空间异质性(Spatial heterogeneity)是植被格局形成的主要原因[4-6],生态系统所展现的异质性和格局是种群动态、群落组织和元素循环的基础[6]。在大多情况下,对空间异质性的理解多局限于定性水平,而定量描述单纯基于数据类型[7]。半方差函数是以区域化变量理论为基础,分析自然现象空间变异和空间相关的统计学方法,是描述空间异质性的有效方法[8-9]。巴丹吉林沙漠发育于温带干旱区,有着独特的植被盖度特征,空间异质性在荒漠的各个尺度上都存在,异质性也是区域荒漠化的重要指标之一[8-10]。
梭梭(Haloxylon ammodendron (C. A. Mey.) Bunge)是藜科(Chenopodiaceae)梭梭属(Haloxylon Bunge)灌木,是温带荒漠分布最为广泛的一种荒漠植被,在遏制土地沙化、改良土壤、恢复植被、维护荒漠生态平衡起着无可比拟的作用,被称为“沙漠植被之王”[11-15]。有关梭梭属植物和以梭梭属植物为优势的荒漠植被的研究倍受关注,主要集中在梭梭种子萌发特性、抗旱生理、生态以及物候期观察等方面[3, 11-24],并取得了大量研究成果。梭梭是荒漠建群种,生态适应幅度较宽,在不同的生境条件下,其种群特征存在差异[3, 14-16, 19-24]。王继和[21]得出随着植被类型由荒漠草原向典型荒漠过渡,梭梭群落相对数量及相对盖度增大,由伴生种转化为建群种;王春玲[22]在准噶尔盆地东南缘发现,梭梭群落天然更新幼苗幼树的分布格局均为聚集分布,以半流动沙丘上的梭梭群落为最多;张锦春[23]通过研究民勤沙区天然梭梭林种群特征,得出由于种群种内或种间竟争和所处的生境不同,沙地梭梭种群呈集群分布格局,盐碱地梭梭种群则为均匀分布;常静[24]研究表明:梭梭种群和白梭梭种群在0—50 m空间尺度范围内均呈现聚集分布,梭梭-白梭梭群落优势种种群以缀块镶嵌形式分布。在不同发育阶段,梭梭的分布格局不同,幼苗和幼树呈聚集分布,成年树呈随机分布[16];随径级的增大,梭梭种群分布由聚集分布向随机分布或均匀分布发展[3, 19]。
位于巴丹吉林沙漠东缘的塔木素,其境内现存珍贵天然梭梭林总面积21.5万hm2,占阿拉善右旗天然梭梭林的92%,为当地“肉苁蓉及梭梭产籽基地”,是研究荒漠生态系统的重要基地。掌握现存梭梭群落的空间分布规律及其与环境之间的互作过程,对更好地保护当地梭梭种群和物种多样性意义重大。本文通过株高、冠幅直径、分布密度等关键指标,分析梭梭群落数量特征空间异质性及与其环境异质之间的关联,旨在阐明极端严酷生境条件下天然梭梭的分布、生长现状,对梭梭种群的演替规律和生态系统稳定性维持机制进行探讨,为当地生态保护和经济发展提供技术支撑。
1 研究地区与方法 1.1 试验地概况试验地位于40°33.059′—40°35.531′N,103°25.162′—103°27.546′E,行政区划隶属于内蒙古阿拉善盟阿拉善右旗塔木素苏木格日勒图嘎查的朝恒扎干(图 1,图 2)。研究区海拔高度范围1255.8—1264.3 m;属典型温带干旱荒漠性气候,年降水量40—80 mm,多集中于5—9月,年潜在蒸发量达到年降水量40倍以上,干燥度7—12;年均温度8—8.9 ℃,绝对最高温度37—43 ℃,平均无霜期150d;风沙频繁,年均风速 4m/s;研究区流沙带以新月型沙丘和简单横向沙丘为主,沙丘主要走向为SW-NE。自然植被稀少,种属贫乏、覆盖率低,地表裸露。

|
| 图 1 产籽基地范围及大样方位置 Fig. 1 Seed production base scope |

|
| 图 2 实验样点分布 Fig. 2 The experimental sample |
产籽基地呈不规则五边形,面积923 hm2,根据实际情况,以梭梭分布较为集中的区域作为试验大样地(图 1),分别在大样地4个端点处,采用手持GPS(彩途N300) 定点记录,以图 3左下角起始作为绝对坐标原点(0,0) ,横向为X轴,纵向为Y轴,然后将其纵横等距离每隔200m设一条样线,形成48个200 m×200 m的相邻网格(面积192 hm2),大样地面积占“产籽基地”总面积的20.8%。

|
| 图 3 梭梭分布图 Fig. 3 Haloxylon ammodendron distribution |
选择梭梭生长旺季,采用手持GPS沿样线记录出现的每一株梭梭基部紧贴地面处的坐标,同时采用5m量程卷尺测量株高(cm)、冠幅直径(cm),对于株高大于150cm的梭梭,架设便携式梯子(XB-3001) 测量。每株梭梭测量5次,取其均值用于结果分析。为使梭梭测定结果与样点(GPS定点,测定经纬度及海拔高度)结合分析,采用Surfer 11.0生成图 3(图中每一“+”均代表一株梭梭),最终形成密度(株/200 m)、株高(cm,每200 m样线内全部梭梭的均值)和冠幅直径(cm,每200 m样线内全部梭梭的均值)三维空间数据点。
1.3 数据分析(1) K-S正态性检验[25]
Kolmogorov-Smirnov(基于经验分布函数(Experience Distributed Function,EDF)的检验)的D = max︱Fn(x)-F0(x)︱统计量检验正态性;检验时,根据样本计算一个统计量即检验统计量D。样本分布的形状和正态分布相比较得出一个数值P,如果P值小于0.05(给定的显著性水平),认为数据不是来自正态分布,反之则认为数据来自正态分布。
首先将梭梭测量点数字化,根据相应梭梭种群数量特征、海拔高度的数据资料,运用Surfer 11.0生成用于地统计学分析的样点分布图,再运用传统统计方法和地统计学方法和原理,对数据进行处理,判断其空间变异性。变异系数(C.V)表示随机变量的离散程度,C.V≤ 10%为弱变异性;10% <C.V <30%为中等变异性;C.V≥ 30%为强变异性。偏度是刻画数据的对称性指标,偏度系数均为正值,表示数据的右侧更分散。峰度系数为正值,表示两侧的极端数据较多,峰度系数为负值,表示两侧的极端数据较少。
半方差函数采用下式计算:
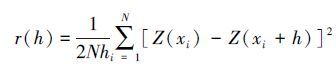
|
式中,r(h)为半方差函数;h为样点空间间隔距离;N(h)为间隔距离为h的样点数;Z(xi)和Z(xi+h)分别为区域化变量Z(x)在空间位置xi和xi + h处的实测值。
半方差模型的选取是基于实验半方差的值用不同类型的模型拟合得到的,拟合最优的模型要求拟合模型的决定系数(R2)较大,残差(RSS)较小。
模型中,C0是块金系数(间距为零时的半方差),表示随机部分的空间变异;C0+ C是基台值(半方差函数随着间距递增到一定程度后出现的平稳值),表示系统中的最大变异;C是结构方差(基台值与块金系数的差值);a是变程(指半方差变异函数达到基台值时所对应的距离),表示观测点之间的最大相关距离。结构比C/(C0 + C)表示自相关部分空间异质性占总空间异质性的程度,按自相关程度的分级标准:结构比C/(C0 + C) >75%,空间自相关性较强;25% <C/(C0 + C) <75%,空间自相关性中等; C/(C0+C) <25%,空间自相关性较弱,若结构比接近0,则表示变量在整个尺度上具有恒定的变异。
(4) 分形维数(D)[6]
用公式D=(4-m)/2计算,式中m为2γ(h)= h(4-2D)双对数回归直线的斜率。分形维数高,意味着空间分布格局简单,空间依赖性越强,空间结构性好;分形维数表示变异函数曲线的曲率大小,该值越接近2,表明越同质。
(5) Kriging插值法[28]
是一种最优无偏线性估值方法。Kriging插值及插值图的生成是由GS+9.0和Surfer 11.0共同完成的。选择Kriging最优内插法,利用区域化变量的原始数据和半方差函数的结构性,对未采样点的区域化变量的取值进行线性无偏最优估计,最终生成梭梭种群数量特征和海拔高度的空间分布图。
2 结果分析 2.1 梭梭林种群数量特征及样地海拔高度描述性统计分析对梭梭的密度、株高、冠幅直径及样点海拔高度观测数列进行K-S正态性检验,其阈值范围分别为S∈[-0.6172,0.6172]、K∈[-1.2344,1.2344],4个空间变量均符合正态分布,可以进行进一步数据分析。描述性统计分析结果表明梭梭林密度、株高、冠幅直径的变异系数分别为43.80%、36.40%、36.38%,均达到强度变异。除海拔高度外,其余3个指标的数据均偏向右侧分布,株高偏离程度最大;且株高和冠幅直径对称轴两侧的极端数据较多,密度和海拔高度对称轴两侧的极端数据较少(表 1)。
| 观测指标Index | 平均值Mean | 标准差STD | 变异系数C.V/% | 最小值MIN | 最大值MAX | 偏度Skewness | 峰度Kurtosis | D | D(110,0.05) | 正态分布Normal distribution |
| 密度Density/(株/200 m) | 10 | 4.38 | 43.80 | 1 | 19 | 0.0984 | -0.5309 | 0.0721 | 0.1297 | 符合 |
| 株高Height/cm | 124 | 45.13 | 36.40 | 31 | 236 | 0.2500 | 0.5086 | 0.0832 | 0.1297 | 符合 |
| 冠幅直径/cmCrown diameter | 152 | 55.29 | 36.38 | 44 | 302 | 0.1781 | 0.4033 | 0.0652 | 0.1297 | 符合 |
| 海拔高度/mAltitude | 1260.4 | 2.08 | 0.16 | 1256 | 1264 | -0.1283 | -0.5651 | 0.1226 | 0.1713 | 符合 |
梭梭林种群数量特征及样地海拔高度的空间变异函数见表 2,结果显示,种群数量特征及样地海拔高度的半方差最适函数模型均为Gaussian理论模型。冠幅直径基台值最高达5050,随机因素引起的空间变异最大;株高、密度、海拔高度结构比均超过75%,空间自相关性较强。海拔高度随机因素引起的空间变异最小,存在较强的空间自相关性,自相关尺度为1255.74 m。
| 观测指标Index | 模型Model | 块金值Nugget | 基台值Sill | 结构比Structure ratio | 范围参数Range parameters | 残差平方和Residual sum of squares | 决定系数Determinationcoefficient | 相关尺度Correlation scale |
| r(h) | C0 | C0+C | C/(C0+C) | a0 | RSS | R2 | Scale | |
| 密度Density/(株/200 m) | Gaussian | 6.1 | 28.21 | 0.784 | 909 | 0.164 | 0.998 | 1574.43 |
| 株高/cmHeight | Gaussian | 1008 | 4126 | 0.756 | 1249 | 3098 | 0.995 | 2163.33 |
| 冠幅直径/cmCrown diameter | Gaussian | 1470 | 5050 | 0.709 | 1035 | 18807 | 0.987 | 1792.67 |
| 海拔高度/mAltitude | Gaussian | 0.5 | 5 | 0.900 | 725 | 0.0635 | 0.990 | 1255.74 |
分形维数反映了复杂形体占有空间的有效性,是没有特征尺度的自相似结构[6]。在变异函数分析的基础上,进行分形维数计算(表 3)。可以看出,梭梭种群数量特征分形维数较高,均大于1.5,表明梭梭群落空间分布格局简单,空间依赖性强,空间结构性好。研究区海拔高度分形维数偏低,为1.442,空间同质度差。
| 观测指标 Index | 密度Density | 株高Height | 冠幅直径Crown diameter | 海拔高度Altitude |
| 分形维数Fractal dimension | 1.639 | 1.754 | 1.753 | 1.442 |
根据半方差函数和克里格插值方法,将梭梭种群数量特征及样点海拔高度绘成三维立体图(图 4,X、Y分别为样点的绝对坐标,代表样地的南方和西方),梭梭的分布以样地的北方最为密集(图 4),当密度范围高于11.7株/200 m时,其密度由最高区域向四周呈环形带状降低,当密度小于11.7株/200 m且大于4.9株/200 m时,梭梭的密度沿北侧和西侧同时呈条带状下降。株高和冠幅直径(图 4)空间变化规律相似,最大值均出现在西侧,且二者的位置大体相同,沿最高区域呈环形带状逐渐降低,当下降到一定程度(株高为158 cm、冠幅直径为153 cm),均同时沿着西侧和北侧呈条带状下降。海拔高度(图 4)以西侧和北侧的交汇处最高,并沿着最高区域呈带状逐渐下降,当海拔高度低于1260m时,其空间分布状态趋于复杂化。图形起伏剧烈程度反映梭梭林种群数量特征及样地海拔高度的空间分布异质性大小,分布不均匀,每个指标形成了一处大的峰值分布。由图 4知,4个指标分布图起伏强烈,说明空间分布比较强烈。对梭梭种群数量特征及样地海拔高度进行了相关性分析(表 4),梭梭自身种群数量特征之间存在极显著的相关性,其中株高和冠幅直径的相关系数最大(0.8771) 。梭梭种群生长特征与海拔高度之间存在极显著相关。

|
| 图 4 梭梭林种群数量特征及样地海拔高度变化趋势 Fig. 4 Changing trend of observation indicators and plots altitude of Haloxylon ammodendron |
| 观测指标Index | 密度Density | 株高Height | 冠幅直径Crown diameter | 海拔高度Altitude |
| 密度Density | 1.0000 | |||
| 株高Height | 0.5026** | 1.0000 | ||
| 冠幅直径Crown diameter | 0.4114** | 0.8771** | 1.0000 | |
| 海拔高度Altitude | 0.3624** | 0.4801** | 0.5950** | 1.0000 |
| **表示0.01水平下显著 | ||||
梭梭种群数量特征及海拔高度的半方差函数和分形维数结果不一致,在样地范围内,梭梭株高、冠幅直径、密度和海拔高度空间分布复杂程度依次递减。这主要是因为半方差函数分析需要先选择最适模型模拟,模型与纵轴的截距定义为块金方差,即随机性因素影响程度,且存在空间研究尺度效应;而分形维数首先不存在研究尺度效应,其只是表征空间自相关结构,通过数值大小判断影响因素的决定作用[29]。因此二者在研究决定性因素上存在差异,突出重点不同,半方差函数主要研究不同尺度上的空间结构,分形维数分析的是没有尺度的空间结构。
对在取样区域内的梭梭种群数量特征及样地海拔高度进行了相关性分析,梭梭本身的种群数量特征之间及其数量特征与海拔高度存在极显著正相关性。而对密度与株高之间相关性存在着不同的看法: Matthew J.[30]和Knowe[31]研究发现随林分密度的增大,株高有增大的趋势,本文研究与之研究结果基本一致;而段劼对不同密度梯度的北京山区侧柏人工幼龄林[32]的研究得出株高表现出随密度减小而有增大的趋势与本文研究结果截然相反;同样也有研究表明,一定密度范围内,Zhang[33]认为林木的密度对树高影响较弱或不显著。
Surfer进行空间分布分析时,选择趋势面回归的模型,将变量空间分布采用克里格插值通过图形表现出来,更为直观的判断变量空间分布特点和变化规律[28],样地的海拔高度以西侧和北侧的交汇处最高,梭梭的株高和冠幅直径(图 4)最大值均出现在西侧,且空间变化存在较强的一致性,密度(图 4)以样地的北方最为密集,均沿最高区域呈环形带状逐渐降低,当下降到一定程度同时沿着西侧和北侧呈条带状下降。分析原因为研究区盛行西北风和辅以东北风和西风,梭梭林的存在增大了下垫面的粗糙度,对外来风沙流具有明显的阻拦作用,当风沙流经过梭梭林时,有一部分气流在梭梭林西北部受阻而回旋,使风速急剧降低,风沙流的携沙能力降低,沙粒在梭梭林西北部沉落[34]。同时,由于梭梭林具有一定的疏透度,仍会有部分携沙气流则穿过灌木林带,受到梭梭林的摩擦、阻挡和分割,气流的能量削弱,部分沙物质会在梭梭林中发生沉降[35-36],从而导致西北角出现沙层厚度的最高区域,正北方受西北风和东北风合力影响,形成沙层厚度的次高区域。3个风向共同作用导致风沙流向东南推移,即形成不同沙丘高度:西北—西侧、北侧—东南方向逐渐降低的变化趋势,从而导致在小地形地貌海拔高度差异,表征的是沙层厚度。当沉降的沙层厚度未达到危害植株正常生长的情况,对植株的生长具有一定的促进作用,即适度的沙埋促进植物生长,可能原因是适度沙埋可能有利于保持植物根系周围的湿度,降低根系周围的土壤温度等,增加新沙子中的营养物质,改善微环境[37],增大根系的生长空间以及拓展的空间,增强了其根系吸收水分和养分的能力。
4 结论采用半方差函数可以较好地表征天然梭梭林的空间分布规律,梭梭株高、密度、冠幅直径均符合正态分布,达到强度变异;半方差最适函数理论模型均为高斯模型,在相应变程范围内,空间变异主要由结构性因素(空间自相关部分)引起。梭梭的种群数量特征空间依赖性强,空间结构简单,保持着相对较好的天然分布和生长状态。
研究区域内,由于长期风蚀作用,小地形海拔高度体现为不同沙层厚度,梭梭株高、冠幅直径和密度在空间上的分布呈西高东低,北高南低的趋势,与海拔高度存在极显著相关,表现出适度沙埋效应。
致谢: 感谢塔木素苏木政府工作人员在实验过程中的帮助及相关资料支持。| [1] | 陈丽, 王炜, 王东波, 王永利.扩展点格局分析方法在灌木种群空间分布格局研究中的应用.生态学杂志,2011,30(12): 2700–2705. |
| [2] | 闫海冰, 韩有志, 杨秀清, 王丽艳, 项小英.华北山地典型天然次生林群落的树种空间分布格局及其关联性.生态学报,2010,30(9): 2311–2321. |
| [3] | 李进军, 薛斌瑞, 柴宗政.民勤县荒漠-绿洲交错带梭梭种群的数量动态及分布格局.东北林业大学学报,2013,41(8): 27–31. |
| [4] | 白永飞, 许志信, 李德新.内蒙古高原针茅草原群落土壤水分和碳、氮分布的小尺度空间异质性.生态学报,2002,22(8): 1215–1223. |
| [5] | Titus J H, Nowak R S, Smith S D.Soil resource heterogeneity in the Mojave Desert.Journal of Arid Environments,2002,52(3): 269–292. |
| [6] | 胡旭, 王海涛, 卢建国, 丛培芳, 何兴东, 高玉葆, 董治宝.干旱和半干旱区油蒿对土壤空间异质性的响应.中国沙漠,2007,27(4): 588–592. |
| [7] | 岳文泽, 徐建华, 徐丽华, 谈文琦, 梅安新.不同尺度下城市景观综合指数的空间变异特征研究.应用生态学报,2005,16(11): 2053–2059. |
| [8] | 颜亮, 周广胜, 张峰, 隋兴华, 平晓燕.内蒙古荒漠草原植被盖度的空间异质性动态分析.生态学报,2012,32(13): 4017–4024. |
| [9] | Qi Y C, Dong Y S, Jin Z, Peng Q, Xiao S S, He Y T.Spatial heterogeneity of soil nutrients and respiration in the desertified grasslands of Inner Mongolia, China.Pedosphere,2010,20(5): 655–665. |
| [10] | Su Y Z, Li Y L, Zhao H L.Soil properties and their spatial pattern in a degraded sandy grassland under post-grazing restoration, Inner Mongolia, Northern China.Biogeochemistry,2006,79(3): 297–314. |
| [11] | 许皓, 李彦, 邹婷, 谢静霞, 蒋礼学.梭梭(Haloxylon ammodendron)生理与个体用水策略对降水改变的响应.生态学报,2007,27(12): 5019–5028. |
| [12] | 田媛, 塔西甫拉提·特依拜, 李彦, 唐立松, 范连连.梭梭幼苗的存活与地上地下生长的关系.生态学报,2014,34(8): 2012–2019. |
| [13] | 陈虞超, 李苗, 吴明朝, 宋玉霞.梭梭属两种植物的根结构和成分.植物生理学报,2013,49(11): 1273–1276. |
| [14] | 宋于洋, 李荣, 罗惠文, 张文辉.古尔班通古特沙漠三种生境下梭梭种群的生殖分配特征.生态学杂志,2012,31(4): 837–843. |
| [15] | 宋于洋, 李明艳, 张文辉.生境对古尔班通古特沙漠梭梭种群波动及谱分析推绎的影响.林业科学,2010,46(12): 8–14. |
| [16] | 宋于洋, 李园园, 张文辉.梭梭种群不同发育阶段的空间格局与关联性分析.生态学报,2010,30(16): 4317–4327. |
| [17] | 张龙. 梭梭生根条件的优化及生根过程中的生理学与解剖学研究[D]. 石河子:石河子大学, 2013. |
| [18] | 孙鹏飞, 周宏飞, 李彦, 李妙伶.古尔班通古特沙漠原生梭梭树干液流及耗水量.生态学报,2010,30(24): 6901–6909. |
| [19] | 吕朝燕, 张希明, 刘国军, 吴俊侠, 闫海龙, 邓潮洲.准噶尔盆地西北缘梭梭种群结构和空间格局特征.中国沙漠,2012,32(2): 380–387. |
| [20] | 刘国军, 张希明, 李建贵, 范冬冬, 邓潮洲, 侯建国, 信汝明.供水量及沙埋厚度对两种梭梭出苗的影响.中国沙漠,2010,30(5): 1085–1091. |
| [21] | 王继和, 张锦春, 袁宏波, 廖空太, 刘虎俊, 张国中.库姆塔格沙漠梭梭群落特征研究.中国沙漠,2007,27(5): 809–813. |
| [22] | 王春玲, 郭泉水, 谭德远, 史作民, 马超.准噶尔盆地东南缘不同生境条件下梭梭群落结构特征研究.应用生态学报,2005,16(7): 1224–1229. |
| [23] | 张锦春, 王继和, 安富博, 孙涛, 刘有军, 李银科, 肖斌.民勤天然梭梭种群特征初步研究.中国沙漠,2009,29(6): 1124–1128. |
| [24] | 常静, 潘存德, 师瑞锋.梭梭-白梭梭群落优势种种群分布格局及其种间关系分析.新疆农业大学学报,2006,29(2): 26–29. |
| [25] | 马兴华, 张晋昕.数值变量正态性检验常用方法的对比.循证医学,2014,14(2): 123–128. |
| [26] | 薛毅, 陈立萍. 统计建模与R软件. 北京: 清华大学出版社, 2007. |
| [27] | Gifford G F, Hawkins R H.Hydrologic impact of grazing on infiltration:a critical review.Water Resources Research,1978,14(2): 305–313. |
| [28] | 赵彦锋, 孙志英, 陈杰.Kriging插值和序贯高斯条件模拟算法的对比分析.地球信息科学学报,2010,12(6): 767–776. |
| [29] | 刘红梅. 短花针茅草原群落特征与草地空间异质性对不同放牧制度的响应[D]. 呼和浩特:内蒙古农业大学, 2011. |
| [30] | Waghorn M J, Mason E G, Watt M S.Influence of initial stand density and genotype on longitudinal variation in modulus of elasticity for 17-year-old Pinus radiate.Forest Ecology and Management,2007,252(1/3): 67–72. |
| [31] | Knowe S A, Hibbs D E.Stand structure and dynamics of young red alder as affected by planting density.Forest Ecology and Management,1996,82(1/3): 69–85. |
| [32] | 段劼, 马履一, 贾黎明, 徐程扬, 贾忠奎, 车文瑞.北京地区侧柏人工林密度效应.生态学报,2010,30(12): 3206–3214. |
| [33] | Zhang J W, Oliver W W, Ritchie M W.Effect of stand densities on stand dynamics in white fir (Abies concolor) forests in northeast California, USA.Forest Ecology and Management,2007,244(1/3): 50–59. |
| [34] | 王翠, 李生宇, 雷加强, 毛东雷, 周杰.近地表风沙流结构对过渡带不同下垫面的响应.水土保持学报,2014,28(3): 52–56. |
| [35] | Wolfe S A, Nickling W G.The protective role of sparse vegetation in wind erosion.Progress in Physical Geography,1993,17(1): 50–68. |
| [36] | 丁国栋. 风沙物理学(第二版). 北京: 中国林业出版社, 2007. |
| [37] | 赵哈林, 赵学勇, 张铜会. 沙漠化的生物过程及退化植被的恢复机理. 北京: 科学出版社, 2007. |