 2015, Vol. 36
2015, Vol. 36文章信息
- 刘晓娟, 孙学刚, 田青
- LIU Xiaojuan, SUN Xuegang, TIAN Qing.
- 垫状植物囊种草对群落物种多样性的影响
- Effect of the cushion plant Thylacospermum caespitosum Camb. on species diversity within a community
- 生态学报[J]. 2015, 36(10): 2905-2913
- Acta Ecologica Sinica[J]. 2015, 36(10): 2905-2913
- http://dx.doi.org/10.5846/stxb201411042163
-
文章历史
- 收稿日期: 2014-11-04
- 网络出版日期: 2015-09-28

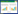




非生物环境条件和物种间的相互关系可以决定自然群落中的物种多样性模式[1-4]。然而,尽管所有的有机体与他们的物理环境相互作用,但其中的一些会通过非营养途径改变环境条件和资源可利用性,从而影响其他物种,进而改变整个群落的性质[5-10]。Jones[11-12]提出了“生态系统工程”一词来指示这种生境改变的生物过程,和“生态系统工程师”来指示那些造成生境改变的有机体。高山垫状植物具有修饰其物理环境的能力[13-17]。例如,与周围暴露的环境相比,垫状植物可以减少98%的风速[13],从而减少由此产生的热损失,使垫状植物内维持比外界环境高的温度[17],为那些接近低温下限的植物提供适合的生境斑块[16]。另外,垫状植物可以维持比周围环境优越的水分[14, 18]和养分条件[15, 18]。另一方面,垫状创造了大的生境斑块,直径可超过3 m,存活几十年[19]甚至上百年[20]。因此,垫状植物能改变生境的能力,加上他们所创造的大而且持久的构造,表明这些生长形式可以作为生态系统工程师[21-24]。垫状植物对微环境的修饰可以提高其他非垫状植物的存活率和生长表现[18,25-26],影响自然群落中其他物种的发生率和多度。Cavieres等[18]通过一系列的移栽实验证明,在垫状植物内部生长的幼苗Cerastium arvense和Hordeum comosum的成活率分别为大约30%和80%,而在非垫状植物条件下其生长幼苗的成活率均低于10%。其他一些研究结果也表明,通过对比同一种植物的生存几率、丰富度以及光合效率,与对照区域相比垫状植物可以促进植物的生存[27-28]。在恶劣的高山环境中,由于垫状植物能够提供相对优越的微环境,一方面成为其他植物生存首选的场所,为群落中增加了新的种类从而增加了群落的丰富度。另一方面也影响了群落中现有物种的丰富度,从而影响群落的均匀度[27]。研究表明垫状植物对植物群落物种丰富度、均匀度和多样性有促进作用,这说明垫状植物对高山植物群落多样性的维持有重要作用[15, 16, 21, 27, 29-32]。
然而,关于垫状植物如何通过改变物种丰富度或均匀度进而影响群落多样性的研究明显不足,且大多数研究并未进行量化研究。囊种草(Thylacospermum caespitosum Camb.)(石竹科)是广泛分布于我国青藏高原高山寒漠带的垫状植物,具有典型的垫状体结构,在高山寒漠带稀疏垫状植被中极具代表性。研究发现[33-34],囊种草可对其覆盖下的土壤微环境起到明显的改善作用,可提高其覆盖下土壤养分含量和土壤含水量,改善土壤通气状况,并且维持其冠层下温度处于相对恒定的范围内,为其自身和其他植物提供了相对优越的微环境。因此,研究了垫状植物囊种草对群落物种多样性的影响,并对其影响程度和潜力进行了量化研究,以期揭示囊种草如何通过改变群落物种丰富度和均匀度进而影响群落物种多样性。
1 研究区概况与研究方法 1.1 研究区概况研究在位于青藏高原北缘的甘肃盐池湾国家级自然保护区内进行,于2012年7—9月展开,7—9月为研究区内植物的生长盛季。保护区内最低海拔2600 m,最高海拔5483 m,山脊多在海拔4000 m以上。该保护区内的高山海拔在永久雪线以上,分布着现代冰川,雪线以下则为高山寒漠带,分布有稀疏垫状植被。在保护区选择典型的囊种草垫状植被设置20 m×40 m研究样地,样地地理位置39°05′10.7″N,96°46′19.5″E,海拔4137 m。样地年均温-3.8 ℃,1月均温-17.0 ℃,7月均温8.6 ℃,年均降水量255.6 mm,年均蒸发量2493.3 mm。样地优势种为囊种草(Thylacospermum caespitosum Camb.)(石竹科),直径大多在10—60 cm。伴生其他一年生和多年生草本。
1.2 研究对象囊种草为多年生垫状草本,常呈球形,全株无毛。茎基部强烈分枝,木质化。叶排列紧密,呈覆瓦状,叶片卵状披针形,长2—4 mm,宽约2 mm,顶端短尖,质硬,有光泽。花单生茎顶,几无梗;花瓣5,卵状长圆形;雄蕊10,短于萼片;花柱3,线形,常伸出萼外。蒴果球形,直径2.5—3 mm,黄色,具光泽,6齿裂。花期6—7月,果期7—8月。分布海拔3600—6000 m[35]。
1.3 研究方法 1.3.1 囊种草对群落物种组成的影响为了控制面积效应,取样前先在样地内设置5条1 m×40 m的平行样带,测定囊种草在群落中的盖度,测定结果显示囊种草在样地内的盖度为15%。在样地内随机选取60个任意大小的囊种草,测量其长径和短径,记录其冠层内分布的其他植物的种类及个体数。对于任一选定的囊种草,用铁丝沿其边缘围出轮廓,然后将铁丝圈放置在远离该囊种草50 cm以上的地方(且此处周围50 cm范围内也无其他任何垫状植物)作为对照斑块,按照盖度比例,共选取340个对照斑块,记录对照斑块内出现的植物的种类和个体数。根据取样斑块数和物种数的关系绘制曲线来判断是否充分调查了群落中的物种丰富度和组成。垫状植物囊种草的面积用公式:面积=(π×长径×短径)/4来计算。
1.3.2 囊种草对群落物种多样性的影响为测定生态系统工程师对群落性质的影响,Wright[36]提出首先要测定没有受到工程师影响的群落性质(未修饰生境斑块),然后将其和被工程师修饰生境斑块的群落性质进行比较(改造后景观=未修饰+被修饰生境斑块)。因此,以对照区域作为未被囊种草改造的生境,以囊种草和对照区域相加作为被囊种草改造后的生境,比较生境在被囊种草改造前后的物种组成、物种丰富度、密度(以单位面积个体数表示)和均匀度的差异。均匀度采用以下公式[37]进行测度:
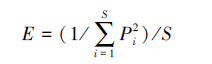
|
式中,E为均匀度指数,Pi为种i的多度比例,S为种的总数。物种丰富度采用Margalef指数[37]:
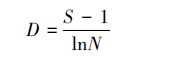
|
式中,D为物种丰富度指数,S为种的总数,N为全部种的个体总数。信息指数采用Shannon-Wiener指数[37]:
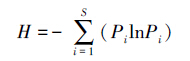
|
式中,H为信息指数,S为种的总数,Pi为种i的多度比例。 所测数据用SPSS11.0软件进行差异显著性分析。
1.3.3 囊种草对群落物种丰富度的影响按照种-面积关系确定最少调查斑块数。将以上调查的所有植物(除囊种草)进行物种鉴定,编制样地植物名录,按照这些植物出现的生境类型对其分类[21]。将囊种草群落的环境划分为两种类型:被囊种草改造过的生境斑块(即囊种草本身及其周围环境)和未被囊种草改造过的生境斑块(即无囊种草生长的地方,即对照)。根据生境类型将植物分为只存在于囊种草内部的植物种类,即囊种草特有种(STc),只存在于对照的植物种类,即对照特有种(SCK)和两种生境中都有的植物种类,即生境一般种(SCL)。
通过两个指标定量测定囊种草对群落物种丰富度的影响[21],在斑块水平测度相对生境物种丰富度(RHR),RHR=(STc + SCL)/(SCK + SCL);在景观水平测度群落物种丰富度增加效应(LRE),LRE= STc /(SCK + SCL)。另外选取两个指标测定囊种草对群落物种丰富度稳定性的维持能力,在斑块水平测度囊种草对周围环境的侵占潜力(HRP),HRP= STc (CL) / SCL,STc(CL)为囊种草内具有较高多度的SCL;在景观水平测度囊种草对群落物种丰富度的维持潜力(LIP),LIP= SCL / (SCL + STc + SCK)。
2 结果与分析 2.1 囊种草对群落物种组成的影响 2.1.1 植物种类对样地内分布的所有植物进行了物种鉴定,编制出样地植物名录(表1)。根据调查斑块数和植物种数的关系绘制曲线(图1),从图中可以看出,当囊种草取样数大于45个,对照斑块取样数大于55个,群落斑块取样数大于55个时,调查植物种类不再增加,表明本研究的取样做到了充分取样。调查发现样地中共分布有植物20种,隶属于11科,其中以十字花科和菊科的种类居多。在所调查的囊种草中共发现17个植物种类,在对照区域中生长的植物为16种,单纯就植物种类来说,囊种草内生长的植物种类多于无囊种草的对照区域。
| 科名 Family | 物种名 Species | 生长型 Growth form | 出现生境 Habitat |
| 毛茛科Ranunculaceae | 单花翠雀花Delphinium candela var. monanthum | 多年生草本 | CL |
| 十字花科Cruciferae | 红花肉叶荠Braya rosea | 多年生草本 | CL |
| 单花荠Pegaeophyton scapiflorum | 多年生草本 | CL | |
| 藏荠Hedinia tibetica | 多年生草本 | CL | |
| 紫花糖芥Erysimum funiculosum | 多年生草本 | CL | |
| 豆科Leguminosae | 马衔山黄耆Astragalus mahoschanicus | 多年生草本 | CL |
| 石竹科Caryophyllaceae | 沙生繁缕Stellaria arenaria | 多年生草本 | CL |
| 罂粟科Papaveraceae | 细果角茴香Hypecoum leptocarpum | 一年生草本 | Tc |
| 龙胆科Gentianaceae | 矮假龙胆Gentianella pygmaea | 一年生草本 | CL |
| 紫红假龙胆Gentianella arenaria | 一年生草本 | CK | |
| 合萼肋柱花Lomatogonium gamosrpalum | 一年生草本 | Tc | |
| 唇形科Labiatae | 白花枝子花Dracocephalum heterophyllum | 多年生草本 | CK |
| 紫草科Boraginaceae | 小花西藏微孔草Microula tibetica var. pratensis | 二年生草本 | CK |
| 菊科Compositae | 肉叶雪兔子Saussurea thomsonii | 多年生草本 | CL |
| 无梗风毛菊Saussurea apus | 多年生草本 | CL | |
| 高山紫菀Aster alpinus | 多年生草本 | CL | |
| 黄白火绒草Leontopodium ochroleucum | 多年生草本 | Tc | |
| 碱韭Allium polyrhizum | 多年生草本 | Tc | |
| 百合科Liliaceae | 粗壮嵩草Kobresia robusta | 多年生草本 | CL |
| 禾本科Gramineae | 堇色早熟禾Poa araratica subsp. ianthina | 多年生草本 | CL |
| CL:两种生境类型,即群落水平 both habitats,community level;Tc:囊种草 Thylacospermum caespitosum Camb.;CK:对照区域 control area | |||

|
| 图 1 种-取样斑块关系 Fig. 1 Relationship between species richness and number of samples |
调查发现(表1),20种植物中,只生长于囊种草内部的植物,即囊种草特有种为4种,一年生草本细果角茴香和合萼肋柱花,以及多年生草本黄白火绒草和碱韭;只生长于对照区域的植物,即对照特有种为3种,一年生草本紫红假龙胆,二年生草本小花西藏微孔草和多年生草本白花枝子花;两种生境中都有的植物,即生境一般种为13种,除矮假龙胆为一年生草本外其余均为多年生草本。对于生境特有种,囊种草内的特有种也多于对照区域。
对于样地中的生境一般种,即在囊种草内和无垫状植物区域均存在的13个种,比较了其在囊种草出现前后的多度差异,以密度来表示(表2)。结果表明,13个生境一般种中,有4个种在被囊种草改造后的生境中具有更高的多度,包括一年生草本矮假龙胆和多年生草本单花翠雀花、红花肉叶荠和马衔山黄耆。本研究中,囊种草并未提高其内部生长的大多数物种的多度,仅表现出对于一年生草本多度的促进效应,调查发现的4种一年生草本中,有2种仅出现在囊种草中,1种出现在两种生境类型中,但在囊种草内多度更高。
Núñez C等[15]研究表明,一些一年生小草本更趋向于出现在物种较为丰富的垫状植物内部,而一些多年生草本更趋于单独出现或出现在物种丰富度较低的斑块。另外,Núez C等[15]也发现,垫状植物生长非常缓慢,1个新种出现在垫状植物内需要6—7a,因此垫状植物内的物种积累也需要一个漫长的过程,而在此过程中,一年生草本则更容易出现在垫状植物内。因此,尽管大多数种在垫状植物内和无垫状植物的区域均存在,但垫状植物的出现由于改善了环境条件,进而导致了某些生境一般种的多度。同样的结果也在其他一些研究中被报道[5, 38-41]。
| 植物种类 Species | 生长型 Growth form | 密度Density/(株/m2) | |
| 被垫状植物改造前 Without cushions | 被垫状植物改造后 With cushions | ||
| 单花翠雀花Delphinium candelabrum var. monanthum (Hand. -Mazz.) W. T. Wang | 多年生草本 | 0.71 | 0.75 |
| 红花肉叶荠Braya rosea (Turcz.) Bunge | 多年生草本 | 10.15 | 10.77 |
| 单花荠Pegaeophyton scapiflorum (Hook. f. et Thoms.) Marq. et Shaw | 多年生草本 | 8.27 | 8.26 |
| 藏荠Hedinia tibetica (Thoms.) Ostenf. | 多年生草本 | 4.62 | 4.56 |
| 紫花糖芥Erysimum chamaephyton Maxim. | 多年生草本 | 8.08 | 7.58 |
| 马衔山黄耆Astragalus mahoschanicus Hand. -Mazz. | 多年生草本 | 1.90 | 1.96 |
| 沙生繁缕Stellaria arenaria Maxim. | 多年生草本 | 21.65 | 21.04 |
| 矮假龙胆Gentianella pygmaea (Regel et Schmalh.) H. Smith apud S. Nilsson | 一年生草本 | 8.12 | 8.68 |
| 肉叶雪兔子Saussurea thomsonii C. B. Clarke | 多年生草本 | 28.25 | 27.40 |
| 无梗风毛菊Saussurea apus Maxim. | 多年生草本 | 33.72 | 31.96 |
| 高山紫菀Aster alpinus L. | 多年生草本 | 14.74 | 14.65 |
| 粗壮嵩草Kobresia robusta Maxim. | 多年生草本 | 28.91 | 27.99 |
| 堇色早熟禾Poa ianthina Keng ex H. L. Yang | 多年生草本 | 21.32 | 20.33 |
通过比较无囊种草影响的生境(对照)和被囊种草改造过的生境(囊种草斑块+对照)间的物种密度、物种丰富度、均匀度和信息指数的差异来测度囊种草对群落物种多样性的影响。研究发现在无囊种草时群落的物种密度为0.34种/ m2,被囊种草影响后的群落物种密度为0.36种/ m2,表明囊种草的出现提高了群落物种密度(表3)。物种丰富度指数选用Magalef指数,该指数仅以群落中的种树和个体总数的关系为基础,而无需考虑研究面积的大小。计算结果显示,被囊种草改造前群落物种丰富度为1.63,被囊种草改造后的群落物种丰富度为2.04(表3),说明囊种草提高了群落物种的丰富度。许多研究都仅使用物种丰富度来表示生物多样性,然而一些研究表明物种丰富度和物种均匀度之间不一定存在正相关关系[42]。对于均匀度的计算结果显示,囊种草降低了群落物种的均匀度,从改造前的0.62降低到了0.50(表3)。此外还选择了Shannon-Wiener指数从群落中种的个体出现的不确定性角度来测度物种多样性,结果同样表明囊种草提高了群落物种多样性。
| 指标 Indexes | 被囊种草改造前 Without cushions | 被囊种草改造后 With cushions |
| 物种密度 Species density/(种/m2) | 0.34 | 0.36 |
| Magalef指数Magalef index | 1.63 | 2.04 |
| 均匀度Eveness | 0.62 | 0.50 |
| Shannon-Wiener指数 Shannon-Wiener index | 2.45 | 2.49 |
对于生态系统工程师对群落多样性影响的量化研究越来越受到关注[8, 21, 43-47],尤其是生态系统工程师对物种丰富度影响的量化研究[21, 36]。Badano等[21]提出了用于测度生态系统工程师对群落物种丰富度影响的量化指标,我们将这些指标引入研究中,用指标的测定值更为直观的说明囊种草对群落物种丰富度的影响。
2.2.1 垫状植物对群落物种丰富度的影响分析被囊种草改造过的生境斑块中是否具有比裸地更多、更少或同样的物种数量,引入指标—相对生境物种丰富度(RHR)(斑块水平)和群落物种丰富度增加效应(LRE)(景观水平)。测定结果如表4所示,RHR测定值为1.06,LRE测定值为0.25。根据公式,当RHR=1时,两种生境具有相同种类;当RHR <1时,表示被囊种草改造过的生境中比无囊种草生长的对照区域具有更少的植物种类;本研究中RHR>1,表示被囊种草改造过的生境中比对照区域具有更多的植物种类,囊种草的出现提高了群落物种丰富度。而LRE反映了囊种草为群落引入新种的程度,如果在囊种草中至少出现一个特有种,那么囊种草斑块的增加将会引起景观水平物种丰富度的增加。当LRE=0时,表明没有只存在于囊种草内的植物种,当只存在于囊种草中的植物种类越多,LRE越大,则随着LRE的增加,表明有更多的物种依赖于被囊种草改造过的环境,本研究中LRE为0.25,表明有少数种类,如细果角茴香、合萼肋柱花、黄白火绒草和碱韭均依赖于被囊种草改造过的环境,而大多数种类更趋向于分布在两种生境中,但囊种草的出现仍然提高了群落物种丰富度。
| 指标 Indexes | 公式 Formula | 测定值 Values |
| 相对生境物种丰富度 Relative habitat richness(RHR) | RHR=(STC+ SCL)/(SCK+ SCL) | 1.06 |
| 群落物种丰富度增加效应 Landscape richness enhancment(LRE) | LRE= STC /(SCK + SCL) | 0.25 |
| 侵占潜力 Habitat rescue potential(HRP) | HRP= STC (CL)/ SCL | 0.23 |
| 群落物种丰富度维持潜力 Landscape insurance potential (LIP) | LIP= SCL /(SCL + STC + SCK) | 0.65 |
如果囊种草能够增加群落物种的丰富度,那么他是否能够将增加的物种丰富度维持下去,或者是否具有持续增加物种丰富度的潜力。于是,引入指标HRP——垫状植物对周围环境的侵占潜力(斑块水平)和LIP——垫状植物对群落物种丰富度的维持潜力(景观水平)。经计算,HRP为0.23,LIP为0.65(表4)。根据公式,当HRP=0时,没有生境一般种在囊种草中具有较高的多度,表明囊种草对周围环境的侵占潜力最小。测定结果表明部分植物种类在囊种草中具有较高的多度,即囊种草的出现为这些种类的生存起到了加固的作用,降低了他们的灭绝风险。类似的效应在景观水平也存在,囊种草创造了和其周围不一样的生存环境,因此,当外界环境压力发生变化时,囊种草和其周围生境会发生不同的变化,两种生境类型的环境条件也会不同程度的波动,假设生境一般种在两种生境中均可存活,那么这些种类如果在一种生境中灭绝则还可能在另一生境中存活,于是,囊种草所创造的生境多样性则成为一种保障,重新维持景观中物种丰富度从而降低物种损失的风险。那么,如果在两种斑块中都存在的种越多,则群落物种丰富度维持的潜力就越大。当群落中所有种均为生境一般种时,LIP达到最大值1,所测定的LIP值为0.65,表明囊种草对群落物种丰富度的维持潜力较高。
3 讨论研究结果表明,囊种草的出现导致了景观中新的生境斑块的创造和维持,这些新的生境斑块具有与周围无囊种草的生境基质十分不同的条件[33-34],而这种生境条件的改变影响了其他物种的分布,提高了整个群落的物种多样性。本研究中囊种草的出现为群落中增加了新的物种,改变了群落物种组成并提高了群落物种丰富度,同时也改变了部分种类的多度,而且囊种草也表现出能够维持物种丰富度的潜力。
对于生物多样性指标的测度表明,囊种草的出现提高了群落物种丰富度,这与大多数的研究结果一致[15, 21, 27]。囊种草由于能够提供相对优越的微环境[33-34],一方面成为其他植物生存的首选场所,为群落中增加了新的种类从而增加群落的丰富度;另一方面也影响了群落中现有物种的丰富度,增加了群落物种多样性[21, 32]。本研究中,囊种草降低了群落物种均匀度,这一结果与大多数的研究结果相反[27]。这可能是由于每个被调查的斑块中的物种数目较少造成的,被调查的囊种草斑块中最多出现的物种数为7个。此外囊种草对于某些植物种类的正向作用使的这些种在囊种草内的多度增加。并且在囊种草斑块中出现了4个囊种草特有种,这4个种仅分布在囊种草内。因此,囊种草的出现为群落中增加了新的物种,改变了某些种的分布和多度,因而造成了群落物种均匀度的降低。
在高山寒漠带严酷的环境条件下,囊种草的出现改善了土壤环境条件[33-34],使得一些原本难以在这样的环境中生长的植物种类能够存活下来[15, 16, 32],从而提高了群落物种多样性。另外,囊种草对于土壤环境条件的改善为景观中创造和维持了新的生境斑块,使得生境多样性提高,进而也导致了物种多样性的提高[12]。有研究发现,被垫状植物修饰后的环境能够保护植物幼苗免受低温和干旱的伤害,提高植物幼苗的成活率[48],因此也提高了一些种类在垫状植物内的多度和生长表现。本研究样地中,有65%的种类既生长在垫状植物囊种草内,也生长在无囊种草的区域,这表示囊种草的出现为这些种类提供了多一种的生境。在高山环境中,植物在生长季节的死亡往往是由于不可预测的非生物事件导致的,包括土壤温度的波动、干旱、雪水径流侵蚀造成的土壤扰动等[49-50]。而垫状植物能够调节土壤温度,减小土壤温度的波动,维持较高的土壤水分,加上垫状植物紧实的垫状体结构和深根性,使得这些非生物事件在垫状植物内发生的可能性会小一些。因而垫状植物成为了现有的群落物种多样性维持的一种保障,同时在以上事件发生时,为其他物种提供了避难场所,降低了物种损失的风险。
| [1] | Connell J H, Orias E.The ecological regulation of species diversity.The American Naturalist,1964,98(903): 399–414. |
| [2] | Connell J H.Diversity in tropical rain forests and coral reefs.Science,1978,199(4335): 1302–1310. |
| [3] | Tilman G D.Diversity by default.Science,1999,283(5401): 495–496. |
| [4] | Tilman G D, Lehman C L, Thomson K T.Plant diversity and ecosystem productivity: theoretical considerations.Proceedings of the National Academy of Sciences of the United States of America,1997,94(5): 1857–1861. |
| [5] | Flecker A S.Ecosystem engineering by a dominant detritivore in a diverse tropical stream.Ecology,1996,77(6): 1845–1854. |
| [6] | Aho K, Huntly N, Moen J, Oksanen T.Pikas (Ochotona princeps: Lagomorpha) as allogenic engineers in an alpine ecosystem.Oecologia,1998,114(3): 405–409. |
| [7] | Berkenbusch K, Rowden A A.Ecosystem engineering - moving away from 'just-so' stories.New Zealand Journal of Ecology,2003,27(1): 67–73. |
| [8] | Lill J T, Marquis R J.Ecosystem engineering by caterpillars increases insect herbivore diversity on white oak.Ecology,2003,84(3): 682–690. |
| [9] | McCabe D J, Gotelli N J.Caddisfly diapause aggregations facilitate benthic invertebrate colonization.Journal of Animal Ecology,2003,72(6): 1015–1026. |
| [10] | Flecker A S, Taylor B W.Tropical fishes as biological bulldozers: density effects on resource heterogeneity and species diversity.Ecology,2004,85(8): 2267–2278. |
| [11] | Jones C G, Lawton J H, Shachak M.Organisms as ecosystem engineers.Oikos,1994,69(3): 373–386. |
| [12] | Jones C G, Lawton J H, Shachak M.Positive and negative effects of organisms as physical ecosystem engineers.Ecology,1997,78(7): 1946–1957. |
| [13] | Hager J, Faggi A M.Observaciones sobre distribución y microclima de cojines enanos de la isla de Cretay del noroeste de la Patagonia.,1990,6 109–127. |
| [14] | Cavieres L A, Penaloza A P G, Papic C.Revista Chilena de Historia Natural,1998,71 337–347. |
| [15] | Núñez C, Aizen M, Ezcurra C.Species associations and nurse plant effect in patches of high-Andean vegetation.Journal of Vegetation Science,1999,10(3): 357–364. |
| [16] | Arroyo M T K, Cavieres L A, Peñaloza A, Arroyo-Kalin M A.Positive associations between the cushion plant Azorella monantha (Apiaceae) and alpine plant species in the Chilean Patagonian Andes.Plant Ecology,2003,169(1): 121–129. |
| [17] | Körner C. Alpine Plant Life.2nd ed. Berlin: Springer Press, 2003. |
| [18] | Cavieres L A, Badano E I, Sierra-Almeida A, Gómez-González S, Molina-Montenegro M A.Positive interactions between alpine plant species and the nurse cushion plant Laretia acaulis do not increase with elevation in the Andes of central Chile.New Phytologist,2006,169(1): 59–69. |
| [19] | Mark A F, Wilson J B.Tempo and mode of vegetation dynamics over 50 years in a New Zealand alpine cushion / tussock community.Journal of Vegetation Science,2005,16(2): 227–236. |
| [20] | Kleier C, Rundel P W.Microsite requirements, population structure and growth of the cushion plant Azorella compacta in the tropical Chilean Andes.Austral Ecology,2004,29(4): 461–470. |
| [21] | Badano E I, Cavieres L A.Ecosystem engineering across ecosystems: do engineer species sharing common features have generalized or idiosyncratic effects on species diversity? Journal of Biogeography, 2006, 33(2): 304-313.Journal of Biogeography,2006,33(2): 304–313. |
| [22] | Antonsson H, Bjork R G, Molau U.Nurse plant effect of the cushion plant Silene acaulis (L.Plant Ecology and Diversity,2009,2(1): 17–25. |
| [23] | Cavieres L A, Badano E I.Do facilitative interactions increase species richness at the entire community level? Journal of Ecology, 2009, 97(6): 1181-1191.Journal of Ecology,2009,97(6): 1181–1191. |
| [24] | Badano E I, Marquet P A, Cavieres L A.Predicting effects of ecosystem engineering on species richness along primary productivity gradients.Acta Oecologica,2010,36(1): 46–54. |
| [25] | SklenářP.Presence of cushion plants increases community diversity in the high equatorial Andes.Flora: Morphology, Distribution, Functional Ecology of Plants,2009,204(4): 270–277. |
| [26] | Callaway R M, Brooker R W, Choler P, Kikvidze Z, Lortie C J, Michalet R, Paolini L, Pugnaire F I, Newingham B, Aschehoug E T, Armas C, Kikodze D, Cook B J.Positive interactions among alpine plants increase with stress.Nature,2002,417(6891): 844–848. |
| [27] | Badano E I, Cavieres L A.Impacts of ecosystem engineers on community attributes: Effects of cushion plants at different elevations of the Chilean Andes.Diversity and Distribution,2006,12(4): 388–396. |
| [28] | Cavieres L A, Quiroz C L, Molina-Montenegro M A, Muñoz A A, Pauchard A.Nurse effect of the native cushion plant Azorella monantha on the invasive non-native Taraxacum officinale in the high-Andes of central Chile.Perspectives in Plant Ecology, Evolution and Systematics,2005,7(3): 217–226. |
| [29] | Badano E I, Marquet P A.Biogenic habitat creation affects biomass-diversity relationships in plant communities.Perspectives in Plant Ecology, Evolution and Systematics,2009,11(3): 191–201. |
| [30] | Le Roux P C, McGeoch M A.The use of size as an estimator of age in the subantarctic cushion plant, Azorella selago (Apiaceae).Arctic, Antarctic, and Alpine Research,2004,36(4): 509–517. |
| [31] | Molina-Montenegro M A, Torres C, Parra M J, Caviares L A.Asociación de especies al cojín Azorella trifurcata (Gaertn.) Hook. (Apiaceae) en la zona andina de Chile central (37 °S),2000,57 161–168. |
| [32] | Cavieres L A, Arroyo M T K, Peñaloza A, Molina-Montenegro M, Torres C.Nurse effect of Bolax gummifera cushion plants in the alpine vegetation of the Chilean Patagonian Andes.Journal of Vegetation Science,2002,13(4): 547–554. |
| [33] | 刘晓娟, 陈年来, 田青.海拔对囊种草(Thylacospermum caespitosum)修饰土壤微环境的影响.中国沙漠,2014,34(1): 191–196. |
| [34] | 刘晓娟, 陈年来, 田青.两种类型垫状植物对土壤微环境修饰作用的比较.草业学报,2014,23(1): 123–130. |
| [35] | 《中国植物志》编委会. 中国植物志: 第二十六卷. 北京: 科学出版社, 1996250–250. |
| [36] | Wright J P, Jones C G, Flecker A S.An ecosystem engineer, the beaver, increases species richness at the landscapescale.Oecologia,2002,132(1): 96–101. |
| [37] | 张金屯. 数量生态学. 北京: 科学出版社, 2004. |
| [38] | Thomas F, Renaurd F, de Meeûs T, Poulin R.Manipulation of host behavior by parasites: ecosystem engineering in the intertidal zone? Proceedings of the Royal Society of London, Series B, 1998, 265(1401): 1091-1096.Proceedings of the Royal Society of London, Series B,1998,265(1401): 1091–1096. |
| [39] | Ceballos G, Pacheco J, List R.Influence of prairie dogs (Cynomys ludovicianus) on habitat heterogeneity and mammalian diversity in Mexico.Journal of Arid Environments,1999,41(2): 161–172. |
| [40] | Crooks J A, Khim H S.Architectural vs.Journal of Experimental Marine Biology and Ecology,1999,240(1): 53–75. |
| [41] | Schooley R L, Bestelmeyer B T, Kelly J F.Influence of small-scale disturbances by kangaroo rats on Chihuahuan Desert ants.Oecologia,2000,125(1): 142–149. |
| [42] | Buzas M A, Heyek L A C.Biodiversity resolution: an integrated approach.Biodiversity Letters,1996,3(2): 40–43. |
| [43] | Fukui A.Indirect interactions mediated by leaf shelters in animal-plant communities.Population Ecology,2001,43(1): 31–40. |
| [44] | Wilby A.Ecosystem engineering: a trivialized concept? Trends in Ecology & Evolution, 2002, 17(7): 307-307.Trends in Ecology & Evolution,2002,17(7): 307–307. |
| [45] | Castilla J C, Lagos N A, Cerda M.Marine ecosystem engineering by the alien ascidian Pyura praeputialis on a mid-intertidal rocky shore.Marine Ecology Progress Series,2004,268 119–130. |
| [46] | Fogel B N, Crain C M, Bertness M D.Community level engineering effects of Triglochin maritima (seaside arrowgrass) in a salt marsh in northern New England, USA.Journal of Ecology,2004,92(4): 589–597. |
| [47] | Wright J P, Jones C G.Predicting the effects of ecosystem engineers on patch-scale species richness from primary productivity.Ecology,2004,85(8): 2071–2081. |
| [48] | Cavieres L A, Badano E I, Sierra-Almeida A, Molina-Montenegro M A.Microclimatic modifications of cushion plants and their consequences for seedling survival of native and non-native herbaceous species in the high Andes of central Chile.Arctic, Antarctic, and Alpine Research,2007,39(2): 229–236. |
| [49] | Chambers J C.Disturbance, life history strategies, and seed fates in alpine herbfield communities.American Journal of Botany,1995,82(3): 421–433. |
| [50] | Forbis T A.Seedling demography in an alpine ecosystem.American Journal of Botany,2003,90(8): 1197–1206. |