 2015, Vol. 35
2015, Vol. 35文章信息
- 刘辉, 姜广顺, 李惠
- LIU Hui, JIANG Guangshun, LI Hui
- 北方冬季有蹄类动物4种数量调查方法的比较
- A comparative study on four survey methods used in ungulate population size estimation in winter in North China
- 生态学报, 2015, 35(9): 3076-3086
- Acta Ecologica Sinica, 2015, 35(9): 3076-3086
- http://dx.doi.org/10.5846/stxb201308172102
-
文章历史
- 收稿日期:2013-08-17
- 修订日期:2014-08-01






2. 国家林业局猫科动物研究中心, 哈尔滨 150040
2. The Feline Research Center of State Forestry Administration, Harbin 150040, China
种群数量的有效评估是野生动物研究和管理的重要基础,而获得较准确的绝对种群密度进而得到种群数量,是非常复杂的,有时得到的结果还会引起较大的争议[1]。在开阔的栖息地最有效的方式莫过于直接进行实体的观察,动物实体可见度高,交通人力等保障充分,可搜索全部调查区域,直接得出某区域的目标物种种群的绝对数量[1]。如果以上条件得不到满足,如动物隐蔽性强或者栖息地异常复杂,不能直接进行密度的估计时,则研究者只能通过间接的痕迹,如动物的足迹链、粪便、脱落的毛发、采食痕迹等来估测动物的相对密度,再通过一定的修正系数,转换为目标物种种群的绝对数量[2]。
北方有蹄类动物调查往往根据动物在雪地上留下的痕迹来间接评估种群数量。随着统计方法和动物痕迹识别技术的发展,常用的调查方法也在不断变化。早期的野生动物研究方法主要包括粪堆统计法[3]、样线法[4]、样带法[5]、截线法[6]等。目前这些方法还在使用,同时引进了大样方法[7],非损伤性CMR法[8]。但使用较多的方法主要是样线法、样带法、大样方法和非损伤性CMR法。然而,各种方法基于的统计学假设和生态学原理不同,适用条件也存在很大的差别,因而用不同方法在同一调查地点调查同一目标种群所得到的调查结果往往存在较大的差异,但至今为止,没有发现就以上方法进行对比性研究,调查方法的选取主观性较强,所得结果的准确性也没有进行科学的评价。为此,设计了此次实验性的调查,于2012 年3 月在汗马自然保护区塔里亚河下游布设10 条长为3.8到4.2 km的样线,设计10个长为3.8到4.2 km,单侧宽为50 m的调查样带,并收集到68 份驼鹿粪便。2012 年12 月,在整个保护区布设11 个5 km ×2 km的大样方,其中2 个在样线和样带布设区域。我们分别采用样线法、样带法、大样方法和非损伤性CMR法调查评估了该保护区约120 km2区域内的驼鹿数量,并对各种方法的原理和适用条件进行了探讨,希望能就当前所使用的方法做出科学评价,探讨适合北方冬季有蹄类动物种群数量调查的方法。
1 研究区域内蒙古汗马国家级自然保护区(东经122°23′34″—122°52′46″,北纬51°20′02″—51°49′48″)位于大兴安岭山脉的西坡北部,东与黑龙江省呼中国家级自然保护区相接,南近呼伦贝尔大草原,行政隶属于内蒙古自治区根河市。保护区南北长约57 km,宽约33 km,总面积为107 348 ha,森林覆被率高达88.4%,该区域的森林类型主要有针叶林、针阔混交林、针灌混交林和阔灌混交林。平均海拔在1 000—1 300 m之间,山坡较缓,坡度一般在10°至20°。
保护区属寒温带大陆性气候,全年平均气温为-5.4 ℃,极端最高气温达到35.5 ℃,极端最低气温可达-49.5 ℃,保护区全年有10 个月的时间有积雪,最大积雪深度达到50 cm,年降水量可以达到450 mm左右,保护区内最大的河为塔里亚河,是黑龙江上游——额尔古纳河的主要支流激流河的发源地之一。保护区主要的树种包括:兴安落叶松、樟子松(Pinus sylvestris var. mongolica) 、云杉(Piceakor aiensis) 、白桦(Betula platyphylla Suk)、山杨(Populus davidiana)、甜杨(Populus suaveolens Fisch)和钻天柳(Chosenia arbutifolia (Pall.) A. Skv.)。由于保护区内从未进行过任何的采伐和经营活动,几乎没有任何人为干扰,该保护区仍保持了原始林状态。保护区内有种类繁多的兽类在此栖居,主要包括猞猁(Lynx lynx) 、紫貂(Martes zibellina)、棕熊(Ursus arctos collaris)、马鹿(Cervus elaphus) 、狍(Capreolus pygargus) 和雪兔(Lepustim idus) 等。由于栖息地环境较原始,较少的人为干扰和丰富的水源,该保护区驼鹿(Alces alces cameloides)资源较为丰富,适合本研究的开展。本项目4 种调查方法的比较试验,在汗马自然保护区塔里亚河下游约120 km2区域展开,该区域主要由针叶林和阔灌林组成。样线、样带、大样方以及粪便样品的分布见图 1。

|
| 图 1 样线法样线的分布(a),样带法样带的分布(b),大样方法大样方的分布(c)和非损伤性CMR法随机收集的粪便样品的分布(d) Fig.1 Distribution map of transects (a),belt transects (b),sample plots (c),and scat samples (d) in the study area |
使用样线法进行驼鹿种群数量调查,在试验区域内共布设10 条长约为4 km的样线,每天完成2 条,每条样线由两人负责调查,其中一人为经验丰富的保护区工作人员,掌握常见大型兽类雪地足迹识别技术。沿样线行进期间,只记录判断为新鲜(<24 h)的驼鹿足迹链数量。
样线法绝对密度计算使用俄罗斯远东地区有蹄类密度调查使用的FMP公式(Formozov-Malyshev-Pereleshin Formula)进行计算[1],即通过24 h内驼鹿通过样线留下的足迹链数目来推算驼鹿密度。
(1) FMP公式,其原理是根据分析动物的活动路径与样线相遇的概率来计算动物的密度:
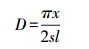 式中,x是总的新鲜足迹链数目,s是总样线长度,l是平均日活动距离。
式中,x是总的新鲜足迹链数目,s是总样线长度,l是平均日活动距离。
(2)总研究样线区域驼鹿的平均密度计算公式:
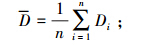 式中,G为所有调查样线驼鹿的平均密度;n为调查样线的数量;Di=第i 条样线驼鹿的分布密度。
式中,G为所有调查样线驼鹿的平均密度;n为调查样线的数量;Di=第i 条样线驼鹿的分布密度。
(3) 驼鹿分布密度的置信区间:
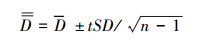 式中,
式中,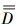 为驼鹿分布密度的置信区间(置信概率P=95%,自由度f = n-1,SD为标准差,t为Student′s分布表的值); n为调查样带总数。
为驼鹿分布密度的置信区间(置信概率P=95%,自由度f = n-1,SD为标准差,t为Student′s分布表的值); n为调查样带总数。
(4) 调查区域驼鹿数量:
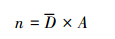 式中,A 为研究区域总面积, N 为研究区驼鹿数量。
式中,A 为研究区域总面积, N 为研究区驼鹿数量。
由于我国尚无驼鹿冬季日活动距离方面的研究,计算公式应用欧洲Neumann等的研究成果,即通过驼鹿冬季每小时的活动距离约为0.075 km来推算其日活动距离为1.8 km[9]。
2.2 样带调查法盛和林等指出,样线法是进行大范围内大型野生动物调查的有效方法[10]。根据1997年国家林业局《全国陆生野生动物资源调查与监测技术规程》,传统意义上的样线法在进行种群密度统计时,一般要求具有一定的样线可视宽度。因此,该方法实质上是样带法[11]。多年来,雪地样带法在我国冬季有蹄类调查中发挥了重要作用,利用该方法曾获得了野猪、马鹿、梅花鹿等多个北方有蹄类物种的密度和数量评估数据[12, 13, 14]。
2.2.1 样带布设该方法和样线法同时进行,即布设10 个长约4 km,单侧宽为50 m的样带,布设的样带间距大于动物的日活动距离,记录样带内48 h内的新鲜足迹链数量,同时记录样带内发现的粪便堆数、卧迹数目以及采食痕迹。
2.2.2 数据处理(1) 换算系数
在样带调查的过程中,若满足以下条件,则换算系数为1[15]。具体条件:1) 样带内新鲜单一足迹链可见。足迹链须是48 h内留下的,可根据雪的融化程度、色泽及被风侵蚀的情况进行判断;2) 若出现判断为群的足迹链,则沿足迹链的两个方向行走观察,直到群足迹链分开,统计驼鹿的实际足迹链数量,不同个体的足迹链可以根据雪地足迹的大小、步距大小及行进方向进行区分,并利用粪便和卧迹数量进行佐证[15]。
(2) 驼鹿密度计算公式
1 ) 样带内驼鹿密度计算公式[16]:
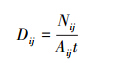 式中,Dij为i样地中第j样带驼鹿的种群密度; Nij为i 样地中第j样带2 d内的活动痕迹数; Aij为i 样地中第j 样带的面积; t为累计时间,t=2 d。
式中,Dij为i样地中第j样带驼鹿的种群密度; Nij为i 样地中第j样带2 d内的活动痕迹数; Aij为i 样地中第j 样带的面积; t为累计时间,t=2 d。
2) 样带内驼鹿的平均密度计算公式:
 式中,G= 所有调查样带中驼鹿的平均密度;n=调查样带的数量;Di=第i 个样带中驼鹿分布密度。
式中,G= 所有调查样带中驼鹿的平均密度;n=调查样带的数量;Di=第i 个样带中驼鹿分布密度。
3 ) 驼鹿分布密度的置信区间:
 式中,为驼鹿分布密度的置信区间(置信概率P=95%,自由度f=n -1,SD为标准差,t为Student′s 分布表的值); n为调查样带总数。
式中,为驼鹿分布密度的置信区间(置信概率P=95%,自由度f=n -1,SD为标准差,t为Student′s 分布表的值); n为调查样带总数。
4) 调查区域驼鹿数量:
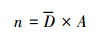 式中,A为研究区域总面积,N为研究区驼鹿数量。
式中,A为研究区域总面积,N为研究区驼鹿数量。
5 ) 调查精度:
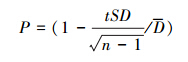
该方法基于进入和离开闭合多边形(样方)的差值来统计种群数量,样方的面积不小于北方有蹄类动物种群的日活动区域的最大面积。通过布设间距500 m长5 km的5 条平行样线,将调查区域分割成4 个小样方,调查人员记录每条样线上动物新鲜足迹链数量及动物的行进方向,动物24h内进入和离开样方的差值为24h内留在该样方内的个体数量,由此计算出该大样方内的目标物种实体数,结合样方面积得到样方内动物密度,并由此推断出栖息地类型分层的密度和总密度及种群数量(图 2)。

|
| 图 2 大样方法调查样方设计示意图 Fig.2 Plot design in the Sample Plot method |
鉴于本区域主要由针叶林、针阔林、针灌林和阔灌林组成,根据各林型所占的比例,分别将11个样方布设在针叶林(5 个)、针阔林(1个)、针灌林(2 个)和阔灌林(3 个),11个样方的总面积为110 km2,抽样面积占整个保护区面积的10.19%,达到大样方调查法规定的10%的最低标准。其中样线法和样带法以及非损伤性CMR 法取样的区域,即塔里亚河下游约120 km2区域内,共布设了2 个大样方,分别代表了该区域的两种主要林型,针叶林和针阔林。由保护区分布在不同林型内的11 个大样方得出各种林型内的驼鹿密度,根据该120 km2区域内针叶林和针阔林的面积,即可得到该区域内驼鹿的种群数量。每个样方由5 组调查人员一天内完成,每组按照方案设计要求步行,并记录沿途所观察到的驼鹿24 h内留下的新鲜足迹链数目及行进方向,遇到群链需要进行正向和反向跟踪,以确认群链的准确个体数量组成。每组队员须走完一条完整的长为5 km的样线及小样方“封口”,保证每个小样方及完整的大样方无缺口,形成一个闭合的多边形(图 2)。
2.3.2 样方内有蹄类数据记录和统计实线指示有蹄类动物个体足迹方向;虚线代表样线位置和布设方向;单箭头代表单个动物,双箭头及多箭头代表动物群组,箭头方向代表动物前进的方向 以图 2动物出现情况为例,说明大样方法种群数量统计方法。统计单个大样方内动物数量时,须首先把大样方分隔为4个小样方,依次编号为①、②、③、④。基于被调查动物24h内的新鲜足迹链的数量与方向,可以根据辐射状的调查足迹链确定每个多边形样方内的动物数量。根据小样方内是否有动物群组出现,计算时应分以下两种情况:
(1) 小样方内无群组出现(距离小于30 m的足迹链确定为1组),如图 2 中的小样方①和②所示,进入和离开小样方的动物均是以单只的形式活动,则各小样方内24 h留下的由新鲜足迹链数之和减去离开的足迹链数量之和,即为该小样方内该目标动物的实体数量。例如,小样方①中有2 只动物进入,2 只动物离开该样方,则认为该样方内动物数量为0只;同理,小样方2中有2 只动物进入,1 只动物离开,则认为有1 只动物扔留在该小样方内,该小样方内的动物数量为“1”。当记录到的进入小样方的足迹链数量小于离开的数量时,该小样方内动物数目记录为“0”。
(2) 小样方内有群组出现,如图 2 中的小样方③和④所示,进入和离开小样方的动物有些是以群组的方式存在的,基于24h内动物群的大小是不会变化的假设来判断群的存在和数量,统计此种情况下的动物数量,应首先判断某一群组是否离开该小样方,如小样方③内,样方的左侧进入了个体数为2的群组,但右侧发现有个体数为2的群组离开该样方,认为这两个体数为2的群是相同的群组,则该小样方内以群组形式出现的动物个体数为“0”。然而,有1 个以单只形式的动物进入该小样方,进入后没有离开,则认为该小样方内的动物数量为“1”;小样方④中,有一个个体数为3的群组进入该样方,然而并没有发现离开该小样方的个体数为3的群组,认为该群组仍留在该样方内,有1 只个体离开该小样方但未发现有单只的个体进入。因此,以单只形式计算的该小样方内的个体数量为“0”,两种形式出现的动物总个体数为“3”。因此,该大样方内的动物实体数量为4 个小样方内的个体数量之和“5”。
2.3.3 有蹄类种群数量和分布密度的统计由2.3.2可以得出某大样方内的个体数量,结合该大样方的面积,可以计算出该样方的动物密度。在根据分层抽样的原理,由各大样方内的动物密度,可得到该抽样层各个样方的平均密度,结合该抽样层的总面积,即可得出该层抽样的动物种群数量及置信区间。再由各抽样层的数量相加和,即可得到所有研究区域的目标物种种群数量。
(1)样方内某一有蹄类的密度:

式中,di为第i个样方内某一有蹄类的密度;ni为第i 个样方有蹄类评估数量;si为第i 个样方面积。
(2) 第j 层样方有蹄类的平均密度:

式中,Dj为第j 层样方平均密度;m 为 第j 层样方数量。
(3) 第j 层有蹄类分布密度的置信区间:

式中, j为有蹄类分布密度的置信区间(置信概率P= 80%,自由度f = n -1,σj 为标准差,t 为 Student′s 分布表的值)。
(4) 计算出各层某种有蹄类数量与估计区间:
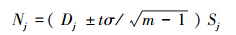
式中,Nj为第j 层有蹄类数量; Sj为第j 层面积。
(5) 计算出调查区域某种有蹄类总体密度与估计区间:
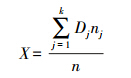
式中,X为总体密度均值; nj为第j 层的样本数; n 为调查样方总数量; k 为分层数。
式中,σ为为总体标准差,N为总体容量,总体密度置信区间X= X±ΔX。
(6)调查估计精度:

式中,n 为调查样方总数量;为调查样方中有蹄类的平均密度。
2.4 非损伤性CMR法统计有蹄类种群数量非损伤性分子技术用于有蹄类种群数量调查的方法主要有两种:非损伤性标志重捕法(CMR)和渐近线法。非损伤性CMR法同传统CMR法相比较,差别在于标志重捕的是动物的非损伤性DNA分子而不是动物个体本身[17]。相对于CMR法来说,渐近线法的统计学原理相对简单,在实际应用中实例较少,且均是配合CMR使用。因此,更多的研究还是使用非损伤性CMR法。
2.4.1 样品采集动物粪便是最易获得的非损伤性实验材料,加上我国北方冬季气温较低,采集到的粪便样品较易保存,因此以粪便DNA为基础的分析已经成为非损伤性种群数量调查的主流[18]。粪便样品的采集通过均匀布设样线来进行,本文还使用了跟踪驼鹿足迹链的方式来采集来自不同个体的粪便,并有意在相同区域的较近距离内采集疑似同一个体的粪便来检验个体鉴定的准确性。尽量挑选动物24 h内排出的埋于雪下的粪便样品,戴上一次性PE(Polyethylene)手套捡取数粒粪球至封口袋内,避光保存,尽快转移至实验室-80 ℃冰箱冷冻保存。
2.4.2 非损伤性样品DNA的提取和个体基因型的确定(1)DNA的提取
由于非损伤性样品DNA含量较少,因此,在DNA的提取及分型的过程中必须减少人为或者实验室环境的污染。粪便DNA的提取主要有两种方法:酚/氯仿法及相应的改进方法、DNA提取试剂盒法。近年来进口DNA提取试剂盒价格的下降及价格相对低廉的国产试剂盒的出现,使得从大批量粪便样品中提取DNA成为可能。
(2)微卫星座位的选取
目前的野生动物种群数量非损伤CMR研究中,高度多态性的微卫星遗传标记成为首选。基因座位的选取要避免因座位选取过少出现“影子效应”,同时也要防止因座位选取过多而导致实验成本的大幅增加及实验误差的增大和种群数量被高估。大多数的研究者利用PI(Probability of Identity)值来确定应选用的微卫星座位数。本文使用了12 对曾被用于驼鹿遗传多样性研究的多态性微卫星位点(RT1,RT24,RT30,BM203,NVHRT01,MCM58,MAF46,CSSM003,CSSM039,RT9,CP21,FCB193,MCM130)[19, 20]用于个体鉴定的研究。
(3) PCR扩增
要获得准确的个体基因型,必须避免分型错误。本实验使用的是Taberlet 等[21]提出的多管扩增PCR法(Multi-tube PCR)。基本流程是先进行3次独立的PCR,记录出现2 次以上的等位基因,对于3次都是纯合或者只出现1次杂合的DNA样品,再进行4 次PCR,总共7 次PCR中,记录出现2 次以上的等位基因。
使用QIAamp DNA Stool Mini Kit (Qiagen,Germany) 试剂盒对采集到的68 份驼鹿粪便样品进行DNA的提取,并使用1.0%的琼脂糖凝胶电泳检验DNA提取是否成功。使用日本东洋纺公司最新的高保真KOD FX NEO酶及操作要求进行12 个位点的PCR扩增,获得每个样品可靠的基因型,利用GIMLET[22]计算12 个基因座的联合P(ID)值。
2.4.3 个体鉴定及种群数量分析(1)个体鉴定
在获得了可靠的基因型后,利用Excel内嵌的MStools插件进行所有样本基因型的匹配。判断不同个体来自同一个体的原则是:1)所有位点上的基因型都相同;2)只有一个位点上的一个等位基因存在差别[23]。
(2)非损伤性CMR分析
在野外取样过程中,常常在不同的地方得到同一个体的粪样,根据CMR法的统计原理,把其中的任何一次记作标记,则其他的记录可视为对该个体的重捕[19]。很多软件可以用来计算此类抽样的种群数量,CAPWIRE[24]即是其中的一种,CAPWIRE软件因取样对象被捕捉概率不同分为两种模型:ECM(Even Capture Model)模型和TIRM(Two Innate Rates Model)。本文使用R软件中的CAPWIRE包中的[25]两种模型进行计算,使用10000次的bootstrap检验来产生两种模型下95%水平上的种群数量N的置信区间。并用该软件包的似然比测试(Likelihood Ratio Test)功能检验采集到的样品数量分布对于每种模型的适用性,根据所得到的P值若某模型检验结果P值显著偏离0,则说明样品数量分布符合该模型的假设。由此,确定适合的模型和并用该模型评估种群数量结果。
3 结果 3.1 样线法调查结果样线法调查区域主要林型为针叶林和阔灌林,共完成每条长为3.8到4.2 km的10 条样线的调查,样线间隔为3 km(图 1a)。统计10 条样线的长度和横穿每条样线上新鲜驼鹿足迹链数量,将每条样线代入驼鹿密度计算公式可得各样线的驼鹿密度,由各样线的密度求平均值,可得平均密度为 =1.433 只/km2,密度分布区间为D =(1.432±0.506) 只/km2。样线法总研究区域总面积为A=117.6 km2,则 该区域内驼鹿的总数量为n=(168±59) 只。调查精度P=75.35%。
3.2 样带法调查结果
共完成随机布设样带10 条,长3.8—4.2 km,单侧宽度确定为50 m的样带进行统计(图 1b)。该调查和2.1中样线调查同时进行,根据各样带内新鲜足迹链和粪便、卧迹等的信息综合确定各个不同样线长度的样带内目标物种的数量。再根据动物的活动时间t=2 d,各样带的面积A,计算可得各样带的平均密度为 = 2.022 只/km2,驼鹿的密度分布区间为=(2.02±0.62) 只/km2。研究区域总面积为A=117.6km2,则 该区域内驼鹿的总数量为n=(237±72) 只。调查精度P=69.13%。
2012年12月,在汗马保护区总面积为10800 km2范围内布设11个2 km ×5 km的大样方来调查该区域内的驼鹿数量(图 1c)。经过对该区域林相图进行矢量化,划出研究区域的边界和研究区域林型分布,由11 个样方得出的针叶林驼鹿密度为0.22(0.07—0.37) 只/km2,针阔林密度为0.6 只/km2,根据实验统计比较目标区域面积为120 km2的林型进行计算,其中针叶林的面积为91.5 km2,针阔林的面积为28.5 km2,计算出该区域内驼鹿的种群数量为37(23—50)只。
3.4 非损伤CMR法调查结果2012年3月,采用样线法和跟踪足迹链的方法,在调查区域共收集到新鲜驼鹿粪便68 份(图 1d)。从这68 份粪便样品中成功提取和扩增了67 份粪便样品的DNA。 12 对多态性微卫星引物(等位基因数最小为CP21,2 个等位基因;最大为RT24,14 个等位基因)的联合P(ID)值为1.57×10-14,即便是全同胞的个体判别率也达到了0.7 ×10-4,满足个体鉴定的要求。根据个体鉴定的原则,从成功提取和扩增的67 份粪便样品中鉴别出42 只驼鹿个体。使用R软件的CAPWIRE包,分别利用两种模型进模拟,使用ECM法得到的结果为55 只,97.5%的判别率下的置信区间为43—68。利用该软件包的似然比测试得到的P值显著偏离0,说明调查区域内的不同的个体具有相同的捕捉率。因此,汗马保护区120 km2的区域内估计分布有55 只驼鹿个体,97.5%的判别率下的置信区间为43—68 只。
3.5 4种调查方法结果的比较本次试验检测了北方有蹄类广泛应用的4 种调查方法,分别估计汗马自然保护区约120 km2区域内的驼鹿数量,比较结果见图 3。

|
| 图 3 4种方法得到的驼鹿种群数量及置信区间 Fig.3 Moose population size estimated with 4 methods and its confidence interval |
由图 3可知,使用不同的调查方法评估该区域内驼鹿的种群数量,得到的结果各不相同,样线法168(109—227) 只和样带法237(165—309)只都明显高于大样方法37(23—50)只和非损伤CMR法55(43—68) 只。其中,样带法所估测的种群密度最高,大样方法和非损伤CMR法得到的结果较为接近,置信区间有部分重叠,且总体估测值远远低于前两种方法。其中,基于大样方法的纯野外调查评估得到的结果最为保守,但其最接近于非损伤CMR法的结果。
4 讨论 4.1 4种方法原理和准确性评估样线(FMP)法仅基于雪地新鲜足迹链数量和样线的长度,综合动物的日活动距离计算动物密度。由于没有考虑更多个体信息,当某一个体或群反复穿过样线,在该样线上留下大量足迹链,伪重复可能会使估算结果偏高。另外,该方法是建立在已知目标动物平均日活动距离的基础上的。然而,不同季节及生境类型,以及动物的性别、年龄组的差异都会导致日活动距离的变化,想要获得准确的平均日活动距离需要做大量的研究试验,所获得结果在不同地区和气候环境下存在适用性的差异,导致调查结果可能存在较大的误差[1];样带法计算简单,统计样带内的新鲜足迹链数量,再利用粪便、卧迹等的信息加以佐证,得到该样带内的实体数量,结合样带的面积,即可得到样带内动物的密度。然而,由于该方法样带的宽度确定缺乏科学性、没有系统的统计标准且主观性较强,一旦样带内出现多条足迹链和多处动物痕迹,不能消除“伪重复”,动物的实体数量会达到很高的水平。另外,该方法难以进行修正,即使修正,转换系数的科学确定也是个难题。因此,在假设转换系数为1的情况下,得到的动物密度可能会达到相当高的水平,之前国内进行的多次有蹄类调查都出现了这样的情况,调查结果可信度不高;大样方法基于一个密闭的多边形,通过24h内进出该多边形的动物实体数量来统计样方内的动物密度。该方法利用分层取样的原理,考虑到动物群体的日活动区域范围,根据实际的生境类型分布信息设定大样方的数量和分布,较科学。另外,在统计样方内动物实体数量时,定义了动物的单只和群组两种情况,并根据动物的行进方向,利用现代化的GIS技术根据由GPS采集的地理信息和调查地区的林相图等,将调查结果可视化,能很好的反应动物在各个小样方及整个大样方内的分布,得到的结果更可靠[26];近年,基于有蹄类个体DNA信息的标志重捕技术,刚刚兴起有蹄类调查的非损伤CMR法。这种方法在不打扰动物种群的情况下获得非损伤遗传样品进行分析,虽然粪便等非损伤性样品DNA往往存在一定的降解,且DNA质量较差,会对实验结果造成影响,但扩大研究区域和增大抽样量能很好的弥补实验误差带来的缺陷[18]。和传统CMR相比较,基于不易发生改变的个体遗传物质的标识信息更易采集,并且更加准确。另外,使用的CAPWIRE 软件包根据不同个体粪便样品被捕捉到的概率的不同,分为ECM和TIRM两种模型,能得到较好的种群数量和估测区间,该方法的准确性已得到多次的验证[8, 18, 25]。
4.2 4种方法可行性比较研究首先应当阐明的是,本文只是以汗马保护区驼鹿调查为试验案例来说明4 种方法的实用性。当然,这几种方法均可用于野猪,梅花鹿,马鹿,狍等其他北方有蹄类动物的调查研究。通过调查结果可知,本文所使用的4 种方法得到了4 个不同的调查结果,但本文只是一个试验性的调查,对于一般的有蹄类调查,需要根据调查结果要求的准确性,调查的费用支持,调查的可操作性以及对野外动物足迹等识别技术的需求等方面来选择适用的调查方法,需要从以下方面来综合衡量:
(1)准确性
据4.1所述,在4 种方法中,非损伤性CMR法或者大样方法所基于的原理和统计方法以及所获得的结果更为科学,且在北方的有蹄类研究中近年也都进行了很好的尝试和应用。例如,田新民等使用非损伤CMR法调查了数量稀少的完达山林区马鹿的种群数量[8]。张常智等使用大样方法估测东北虎分布区猎物的密度[7]。传统使用的样带法和样线FMP法的调查结果准确性低于大样方法和非损伤CMR法。但是,对于样线法和样带法调查结果,可以通过多年调查相对数量(如,足迹链数量/km)的变化揭示种群资源的相对变化趋势。
(2)费用支持
进行大规模的有蹄类调查需要耗费大量的人力物力和财力。因此,可根据不同方法需要的费用来选择合适的方法。很显然,样线FMP法只需要沿既定的样线,收集动物的新鲜足迹链信息,此方法较为快捷,花费最少。而样带法在记录动物足迹链信息的同时,还要收集两侧样带内的辅助性信息,如粪便,食痕等,花费时间和费用将大于样线法。大样方法一般要求每个样方在24h内完成,且调查样线间距一般小于500 m,这样就需要多组调查队伍同时开展调查,要兼顾目标物种不同生境类型都能被取样,且要保证10%—30% 的抽样率,有时样方的数目会比较大,花费会稍高于以上2 种方法。然而,上述3 种方法中大样方法获取的数据最为科学,近50 多年来一直是俄罗斯狩猎管理部门对狩猎场有蹄类动物种群资源精细管理的主要调查方法。这3 种方法都是纯粹的野外调查统计工作,花费也仅限于野外工作,与此相比,非损伤性CMR法尽管更为科学,不但需要野外遗传取样的大量工作,还要对所采集到的样品进行DNA分析,微观实验的花费往往会数倍于野外工作,一旦采集的样品质量不高,实验结果一致性不高,需要进行多次的重复性实验,费用会大大增加。
(3)可操作性
根据调查地区和物种的不同,必须考虑到不同方法的可操作性。首先是调查人员的专业素质,样线法对调查人员的专业性要求最低,但必须准确的识别目标物种的足迹链种类和数量。样线法进行统计计算的前提是知道动物的日活动距离,如前文所述,动物在不同生境类型和环境下的日活动距离会有较大的差异,如何设计试验,利用无线电项圈等技术或进行足迹链的跟踪获得准确的日活动距离,是使用该方法的前提。样带法同样要求调查者具备专业的动物足迹链和痕迹识别技术,以及综合辅助信息准确判断不同足迹链来源实体数量的技术。大样方法要求调查人员能熟练掌握动物足迹识别技术,并能辨别出“群组”及个体数量,经过专业的培训,地方性的林业部门和保护区都可以使用该方法。另外,由于只要保证10%以上面积的抽样即可代表调查区域,该法相对比较节省时间,但必须指出的是该方法只适用于数量较为丰富的有蹄类的数量调查。非损伤性CMR法的样品较易采集,粪便的保存等也相对比较简单。但就可操作性来说,该法应用起来难度最大,首先,它是建立在外业的基础上的,对样品的采集要求样本量和抽样区域要尽可能大,最好是地毯式搜索[18]。其次,该方法又建立在现代分子生物统计学的基础上的,分子生物学实验室和相关的实验材料和方法是必不可少的,对人员分子生物学技术素质和设备的要求很高,一般性的调查通常很难满足这样的条件,需要和高等院校或者研究所进行合作。再次,由于是分阶段式的工作模式,同以上方法相比较,该方法较为费时。
5 结论(1)北方冬季有蹄类调查应该根据调查对象和目的,从现有方法中筛选适合调查区域和目标物种的调查方法。对于一般性大规模的调查,仅获得相对数量和分布的动态变化,且对调查结果精度要求不高,不需要较准确的绝对数量评估时,可采用常规的样线法或样带法,但如计划获取绝对数量,建议使用试验性研究探索科学确定修正系数的方法,使结果更准确。
(2)对于有蹄类动物的精细管理时(如狩猎场有蹄类动物的年度数量动态监测、东北虎分布区猎物的恢复等),推荐选择大样方法。就野外调查方法来说,该方法所基于的科学假设最为合理,只需对基层调查队伍进行适当的技术培训,地方林业部门和保护区即可独立使用该方法,从而获得较为准确数量评估结果。而非损伤性CMR法调查结果的准确度虽最高,但获取相应结果所需要的花费时间搜寻野外遗传样本和大量的实验室分析经费,投入显然高于纯粹的野外调查方法,适用于小面积和特别濒危物种的种群数量调查和研究。
致谢: 内蒙古汗马国家级自然保护区同仁帮助野外调查,张雪同学帮助提取DNA和实验,中国科学院动物研究所胡义波博士帮助工作,特此致谢。
| [1] | Stephens P A, Zaumyslova O Y, Miquelle D G, Myslenkov A I, Hayward G D. Estimating population density from indirect sign: track counts and the Formozov-Malyshev-Pereleshin formula. Animal Conservation, 2006, 9(3): 339-348. |
| [2] | Wilson G J, Delahay R J. A review of methods to estimate the abundance of terrestrial carnivores using field signs and observation. Wildlife Resources, 2001, 28(2): 151-164. |
| [3] | 马建章, 常弘, 孟宪林. 马鹿粪堆分布型的研究及其应用. 野生动物, 1989, (6): 6-12. |
| [4] | 朴仁珠, 关国生, 张明海. 中国驼鹿种群数量及分布现状的研究. 兽类学报, 1995, 15(1): 11-16. |
| [5] | Harris R B, 刘永生, 蔡桂全, O'Gara B. 使用样线法进行马麝粪堆密度的测量. 兽类学报, 1992, 12(4): 302-305, 293-293. |
| [6] | 朴仁珠. 截线法对西藏盘羊种群数量的估计. 生态学报, 1996, 16(3): 295-301. |
| [7] | 张常智, 张明海. 黑龙江省东完达山地区东北虎猎物种群现状及动态趋势, 生态学报, 2011, 31(21): 6481-6487. |
| [8] | 田新民, 张明海. 基于粪便 DNA 的马鹿种群数量和性比. 生态学报, 2010, 30(22): 6249-6254. |
| [9] | Neumann W, Ericsson G, Dettki H, Radeloff V C. Behavioural response to infrastructure of wildlife adapted to natural disturbances. Landscape and Urban Planning, 2013, 114: 9-27. |
| [10] | 盛和林, 徐宏发. 哺乳动物野外研究方法. 北京: 中国林业出版社, 1992: 161-169. |
| [11] | 高中信, 陈华豪, 陈化鹏, 罗理扬, 刘景全, 金崑. 动物生态学试验与实习方法. 哈尔滨: 东北林业大学出版社, 1991: 82-84. |
| [12] | 孟根同, 张明海, 周绍春. 黑龙江凤凰山国家级自然保护区野猪冬季容纳量及最适种群密度. 生态学报, 2013, 33(3): 957-963. |
| [13] | 周绍春. 生境边缘效应对完达山林区马鹿种群数量分布的影响 [D]. 哈尔滨: 东北林业大学, 2005. |
| [14] | 于江傲, 鲁庆彬, 刘长国, 周圻, 章叔岩. 清凉峰自然保护区华南梅花鹿种群数量与分布研究. 浙江林业科技, 2006, 26(5): 1-4. |
| [15] | 朱洪强, 葛志勇, 刘庚, 常素慧, 吴景才, 石晓军, 徐吉凤, 毛之夏. 吉林省长白山区野猪种群资源现状调查. 氨基酸和生物资源, 2011, 33(4): 13-16. |
| [16] | 郑祥, 鲍毅新, 葛宝明, 周元庆, 郑英茂, 华文礼. 九龙山自然保护区黑麂的种群密度, 分布与保护. 浙江师范大学学报: 自然科学版, 2005, 28(3): 313-318. |
| [17] | Taberlet, P, Waits L P, Luikart G. Noninvasive genetic sampling: look before you leap. Trends in Ecology and Evolution, 1999, 14(8): 323-327. |
| [18] | 詹祥江. 利用非损伤性遗传取样研究大熊猫的种群数量和扩散模式 [D]. 北京: 中国科学院研究生院(中国科学院动物研究所), 2006. |
| [19] | Wilson G A, Strobeck C, Wu L, Coffin J W. Characterization of microsatellite loci in caribou Rangifer tarandus, and their use in other artiodactyls. Molecular Ecology, 1997, 6(7): 697-699. |
| [20] | Røed K H, Midthjell L. Microsatellites in reindeer, Rangifer tarandus, and their use in other cervids. Molecular Ecology, 1998, 7(12): 1773-1776. |
| [21] | Taberlet P, Griffin S, Goossens B, Questiau S, Manceau V, Escaravage N, Bouvet J. Reliable genotyping of samples with very low DNA quantities using PCR. Nucleic Acids Research, 1996, 24(16): 3189-3194. |
| [22] | Valière N, Berthier P, Mouchiroud D, Pontier D. Gemini: software for testing the effects of genotyping errors and multitubes approach for individual identification. Molecular Ecology Notes, 2002, 2(1): 83-86. |
| [23] | Bellemain E V A, Swenson J E, Tallmon D, Brunberg S, Taberlet P. Estimating population size of elusive animals with DNA from hunter-collected feces: four methods for brown bears.. Conservation Biology, 2005, 19(1): 150-161. |
| [24] | Miller C R, Joyce P, Waits L P. A new method for estimating the size of small populations from genetic mark-recapture data. Molecular Ecology, 2005, 14(7): 1991-2005. |
| [25] | Pennell M W, Stansbury C R, Waits L P, Miller C R. Capwire: a R package for estimating population census size from non-invasive genetic sampling. Molecular Ecology Resources, 2013, 13(1): 154-157. |
| [26] | 李冰. 珲春自然保护区东北虎及猎物种群现状及保护研究 [D]. 上海: 华东师范大学, 2010. |