 2015, Vol. 35
2015, Vol. 35文章信息
- 陈斯养, 靳宝
- CHEN Siyang, JIN Bao
- 一类具分段常数变量的捕食-食饵系统的Neimark-Sacker分支
- Neimark-Sacker bifurcation behavior of predator-prey system with piecewise constant arguments
- 生态学报, 2015, 35(7): 2339-2348
- Acta Ecologica Sinica, 2015, 35(7): 2339-2348
- http://dx.doi.org/10.5846/stxb201306051340
-
文章历史
- 收稿日期:2013-06-05
- 网络出版日期:2014-05-08






种群生态学是迄今数学在生态学中应用最为广泛、发展最为成熟的生态学的分支。捕食-食饵系统是种群生态学中生物种群相互之间的基本关系之一,是构成复杂食物链、食物网和生物化学网络结构的基石,从而引起了广大数学工作者和生物学家的关注。祁君和苏志勇[1]在经典的捕食-食饵系统中考虑到由于捕食效应对食饵种群带来的正向调节作用后,提出了具有捕食正效应的捕食-食饵系统。从理论上说明了正向调节作用对系统的影响,并就第一象限内平衡点存在时的相图解释了捕食正效应的作用。杨立和李维德[2]利用概率元胞自动机模型对空间隐式的、食饵具Allee 效应的一类捕食-食饵模型进行模拟,发现随着相关参数的变化,种群的空间扩散前沿由连续的扩散波逐渐转变为一种相互隔离的斑块向外扩散。Freedman 与 Wolkowicz 在Rosenzweig-MacArthur模型[3]中选取第4功能反应函数进行了全局范围内的分支情况的研究。经典的捕食-食饵模型可以被表达成如下的非线性微分方程模型:

该类模型稳定性、Hopf分支、极限环等问题被广泛的给予研究[4, 5, 6, 7, 8, 9, 10, 11, 12],式中x(t),y(t),分别表示t时刻捕食者和食饵种群的数量,s表示捕食者的自然死亡率,f(x)表示食饵在无捕食者时的相对增长率,捕食者单位时间内捕获食饵的数量用功能反应函数g(x,y)表示。实际上,由于受到气候、周围环境和捕食者所固有的特性等因素影响,捕食者对食饵的捕获只在一定时间段或整数时刻并且对食饵的捕获具有滞后效应,故可选择更加符合实际的具分段常数变量功能反应函数: ,h(x,y)=b1x([t])-b2y(t)本文考虑食饵在无捕食者时按通常的Logistic方式增长,f(x)=r(1-a1x(t)),则模型可描述为:
,h(x,y)=b1x([t])-b2y(t)本文考虑食饵在无捕食者时按通常的Logistic方式增长,f(x)=r(1-a1x(t)),则模型可描述为:

模型(1)满足初始条件:
式中,r表示食饵的内禀增长率,a1表示食饵的环境容纳量,a2表示捕食系数,b1表示捕食效率常数,[t]表示对变量t∈[0,+∞)取整。 1 正平衡态稳定性分析
由模型(1)可知b1>a1s时,模型(1)存在惟一的正平衡态:

定理1 模型(1)满足初始条件(2)的解为正、全局存在且有界(∀t≥0)。
说明:对定理1运用反证法和比较原理即可得证,故将其证明略去。
当n≤t<n+1时,模型(1)为:

对(3)由n到t积分并令t→n+1,即得:

其中:h=r[1-a2y(n-1)],q=-s+b1x(n),对(4)在平衡态E(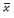 ,
, )处Taylor展开,令:
)处Taylor展开,令:

得(4)式的线性近似系统

其中 υ(n)=(ψ(n),φ(n))T

则线性系统(5)的特征方程为

其中 
以下应用Schur-Cohn判据[13]对模型(1)正平衡态稳定性进行分析,给出捕食者和食饵共存且数量保持稳定的条件。
定理2模型(1)满足下列5种情况之一:
(1) 当M3>1,M4<0,Δ>0时,
(2)当M3=1时,P5<P<1;
(3)当M3<1且Δ<0或M4<0,M5<0,Δ>0或M4>0,M5<0,Δ>0, <1时,0<P<1;
<1时,0<P<1;
(4)当M3<1,M4>0,M5<0,Δ>0,>1时,0<P<P2或 P1<P<1;
(5)当M3<1,M5>0,Δ>0时,P1<P<1;
则正平衡态E(,)局部渐近稳定。
其中: 
证明 由P=e-ra1x知0<P<1,根据文献[13],模型(1)满足如下3个条件:
(Η1)|ν 1+ν3|<ν2+1;(Η2)|ν3|<1;(Η3)|ν2-ν1ν3|<|1-ν32|
则正平衡态E(,)局部渐近稳定。
首先考虑条件(Η1)  且
且 等价于如下条件(i)或(ii):
等价于如下条件(i)或(ii):
(i)当M2<0时,0<P<1;(ii)当M2>0时, <P<1。
<P<1。
其次考虑条件(Η2) 由ν3>0知 等价于如下条件(iii)或(iv):(iii)当M3≤1时,0<P<1;(iv)当M3>1时,
等价于如下条件(iii)或(iv):(iii)当M3≤1时,0<P<1;(iv)当M3>1时, <P<1。
<P<1。
最后考虑条件(Η3) 由ν1<0,ν2>0,ν3>0知(Η2)和 等价于如下条件(v)或(vi):
等价于如下条件(v)或(vi):
(v) 当M3>1时,知M5>0,若Δ<0或Δ>0,M4>0(此时Pi<0(i=1,2)),则其交集为空集;若Δ>0,M4<0,知 Pi>0(i=1,2),P1<P<P2,设函数
f(x)=(M32-M3)P2+M4P+M5,由f(1)=M32-M3+M4+M5=e-b2y-1<0知P1<1<P2,故P1<P<1
(vi)当M3=1时,知M2=-e-b2y,M4=-1,M5=e-b2y,则M5<P。
(vii) 当M3<1时,知M32-M3<0,若Δ<0时,0<P<1;若Δ>0,M4<0,M5<0,则当Pi<0(i=1,2)时,0<P<1;若Δ>0,M4>0,M5>0或Δ>0,M4<0,M5>0时,知P1>0,P2<0,由f(1)<0得P1<1,则P1<P<1;若Δ>0,M4>0,M5<0,
由f(1)<0知1<P2<P1或0<P2<P1<1,若<1,
则f′(1)=-(M3-e-b2y+e-b2yM3)=-[2M3+(1-e-b2y)-(1-e-b2y)M3-1]=-[-1]>0,由此知1<P2<P1,故0<P<1.若>1,则f′(1)<0,
由此知0<P2<P1<1,故0<P<P2或P1<P<1。综合上述讨论与计算,由(i)、(iv)、(v)或(ii)、(iv)、(v)取交集可得定理中(1)的结论,由(i)、(iii)、(vi)取交集可得定理中(2)的结论,由(i)、(iii)、(vii)取交集可得定理中(3)、(4)、(5)的结论,证毕。
2 Neimark-Sacker分支分析本节以r作为分支参数,分别讨论模型(1)的Neimark-sacker分支存在性及其分支方向与稳定性。因情况(2)不会产生分支(分支临界值r0趋于零或无穷大),故下文对定理2中(1)的情况给出产生分支的条件,情况(3)的分支条件可同理给出。
定理3 设(1)中的参数满足M3>1,M4<0,Δ>0,则当max(P1,)<P<1时,(1)的正平衡态E(,)局部渐近稳定;当P=max(P1,)时,(6)存在一对位于单位圆环上共轭的特征根,且不存在λ=±1的根;当P>max(P1,)时,(1)的正平衡态E(,)不稳定,则(1)产生Neimark-sacker分支。
证明:由定理2可知,当M3>1,M4<0,Δ>0时,若max(P1,)<P<1,则(1)正平衡态E(,)局部渐近稳定;当P=max(P1,)时,假设λ1,2=e±iθ1是特征方程(6)的一对共轭纯虚根,则有
 求解得
求解得
由定理2中对(Η1)分析和<知,(6)不存在λ=±1的根,证毕。
下面讨论模型(1)的分支方向及其稳定性. 将(4)式写作如下变换形式:

其中,h1=r[1-a2x3],q1=-s+b1x1,(7)式在平衡态E(,,)的临界Jacobi矩阵:

由定理3可知Ζ0存在一对共轭纯虚根 λ1,2=e±iθ0, ,
, 。矩阵Ζ0和Ζ0T满足:Ζ0q=eiθ0q,Ζ0Tp=e-iθ0p,且其特征向量分别为:
。矩阵Ζ0和Ζ0T满足:Ζ0q=eiθ0q,Ζ0Tp=e-iθ0p,且其特征向量分别为:

取 
则 ,其中,
,其中, ,
,  i为qi(i=1,2,3)的共轭复数。将(7)式写为如下形式:
i为qi(i=1,2,3)的共轭复数。将(7)式写为如下形式:

其中F(x)=O(‖x‖2)是光滑函数且在平衡态E(,,)的Taylor展开式为:

B(x,y)和C(x,y,z)分量分别为:

当eikθ0≠1(k=1,2,3,4)时,映射(7)经坐标变换可化成:η→eiθ0η(1+ζ|η|2)+O(|η|4),其中ζ决定闭不变曲线的分支方向,可由下面公式计算:

记 
经计算可知:


将q,代入(8)和(9)式可得:
B(q,q)=(Ω1,Ω2,0)T,B(q,)=(Ω3,Ω4,0)T,C(q,q,)=(Ω5,Ω6,0)T
其中: 

经计算:

其中:
其中:

经计算可知:

其中 为κ的共轭复数。
为κ的共轭复数。
由以上计算知ζ的表达式如下:
由如上分析和推理可得如下定理4。
定理4[14]当 时(P由定理3确定),模型(1)在正平衡态E(,)产生Neimark-sacker分支;若ζ<0(>0),则模型(1)从正平衡态E(,)分支出惟一(不)稳定的超(亚)临界Neimark-sacker分支。
时(P由定理3确定),模型(1)在正平衡态E(,)产生Neimark-sacker分支;若ζ<0(>0),则模型(1)从正平衡态E(,)分支出惟一(不)稳定的超(亚)临界Neimark-sacker分支。
本节将通过实例,运用Matlab软件绘出相应的分支图,验证以上理论的可行性,并通过图形说明该模型复杂的动力学行为。
例 在模型(1)中,取a1=0.5,a2=0.4,b1=4,b2=2.5,s=0.1计算可得:
M3=1.2400,M4=-1.8426,Δ=2.7092,ζ=-0.4851<0,Ρ1=0.3304>(M3-1)/M3=0.1935
则分支参数的临界值r0=2.4852,惟一正平衡态E(0.8912,1.3860)
对应分支图为图 1。由图 1可知,当r<r0=2.4852,正平衡态E局部渐近稳定(图 2,图 3);当r=r0=2.4852时,产生N-S分支(图 4)及像平面图和解图(图 5);当r>r0=2.4852时,模型(1)出现复杂的动力学行为(图 1)。

|
| 图1 r-x,r-y分支图 Fig.1 r-x,r-ybifurcation map |

|
| 图2 x,y稳定解图(r=2.26<r0) Fig.2 stability solution map of x and y(r=2.26<r0) |

|
| 图3 稳定图(r=2.26<r0) Fig.3 stability map(r=2.26<r0) |

|
| 图4 N-S分支图 Fig.4 N-S bifurcation map |

|
| 图5 x,y像平面图和空间解图(r=r0=2.4852) Fig.5 phase plane and space solution map of x and y(r=r0=2.4852) |
本文应用Schur-Cohn判据、分支理论及中心流形投影等理论给出了具有时滞与分段常数变量捕食-食饵模型的稳定性及Neimark-sacker分支的存在性以及稳定性条件。通过模型的分析得到如下两个主要结论:
H1) 在捕食-食饵系统中考虑捕食者只在一定时间段或整数时刻且具有滞后效应捕食时,由定理2可知,系统的稳定性(捕食者和食饵共存且数量保持稳定)将会变得非常复杂。
H2) 由实例可知,系统在其它参数不变的情况下,当食饵的内禀增长率r<2.4852时,由图 1—图 3可知捕食者和食饵的数量处于稳定状态;当r=2.4852,由图 4,图 5知捕食者和食饵的数量将呈现周期性变化,系统产生Neimark-sacker分支;当r>2.4852时,由图 1知系统的正平衡态由稳定到不稳定。
综上所述,在捕食-食饵系统中,若考虑捕食者只在一定时间段或整数时刻且具有滞后效应捕食时,模型动力学行为将变得更为错综复杂;食饵的内禀增长率达到确定的临界值时,种群数量将失去原有的稳定性,模型将产生惟一稳定的超临界Neimark-Sacker分支。
| [1] | 祁君, 苏志勇. 具有捕食正效应的捕食-食饵系统. 生态学报, 2011, 31(24): 7471-7478. |
| [2] | 杨立, 李维德. 利用元胞自动机研究一类捕食食饵模型中的斑块扩散现象. 生态学报, 2012, 32(6): 1773-1782. |
| [3] | Kot M. Elements of Mathematical Ecology. London: The Cambridge University Press, 2001: 107-160. |
| [4] | He Z M, Lai X. Bifurcation and chaotic behavior of a discrete-time predator-prey system. Nonlinear Analysis: Real world Applications, 2011, 12(1): 403-417. |
| [5] | Wang W M, Ling L. Stability and Hopf bifurcation analysis of a delayed predator-prey model with constant rate harvesting. Journal of Mathematical Biology, 2009, 24(4): 1-14. |
| [6] | Wang J F, Shi J P, Wei J J. Predator-prey system with strong Allee effect in prey. Journal of Mathematical Biology, 2011, 62(3): 291-331. |
| [7] | Beretta E, Kuang Y. Global analyses in some delayed ratio-dependent predator-prey systems. Nonlinear Analysis: Theory, Methods & Applications, 1988, 32(3): 381-408. |
| [8] | Skalski G T, Gilliam J F. Functional responses with predator interference: viable alternatives to the Holling type II model. Ecology, 2001, 82(11): 3083-3092. |
| [9] | Mischaikow K, Wolkowicz G. A predator-prey system involving group defense: a connection matrix approach. Nonlinear Analysis: Theory, Methods & Applications, 1990, 14(11): 955-969. |
| [10] | Jost C, Arditi R. From pattern to process: identifying predator-prey models from time-series data. Population Ecology, 2001, 43(3): 229-243. |
| [11] |
Salt G W. Predator and prey densities as controls of the rate of capture by the predator |
| [12] | Martin A, Ruan S G. Predator-prey models with delay and prey harvesting. Journal of Mathematical Biology, 2001, 43(3): 247-267. |
| [13] | Jury E I. Theory and Application of the Z-transform Method. New York: Wiley, 1964. |
| [14] | Kuznetsov Y A. Elements of Applied Bifurcation Theory. New York: Springer-Verlag, 2004. |