 2015, Vol. 35
2015, Vol. 35文章信息
- 胡晓静, 张文辉, 何景峰, 尹艺凝
- HU Xiaojing, ZHANG Wenhui, HE Jingfeng, YIN Yining
- 不同生境栓皮栎天然更新幼苗植冠构型分析
- Architectural analysis of crown geometry of Quercus variablis BL. natural regenerative seedlings in different habitats
- 生态学报, 2015, 35(3): 788-795
- Acta Ecologica Sinica, 2015, 35(3): 788-795
- http://dx.doi.org/10.5846/stxb201304110676
-
文章历史
- 收稿日期:2013-04-11
- 网络出版日期:2014-04-03






2. 石河子大学农学院, 石河子 832000
2. Agriculture College, Shihezi University, Shihezi 832000, China
自然界绝大多数植物都是构件的集合体,每种植物的个体都有一个精确的并受遗传因素决定的“生长方案”,植物在任何一个发育阶段,其内部生长方案的表达形式都可用构型来表示,因此构型是植物内在遗传信息在一定时间表现的外部形态特征[1, 2]。基因是决定形态特征的内在因素[3];环境条件和各种生态因子影响着植物次生形态和构型;森林经营措施通过改善林地生境条件,改变树木生存空间,使林木向着理想的构型发育。林木的幼年阶段形态可塑性大,在不同生境条件下表现出较高的形态多样性[4]。研究生境因素对植物幼苗各构件(枝、叶、芽、根等)形态产生的影响以及构型差异对森林结构优化以及经营措施制定具有重要的参考意义[5]。
栓皮栎(Quercus variabilis)为壳斗科(Fagaceae)栎属落叶乔木,是重要的用材、软木、栲胶和能源树种。栓皮栎速生、耐旱、耐瘠薄,分布广泛,是我国暖温带落叶阔叶林和亚热带常绿落叶阔叶混交林的重要建群种[6]。国外没有栓皮栎,但对于栎属其它树种构型的研究开展较早。Oohata和Shidei研究发现在不同光强和密度条件下,7年生的Quercus philler分枝无显著改变[7]。McMahon和Kronauer对红栎和Quercus alba的研究结果[8]也与之类似。但Pickett和Kempf发现红栎个体内枝条和叶的排列方式都有不同程度的改变[9]。国内对于栎属树种的构型研究开始于20世纪90年代,且主要集中于辽东栎上[10, 11, 12]。对栓皮栎的研究主要是种群生殖动态、种群年龄结构和生物学生态学特性[13, 14, 15],以及不同经营措施促进林木生长和改善林地生境的内容[16],但有关栓皮栎天然幼苗构型分析的研究还未见报道。马莉薇等[17]对秦岭北坡栓皮栎实生苗的研究结果表明,林隙、林缘、林冠下由于资源的再分配和微生境的异质性,对幼苗的存活、生长产生显著影响。微生境差异是否也会对栓皮栎天然更新幼苗的构型产生影响?本研究以不同生境中(林冠下、林隙、林缘)栓皮栎天然幼苗为研究对象,通过统计栓皮栎幼苗的侧枝、叶片以及树冠的特征,分析不同生境中栓皮栎幼苗构型变化,阐明栓皮栎幼苗构型对生境因素的适应策略,探讨构型与生境关系,为森林结构及其经营措施优化提供依据。
1 研究地区与研究方法 1.1 研究地区自然概况研究区域位于秦岭南坡的山阳县(33°09′—33°42′N,109°32′—110°29′E),属北亚热带向暖温带过渡的季风性半湿润山地气候,年平均气温13.1℃,年平均无霜期为207d,年平均日照2133.8h,日照率48%,年平均降水量709.3mm。栓皮栎林是当地主要森林类型,主要分布在海拔800—1500m,乔木层郁闭度0.7—0.8,伴生树种主要有麻栎(Quercus acutissima)、槲栎(Quercus aliena)等,高度为7—12m;林下灌木盖度为0.2—0.4,高度为0.5—1.5m,主要有胡枝子(Lespedeza bicolor)、黄栌(Cotinus coggygria)、杜梨(Pyrus betulaefolia )等;草本层盖度为0.2—0.4,高为10—40cm,以百合科的麦冬(Ophiopogon Japonicus)和莎草科的苔草属(Carex)植物为主。研究区的林分近年来没有进行过较大规模的采伐或破坏活动,林相发育良好。本研究以阳坡的栓皮栎林为研究对象。
栓皮栎天然更新幼苗的生长环境分为林冠下、林隙以及林缘处,分别在这3个生境设置样地。选择林相相对整齐,林木分布均匀的地段,设置3块20m×20m林冠下生境样地(样地中心距林缘>40m)。在林内寻找林隙,林隙面积60—100m2,共3个林隙样地。在栓皮栎林边缘,即森林与农田的交汇处,设置3个10 m×50 m的样地。
1.2 研究方法 1.2.1 调查分析方法研究所用的栓皮栎幼苗为高度h<1.5m的天然更新苗,在每个生境(林冠下、林隙和林缘)样地中抽取20株幼苗作为研究对象。采用典型抽样法[18](排除1—2年生幼苗),在选定更新苗后,先测定树高、地径(或胸径)、冠幅、冠长,用半圆仪测量分枝角,测定各侧枝在树干上的位置(距主干顶芽的距离),统计各级枝条上的现存叶数,并计算树冠率,树冠率是指冠长占树高的百分比。为了确定不同树冠层次中侧枝和叶片分布格局,按照每株幼苗的树高分成4个层次,从冠顶起将树高分为:1H—3/4H、3/4H—1/2H、1/2H—1/4H、1/4H—0H(H为树高)4个层次,分别以 Ⅰ、Ⅱ、Ⅲ、Ⅳ标记。以每个1级侧枝在主干上的位置为准,按侧枝所处的高度确定所属的冠层。按下列标准确定各指标:1级侧枝是着生在主干上的枝条,2级侧枝则是着生在1级侧枝上的枝条,3级侧枝是着生在2级侧枝上的枝条,以此类推;分枝角度是指1级侧枝与树干方向的夹角;数量叶密度是指单位长度1级侧枝上所着生的叶片数量;面积叶密度是指单位长度1级侧枝上所着生叶片的面积;1级侧枝密度是指单位长度树干上着生的1级侧枝数量;2级侧枝密度则是单位长度1级侧枝上的2级侧枝数量[18, 19]。树干上发生侧枝的分枝概率和平均分枝数则根据一定时期内树干上每年发生的侧枝(或者从一级侧枝上发生的二级枝条)的调查结果,利用统计分析方法分别按照下式计算[20]:

式中,i为分枝数;Pi是分枝数为i的分枝概率;E(x)是平均分枝数。
1.2.2 实生苗年龄确定方法1—4a 实生苗个体年龄通过主茎上的芽鳞痕、茎干颜色确定,4a 以上实生苗个体的年龄通过观察主茎上的芽鳞痕,同时用放大镜观察基部年轮数确定[15]。
1.2.3 叶片特征测定方法叶片带回实验室后,先用EPSON PERFECTION 4870 扫描仪将叶片扫描成图片文件,用WinFOLIA 2004a 软件分析叶面积(LA)[20],然后将叶片放入80℃烘箱下烘48h至恒重,再称重,即为叶干重。比叶面积(SLA)为叶面积和叶干重的比值;单株的叶面积指数为:
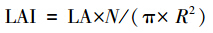
式中,LAI 为叶面积指数;LA为平均单叶面积;N为单株实生苗总叶片数;R为实生苗冠幅的半径。
1.2.4 数据处理采用EXCEL和SPSS 16.0进行数据处理及运算;ORIGIN 7.5进行图形处理。
2 结果与分析 2.1 树冠结构林隙、林缘、林冠下的无机环境和生物环境都存在不同程度的差异,使得更新幼苗产生不同的适应策略,以保证植株在资源利用和竞争中取得优势。不同生境对天然更新的栓皮栎幼苗的冠长和树冠率影响不大,但显著影响了幼苗冠幅及地径(表 1)。从郁闭的林冠下到林隙直到林缘存在一个资源梯度,最显著的为光照条件的差异。林冠下由于光照水平的降低,栓皮栎幼苗具有明显的开阔型树冠,冠幅随之增加。这表明在庇荫环境条件下,天然更新的栓皮栎幼苗采取了可塑性调节对策,通过冠幅的扩大尽可能多的获取光合有效辐射。栓皮栎天然更新幼苗地径表现为林隙>林缘>林冠下,高度也表现为林隙最大,这与对秦岭北坡栓皮栎实生苗的研究结果[17]一致,林隙较林缘、林冠下更有利于栓皮栎幼苗的生长发育,形成大的实生苗。
| 生境 Habitat | 冠长 Crown length/cm | 冠幅 Crown width/cm | 树冠率 Crown ratio/% | 树高 Height/cm | 地径 Collar diameter/cm | 苗龄 Seedling age/a |
| 不同小写字母表示生境间差异显著(P<0.05) | ||||||
| 林缘Forest edge | 16.7±6.3 | 22.8±7.6 a | 38.5±15.2 | 41.6±16.6 | 0.4±0.1a | 5.0±2.3 |
| 林隙Forest gap | 23.1±7.4 | 32.1±9.6 ab | 43.4±17.7 | 57.0±14.0 | 0.7±0.4b | 5.6±3.1 |
| 林冠下Under canopy | 37.6±12.4 | 45.6±6.4 b | 48.8±14.0 | 42.9±12.8 | 0.4±0.2a | 5.6±2.8 |
| P | 0.207 | 0.045 | 0.608 | 0.394 | 0.033 | 0.856 |
从表 2可看出,不同生境个体间除1级侧枝直径和1级侧枝长度相差不大外,其余各侧枝属性因子间的差异均达显著或极显著水平。从林冠到林隙到林缘,由于环境资源的变化,1级侧枝属性随之发生变化,栓皮栎天然更新幼苗形态对不同生境表现出一定程度的可塑性响应。林冠下栓皮栎幼苗的1级侧枝密度、分枝角度、1级侧枝长度、1级侧枝直径均为各生境之首,且1级侧枝密度与分枝角度与林缘、林隙相比达到显著水平。2级侧枝密度则以林隙中的个体最高,林冠下个体的最低,林缘与林隙的个体差异不显著。说明在林冠下栓皮栎幼苗个体通过抑制主干生长来促进1级侧级生长,不但通过侧枝的水平生长增大冠幅来获取更多的光辐射,而且通过强烈分枝来提高光利用率,但对其高生长不利,可能会导致幼苗的高死亡率。实际调查中也发现林冠下栓皮栎幼苗的顶端枯死现象严重,不利于种群更新。
| 生境Habitat | DSB/(个/m) | DFB/(个/m) | CDFB/ cm | LFB/ cm | AFB/ (°) |
| DSB:2级侧枝密度Density of second order branch;DFB:1级侧枝密度Density of first order branch;CDFB:1级侧枝直径Collar diameter of first order branch;LFB:1级侧枝长Length of first order branch;AFB:分枝角度Angle of first order branch;不同小写字母表示差异显著(P<0.05);不同大写字母表示差异极显著(P<0.01) | |||||
| 林缘Forest edge | 24.7±4.1 B | 4.2±1.2 a | 0.1±0.1 | 7.9±3.6 | 43.5±5.8 a |
| 林隙Forest gap | 32.7±7.6 AB | 7.3±3.1 ab | 0.2±0.1 | 8.8±2.2 | 62.4±11.2 ab |
| 林冠下Under canopy | 5.7±1.1 A | 9.1±4.2 b | 0.2±0.1 | 13.0±3.5 | 66.2±5.8 b |
| P | 0.000 | 0.026 | 0.101 | 0.054 | 0.049 |
从各冠层平均结果看(图 1),1级侧枝在各种生境条件下的垂直分布趋势不同。在林冠下,栓皮栎幼苗的1级侧枝主要分布在树冠最高层次中;林隙中幼苗的1级侧枝主要分布在Ⅰ、Ⅱ两层,且在这两层的分布比例基本相同(P>0.05);在林缘处,由于处于全光环境下,栓皮栎幼苗的1级侧枝主要分布在次高层次中。2级侧枝在不同生境中的垂直分布也不同。林缘处的栓皮栎幼苗2级侧枝在第Ⅲ层中所占比例最高,占全冠的50%左右;而林隙和林冠下个体2级侧枝在第Ⅱ层内的比例最大,但在树冠上部3层内的分布较均匀。在3种生境中,在树体下端产生分枝的栓皮栎幼苗数量较少,都表现出良好的向上生长的态势。

|
| 图 1 同生境中栓皮栎幼苗的侧枝分布属性 Fig. 1 Distributive attributes of branches of Q. variabilis seedlings in different habitats |
在3种生境内栓皮栎幼苗发生侧枝的分枝概率存在差异(表 3),发生5个以上1级侧枝的分枝概率以在林冠下最大,且1级侧枝的平均分枝数高达5.1个;而2级侧枝的平均分枝数则以林隙处最大,为4.6个,不发生2级侧枝的分枝概率以林缘处最大,这与表 2的结果一致,在林冠下通过强烈的1级分枝增加对光的吸收面积,但1级分枝的生长又会抑制2级侧枝的发生;林缘处则由于强烈的光照抑制了1级侧枝的发生。可见为了充分利用各生境内的资源,栓皮栎幼苗通过合理控制分枝来实现其正常生长发育过程,形成对不同生境的适应策略。
| 分枝数i Branching number | 1级侧枝First order branch | 2级侧枝Second order branch | |||||
| 林缘 Forest edge | 林隙 Forest gap | 林冠下 Under canopy | 林缘 Forest edge | 林隙 Forest gap | 林冠下 Under canopy | ||
| 分枝概率Pi | i=0 | 0.3 | 0.0 | 0.0 | 0.7 | 0.3 | 0.6 |
| Branching probability | 1≤i<5 | 0.5 | 0.8 | 0.4 | 0.0 | 0.3 | 0.2 |
| i≥5 | 0.2 | 0.2 | 0.6 | 0.3 | 0.4 | 0.2 | |
| 平均分枝数E(x) Mean branching number | 1.9 | 3.2 | 5.1 | 0.4 | 4.6 | 2.3 | |
3 种生境中,栓皮栎幼苗的分枝角度在树干上从下自上变化格局有差异,在Ⅰ、Ⅱ、Ⅲ层差异不显著,在IV层中差异显著(图 2)。在林冠下1级侧枝在4个树冠层内的平均分枝角度相差不大,分别为68.3°(Ⅰ)、66.2°(Ⅱ)、70.2°(Ⅲ)、70°(Ⅳ),。林隙中1级侧枝的分枝角度在III层内最大(63.8°),Ⅳ层最小,而且Ⅰ、Ⅱ、Ⅲ层的平均分枝角度变化较小,其变化幅度不到5°。林缘处1级侧枝的分枝角度在I层内最大,从树干上部到下部平均分枝角度逐渐减小,减小的幅度高达40°,但在Ⅱ、Ⅲ层相差不大,仅减少3°。分枝角度的大小不仅影响树干的几何形态,还影响林分的光照强度。林缘处的栓皮栎幼苗接受阳光充分,生长旺盛,下部枝条上举有利于吸收直射光以加强光合作用,进行营养积累;生活在林冠下的栓皮栎幼苗受到树冠的荫蔽,植株下部枝条接受的光照较弱,枝条渐趋平展,有利于吸收林内的散射光。

|
| 图 2 不同生境中栓皮栎幼苗分枝角度的变化格局 Fig. 2 Change of branching angle of Q. variabilis seedlings in different habitats |
叶片在植物的生活史中起着重要的作用,枝条上叶片的大小及数量分布等会影响到植物对碳的获取,并进一步影响到枝系的生长和生存,因此,叶片大小和数量也是构成植冠形态的重要因素[2]。在不同生境条件下,栓皮栎幼苗的叶片特征存在差异(表 4)。从林缘、林隙到林冠下,叶长、叶宽、单叶面积、叶面积指数逐渐降低,但差异没有达到显著水平,且在3种生境中的叶面积指数小于或等于1,说明叶片在树冠中基本上不发生重叠现象;数量叶密度和比叶面积则逐渐增大,且数量叶密度在各生境间的差异极显著,以林冠下最大,面积叶密度也以林冠下最高。3种生境中,叶片数量均在树冠第Ⅱ层位置上达到最高水平,树冠的底部最低。林冠下栓皮栎幼苗的叶片数量在Ⅰ、Ⅱ、Ⅲ冠层中的分配比例均高于其它生境个体(图 3),主要着生在树冠中、上层侧枝上,且在第Ⅰ层与林缘处幼苗差异显著(P<0.05)
| 生境Habitat | LL/cm | LW/cm | LLWR | LA/ cm2 | LAI | QLD/ (片/m) | LAD/ (m2 /m) | SLA/ (cm2 /g) |
| LL:叶长Leaf length;LW:叶宽Leaf width;LLWR:叶长宽比Leaf length-width ratio;LA:平均单叶面积Leaf area;LAI:叶面积指数Leaf area index;QLD:数量叶密度Quantitative leaf density;LAD:面积叶密度Leaf area density;SLA:比叶面积Leaf area per unit dry mass(Specific leaf area) | ||||||||
| 林缘Forest edge | 9.7±1.8 | 4.0±1.0 | 2.5±0.3 | 31.7±14.6 | 1.0±0.6 | 28.1±15.4 B | 1195.2±387.7 | 246.6±132.3 |
| 林隙Forest gap | 9.6±2.1 | 3.8±0.9 | 2.6±0.3 | 29.6±12.7 | 0.8±0.3 | 38.6±19.6 AB | 1067.6±398.6 | 254.3±118.8 |
| 林冠下Under canopy | 8.9±1.9 | 3.6±0.6 | 2.5±0.5 | 25.4±8.0 | 0.6±0.3 | 78.7±48.2 A | 2724.3±582.2 | 267.2±132.9 |
| 显著性P-value | 0.604 | 0.531 | 0.865 | 0.503 | 0.081 | 0.003 | 0.066 | 0.940 |

|
| 图 3 不同生境中栓皮栎幼苗叶片的分布属性 Fig. 3 Distribution characteristics of leaves for Q. variabilis seedlings in different habitats |
构型是植物本身与其生长环境相互作用的结果,一般认为在特定生境中,植物体可以通过性状的可塑性表达,形成与环境相适应的有利特征[11]。因而,对不同生境中的植物构型进行比较一定程度上可以反映植物体的不同适应对策。研究者普遍认为,体积小、着生大量高重叠率短枝叶片的紧密型树冠通常被认为是树木对强光环境的一种适应行为[21];在荫庇环境中,着生大叶片的长侧枝所构成的开阔型树冠,是提高耐阴树种个体捕光能力的象征[22],这与本研究的结果一致。与林隙、林缘这两种生境相比,林冠下的光照条件差,在林冠下生长的栓皮栎幼苗形成长枝,发生5个以上1级侧枝的分枝概率最大,1级侧枝的分枝数高,在4个树冠层内的平均分枝角度也都最大;同时幼苗的叶片密度也逐渐提高,这种树冠结构变化是提高幼苗对光的捕获能力的一种有益适应。可见,生活在荫蔽条件下的栓皮栎幼苗,通过侧枝的水平生长、分枝角度加大,强烈分枝的来增大冠幅,形成较明显的开阔型树冠。树冠的这种侧向发展有利于幼树降低自我遮荫程度,并使之沿资源获得的水平梯度占据资源相对较丰富的微环境[23, 24]。
不同环境条件下的植物个体,在主导环境因子的诱导下,生长对策存在着一定的差异性:Horn[25]认为生活在荫蔽生境中的个体主要对策是尽可能减少叶的相互重叠,因而叶呈单层排列,叶面积指数较小。本研究也发现,与其它两种生境相比,生活在林冠下的栓皮栎幼苗个体,叶长、叶宽、单叶面积以及叶面积指数小,叶面积指数为0.6±0.3,明显小于1,说明叶片在树冠中不发生重叠现象,对光的捕获能力不会有显著影响[26]。比叶面积反映了植物对碳的获取与利用的平衡关系,即碳源与汇的关系,与植物的相对生长速率和光合能力有紧密的关系[27, 28]。林冠下栓皮栎幼苗的比叶面积增加,使叶的光合能力提高,增强了对于荫蔽环境的适应能力。由于植物固着生长,在整个生活史中其自身的生理生长及外部环境都处于不断变化中,植物通过构型特征的可塑性以及不断变化来适应环境[29]。栓皮栎幼苗通过枝叶形态上的可塑性来完成对不同生境中资源异质性的反应,这是它适应环境的对策之一。
在天然更新过程中,微生境的有利与否对更新幼苗的发生、定居与生长具有非常重要的作用。相对于非林隙,林隙更有利于幼苗更新[30],是森林循环更新的主要方式[31]。本研究中,林隙内更新的栓皮栎幼苗的树高、地径明显优于林缘和林冠下幼苗,这是因为林隙中光和水分的有效性增加,凋落物分解加快[30],所以林隙中栓皮栎幼苗能更好的利用有效资源,生长明显较快,缩短了苗木进入主林层的时间,林隙对栓皮栎种群更新有利。在今后的栓皮栎次生林经营过程中,可以通过适当间伐来增加林隙的数量,从而增加苗木可利用的有效资源,为森林更新和森林结构的优化的提供有利条件。
| [1] | White J. The plant as a metapopulation. Annual Review of Ecology and Systematics, 1979, 10(1): 109-145. |
| [2] | 陈波, 达良俊. 栲树不同生长发育阶段的枝系特征分析. 武汉植物学研究, 2003, 21(3): 226-231. |
| [3] | Sussex L M, Kerk N M. The evolution of plant architecture. Current Opinion in Plant Biology, 2001, 4(1): 33-37. |
| [4] | 李俊清, 臧润国, 蒋有绪. 欧洲水青冈(Fagus sylvatical L.)构筑型与形态多样性研究. 生态学报, 2001, 21(1): 151-155. |
| [5] | 林勇明, 洪滔, 吴承祯, 洪伟, 胡喜生, 范海兰, 宋萍. 不同起源与方位下桂花的构型差异. 福建林学院学报, 2009, 29(2): 97-102. |
| [6] | 郑万均. 中国树木志 (第二卷). 北京: 中国林业出版社, 1985: 2330-2331. |
| [7] | Oohata S, Shidei T. Studies on the branching structure of trees. I. bifurcation ratio of trees in Horton's law. Japanese Journal of Ecology, 1971, 21: 7-14. |
| [8] | Mcmahon T A, Kronauer R E. Tree structures: deducing the principle of mechanical design. Journal of Theoretical Biology, 1976, 59(2): 443-466. |
| [9] | Pickett S T A, Kempf J S. Branching patterns in forest shrubs and understory trees in relation to habitat. New Phytologist, 1980, 86(2): 219-228. |
| [10] | 孙书存, 陈灵芝. 辽东栎植冠的构型分析. 植物生态学报, 1999, 23(5): 433-440. |
| [11] | 孙书存, 陈灵芝. 不同生境中辽东栎的构型差异. 生态学报, 1999, 19(3): 359-364. |
| [12] | 孙书存, 陈灵芝. 辽东栎芽库统计: 芽的命运. 生态学报, 2001, 21(3): 385-390. |
| [13] | 吴明作, 刘玉萃, 姜志林. 栓皮栎种群生殖生态与稳定性机制研究. 生态学报, 2001, 21(2): 225-230. |
| [14] | 韩照祥, 山仑. 栓皮栎种群变异与适应对策研究. 林业科学, 2005, 41(6): 16-22. |
| [15] | 张文辉, 卢志军. 栓皮栎种群的生物学生态学特性和地理分布研究. 西北植物学报, 2002, 22(5): 1093-1101. |
| [16] | 卢彦昌. 不同经营措施对栓皮栎群落稳定性的影响 [D]. 杨凌: 西北农林科技大学, 2007. |
| [17] | 马莉薇, 张文辉, 薛瑶芹, 马闯, 周建云. 秦岭北坡不同生境栓皮栎实生苗生长及其影响因素. 生态学报, 2010, 30(23): 6512-6520. |
| [18] | 徐程扬. 不同光环境下紫椴幼树树冠结构的可塑性响应. 应用生态学报, 2001, 12(3): 339-343. |
| [19] | 胡喜生, 洪伟, 吴承祯, 洪滔, 范海兰, 宋萍. 不同光环境下木荷幼苗树冠结构的可塑性响应. 植物资源与环境学报, 2006, 15(2): 55-59. |
| [20] | 祁建, 马克明, 张育新. 北京东灵山不同坡位辽东栎(Quercus liaotungensis)叶属性的比较. 生态学报, 2008, 28(1): 122-128. |
| [21] | Cornelissen J H C. Aboveground morphology of shade-tolerant Castanopsis fargesii saplings in response to light environment. International Journal of Plant Sciences, 1993, 154(4): 481-495. |
| [22] | Gilmore D W, Seymour R S. Crown architecture of Abies balsamea from four canopy positions. Tree Physiology, 1997, 17(2): 71-80. |
| [23] | Poulson T L, Platt W J. Replacement patterns of beech and sugar maple in Warren Woods, Michigan. Ecology, 1996, 77(4): 1234-1253. |
| [24] | Givnish T J. Leaf and canopy adaptations in tropical forests//Medina E, Mooney H A, Vazquez-Yznez C. Physiological Ecology of Plants of the Wet Tropics: Tasks for Vegetation Science. Netherlands: Springer, 1984: 51-84. |
| [25] | Horn J S. Adaptation from the perspective of optimality//Solbrig O T, Jain S, Johnson G B, Raven P H. Topics in plant population biology. New York: Columbia University Press, 1979: 48-61. |
| [26] | Planchais I, Sinoquet H. Foliage determinants of light absorption in sunny and shaded branches of Fagus sylvatica (L.). Agricultural and Forest Meteorology, 1998, 89(3/4): 241-253. |
| [27] | Wilson P J, Thompson K, Hodgson J G. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytologist, 1999, 143(1): 155-162. |
| [28] | Wright I J, Reich P B, Cornelissen J H C, Falster D S, Garnier E, Hikosaka K, Lamont B B, Lee W, Oleksyn J, Osada N, Poorter H, Villar R, Warton DI, Westoby M. Assessing the generality of global leaf trait relationships. New Phytologist, 2005, 166(2): 485-496. |
| [29] | Valladares F, Gianoli E, GóMez J M. Ecological limits to plant phenotypic plasticity. New Phytologist, 2007, 176(4): 749-763. |
| [30] | 宋新章, 张智婷, 肖文发, 李冬生, 张慧玲. 长白山杨桦次生林采伐林隙幼苗更新动态. 林业科学, 2008, 44(3): 13-20. |
| [31] | Silbernagel J, Moeur M. Modeling canopy openness and understory gap patterns based on image analysis and mapped tree data. Forest Ecology and Management, 2001, 149(1/3): 217-233. |