 2015, Vol. 35
2015, Vol. 35文章信息
- 薛盈文, 张英华, 黄琴, 王志敏
- XUE Yingwen, ZHANG Yinghua, HUANG Qin, WANG Zhimin
- 窄行匀播对晚播冬小麦群体环境、个体性状和物质生产的影响
- Effects of narrow row spacing and uniform sowing on canopy environment,individual plant traits, and biomass production in late-sowing winter wheat
- 生态学报, 2015, 35(16): 5545-5555
- Acta Ecologica Sinica, 2015, 35(16): 5545-5555
- http://dx.doi.org/10.5846/stxb201402150258
-
文章历史
- 收稿日期: 2014-02-15
- 修订日期: 2014-10-08






2. 黑龙江八一农垦大学农学院, 大庆 163319
2. College of Agronomy, Heilongjiang Bayi Agricultural University, Daqing 163319, China
我国华北北部平原区光温资源有限,水资源十分紧缺,为提高“冬小麦—夏玉米”一年两熟制资源利用效率,近年来广泛推行“冬小麦晚播、夏玉米晚收”的“双晚”栽培模式[1, 2]。在此模式中,冬小麦晚播不仅延长了夏玉米生育期、提高了夏玉米产量,而且减少了冬小麦前期水肥消耗、有利于节水生产[2, 3]。但晚播冬小麦因个体发育和生物量受到限制,必须通过增加播种量、增加群体数量来保证足够穗数,从而稳定或提高群体产量。在播期推迟、播量增大的情况下,如果措施不当,如行距配置不合理、整地播种质量不高、落籽不均等,极易造成行内苗间过度拥挤竞争,使得弱株率增加、群体质量恶化。因此,如何调节群体分布形式、减小个体间竞争是晚播小麦极为重要的生产问题。关于小麦群体的调节,国内外研究报道较多,多数是通过调整播量和行距来协调群体内个体之间的矛盾[4, 5, 6, 7, 8]。但有关行距对小麦产量的影响效应,在不同生态区、不同播期、播量以及不同品种条件下可能是不一样的[4,9,10,11,12],我国北方干旱地区晚播小麦节水高产栽培的适宜行距仍有待明确。近年来,在我国部分地区推广了“宽幅精播技术”[13, 14],在适宜播期和较低播量下通过扩大行距和增加播幅宽度来提高群体均匀性和群体产量,取得较好效果。但在晚播大播量条件下如何调整播种方式来优化群体结构、提高群体均匀性还有待探讨。本研究冬小麦晚播节水栽培,通过缩小行距,增加行数,扩大行内株距,可以减少行内竞争,并能减少行间土壤水分蒸发[2, 15];在缩行的基础上,优化调整行内株间分布形式可能是进一步提高群体均匀性的重要措施。但缩小行距的效应机制以及合理的行内株间配置方式及其效应仍需要深入的研究。本项研究在晚播、高密度栽培条件下比较考察并分析了不同行距和行内植株分布形式对群体内微环境、冠层结构、物质积累和转运及产量的影响效应,以期为晚播小麦节水高产栽培群体调控提供部分理论依据和技术指导。
1 材料与方法 1.1 试验条件与试验设计试验于2011—2012和2012—2013年度在中国农业大学吴桥实验站进行,试验地为壤质底黏潮土,两年度内小麦播种前耕层土壤的养分含量见表1。两个年度的小麦前茬均为玉米。小麦晚播,2011—2012年度的播种日期为10月29日,2012—2013年度的播种日期为10月25日,田间保苗数量均为675万株/hm2,两个年度生育期内降雨分布如图1所示。小麦供试品种为济麦22,参照冬小麦节水省肥简化栽培模式[2]进行水肥管理。施肥量为:尿素 225 kg/hm2、磷酸二铵 300 kg/hm2、硫酸钾 150 kg/hm2、硫酸锌 15 kg/hm2,均作底肥一次性施入;在足墒播种基础上,春季灌拔节水和开花水。
| 年度 Year |
全氮 Total nitrogen/ (g/kg) |
碱解氮 Alkali-hydrolyzable Nitrogen/ (mg/kg) |
速效磷 Rapidly available Phosphorus/ (mg/kg) |
速效钾 Rapidly available Potassium/ (mg/kg) |
有机质 Organic matter/ (g/kg) |
pH |
| 2011—2012 | 0.92 | 57.8 | 44.6 | 91.0 | 10.7 | 7.8 |
| 2012—2013 | 0.90 | 55.4 | 45.5 | 88.3 | 10.1 | 7.8 |

|
| 图 1 2011—2013年度冬小麦生育期内降雨分布 Fig.1 Rainfall distribution during winter wheat growing seasons in 2011—2013 |
试验采用2因素裂区设计,以行距为主区,设置3种不同行距处理:10、15、20 cm;以种子分布形式为副区,设两种种子分布形式处理:(1) 随机分布形式(R):常规播种,行内种子间距不统一;(2) 均匀分布形式(A):行内双小行等宽 (宽度2 cm) 等距播种,种子间距基本一致。对于3种行距内的均匀分布处理,种子在行带内的分布形式如图2所示,为了确保落种数量和距离的相对准确,均采用人工将种子按照理论距离(行带内双小行间距宽度2 cm,行内的种子间距列于表2)播前粘贴在薄卫生纸上,播种时铺覆于行内。各行距的随机分布形式处理是模拟一般生产田的常规播种方法,用开沟播种器人工开沟行,沟内条状撒种,播幅2 cm以内,种子在行内随机撒落,种子间距不均匀。试验小区面积6 m2,重复3次。

|
| 图 2 各行距内均匀分布形式示意图 Fig.2 The patterns of uniform seed distribution in different row spacing |
| 行距Row spacing/cm | 10 | 15 | 20 | |||
| 行内种子分布形式Seed distribution | 随机 | 均匀 | 随机 | 均匀 | 随机 | 均匀 |
| 处理代号Treatment code | 10R | 10A | 15R | 15A | 20R | 20A |
测定时期为开花期晴朗天气,测定部位为各处理行间的冠层下部(距地上部10 cm),使用浙江托普仪器公司TNHY-4型环境监测仪的光照、温度和湿度传感器,在白天定点(3点/处理)、定时(时间间隔为15 min)进行连续监测,仪器自动记录不同测定时间冠层下部的温度(℃)、相对湿度(RH%)和光合有效辐射(μmol m-2 s-1),同时测定冠层上部的上述3个指标,将冠层下部与冠层顶部光照强度的比值作为相对光强值。
1.2.2 冠层性状分析在测定上述指标的区域,利用CI-110冠层分析仪在冠层底部垂直向上截取冠层图像,将天顶角划为5 环,步长值设定为12°,5个天顶角度分别为13°、25°、37°、49°和61°,测定时间选择在傍晚,操作时鱼眼镜头放在行间地表之上1cm 处,每个处理测定5点。将获得的图像用CI-110软件进行图像提取和分析,可以得到以下参数:叶面积指数(LAI)、平均叶倾角(MLA) 、散射辐射透过系数(TD)、不同天顶角内直接辐射透过系数(TR)和不同角度冠层的消光系数(K)。
1.2.3 植株性状及产量测定在小麦开花期和成熟期于各处理小区取植株样段(2 cm×50 cm)测定植株干物重。在成熟期,于各处理区随机调查6行、每行1 m长的行段内有效穗数,同时,在每个处理内随机选9个样点,每个样点连续取20个成穗单株,调查各株株高、穗长、小穗数、穗粒数、穗粒重和单株(单茎)干重等性状,分株记载,分析各性状指标的株间变异性。每个小区选择2 m2收获测产,以14%含水量为基础计算实际产量。
1.3 数据分析和统计方法用Excel 2010整理群体内环境监测、冠层分析和产量数据,使用DPS7.05进行数据统计分析。
花期前后贮藏物以及向籽粒的转运数量和贡献率的计算方法如下[15]:
营养器官花前贮藏物质转运量= 开花期干重-成熟期干重(籽粒除外)
营养器官花前贮藏物对籽粒产量的贡献率= 花前贮藏物质转运量 / 成熟期籽粒干重×100%
花后干物质积累量= 成熟期籽粒干重-花前贮藏物质转运量
花后同化物对籽粒产量的贡献率= 花后干物质积累量 / 成熟期籽粒干重×100%
2 结果与分析 2.1 不同处理冠层结构与微环境变化 2.1.1 冠层结构性状合理的冠层结构是提高群体光能利用率和产量的基础,叶面积指数和叶倾角是冠层结构的重要指标,与群体光能截获密切相关[16]。对开花期的冠层结构分析(表3)表明,3种行距和2种种子分布方式处理比较,叶面积指数(LAI)以较小行距处理高于较大行距处理,同一行距下以均匀方布处理高于常规随机分布处理;冠层平均叶倾角(MLA)则相反,以较小行距处理低于较大行距处理,以均匀方布处理低于常规随机分布处理。具体而言,不同处理的叶面积指数大小依次是10A>10R>15A>15R>20A>20R,叶倾角大小顺序是20R>20A>15R>15A>10R>10A,20R处理叶面积指数最低、叶倾角最大,10A 处理叶面积指数最高、叶倾角最小。冠层内散射辐射透过系数(TD)随着行距的加宽而增大、随着行内植株分布均匀度的增加而减小,20 cm行距显著高于其它两种行距处理。冠层直接辐射透过系数(TR)能反映入射光被冠层的截获程度,TR值越高,群体的光截获越少,漏光损失就越大。不同天顶角内的TR值总体上随着天顶角的增大而降低、随着行距的增加而增大,其中R20处理在13°天顶角内的TR值达到0.32。同一行距内,均匀分布形式(A)的TR值低于对应的随机分布处理(R)。冠层消光系数(K)的变化趋势与TR值不同,K值随着天顶角、行内植株分布均匀度的增加而增大,随着行距的增加而降低。
| 处理 Treatment |
叶面积指数 Leaf area index(LAI) |
平均叶倾角 Mean leaf inclination angle(MLA) |
散射辐射透过系数 Transmission coefficient for diffuse penetration (TD) |
不同角度的直接辐射透过系数 Transmission coefficient for radiation penetration(TR) |
不同角度的消光系数 Light extinction coefficient(K) |
||||||||
| 13° | 25° | 37° | 49° | 61° | 13° | 25° | 37° | 49° | 61° | ||||
| 同一列数据后不同字母表示处理间差异达5%显著水平 | |||||||||||||
| 10R | 7.50 b | 63.90 bc | 0.07 b | 0.22c | 0.19a | 0.13c | 0.05cd | 0.02bc | 0.41a | 0.42ab | 0.54b | 0.75a | 1.08a |
| 10A | 8.10 a | 60.20 c | 0.07 b | 0.15d | 0.15a | 0.11c | 0.04d | 0.02c | 0.43a | 0.45a | 0.63a | 0.75a | 1.09a |
| 15R | 6.90 c | 69.70 ab | 0.11 b | 0.26bc | 0.21a | 0.17abc | 0.07c | 0.03abc | 0.34ab | 0.40b | 0.53b | 0.74a | 1.07ab |
| 15A | 7.50 b | 65.30 bc | 0.10 b | 0.24bc | 0.19a | 0.15bc | 0.06cd | 0.03abc | 0.37a | 0.40b | 0.53b | 0.74a | 1.08a |
| 20R | 6.70 c | 76.20 a | 0.17 a | 0.32a | 0.26a | 0.22a | 0.18a | 0.05a | 0.25b | 0.35d | 0.50c | 0.73a | 0.99b |
| 20A | 6.80 c | 70.70 ab | 0.16 a | 0.28ab | 0.24a | 0.20ab | 0.12b | 0.05ab | 0.26b | 0.36cd | 0.50c | 0.74a | 1.06ab |
(1) 光照
开花期3种行距内不同处理冠层下部的相对光强(下部实际光强与冠层顶部自然光强的比值)变化如图3、4所示,各处理一日中冠层下部的相对光强均在10:00—14:00时段内变化幅度较大,且行距越宽变化幅度越大,20R下层最大相对光强值在0.3以上。各行距内均匀分布群体(A)相对光强值小于对应的随机分布群体(R),以10 A处理的下层相对光强最小,其在中午的相对光强值为0.06。这也说明,开花期窄行距处理的叶片空间分布较为均匀,在全天各个时期都能较充分截获光能,漏光损失少。

|
| 图 3 各行距内随机分布处理冠层下部相对光强的变化趋势 Fig.3 Variation trend of relative light intensity at the bottom of canopy for random seed distribution treatment in different row spacing(2012—2013) |

|
| 图 4 各行距内随机分布与均匀分布处理冠层下部相对光强的变化 Fig.4 Variation trend of relative light intensity at the bottom of canopy for random and uniform seed distribution treatments in different row spacing(2012—2013) |
(2) 温度
开花期群体冠层下部温度的变化如图5、图6所示。一日中不同处理的下层温度有所差异,以中午时段差异较为明显,20R处理冠层透光多,下层温度上升快,其在中午的温度最高。3种行距的两种分布形式比较,10 cm行距的两种分布形式(10R和10A)冠层下部温度在中午之前比较接近,13:00后10A低于10R的趋势比较明显;15 cm和20 cm行距的均匀分布处理(A)各个监测点的温度均低于对应的随机分布处理(R),特别是20A和20R两处理冠层下部温度的差异更为明显。结果表明,缩小行距、增加行内植株分布均匀性可以降低群体冠层下部温度,特别是能明显降低午间高温阶段冠层内部温度。

|
| 图 5 各行距内随机分布处理冠层下部与冠层上部的温度变化趋势 Fig.5 Variation trend of temperature at the bottom and top of canopy for random distribution treatment in different row spacing (2012—2013) |

|
| 图 6 各行距内随机(R)与均匀(A)分布群体冠层下部和冠层顶部的温度变化趋势 Fig.6 Variation trend of temperature at the bottom and top of canopy for random and uniform distribution treatments in different row spacing (2012—2013) |
(3) 相对湿度
不同处理冠层下部相对湿度变化如图7、图8所示。在中午之前,3种行距内随机分布处理冠层下部相对湿度的大小依次为:10R>15R>20R,即:随着行距缩小,冠层下部的相对湿度呈现增加的趋势。在中午之后,不同行距的相对湿度差异很小。不同行距2种分布形式比较,10R和10A两个处理冠层下部相对湿度差异不明显,仅在12:00以后10A处理的相对湿度较高于10R;其它2个行距处理,冠层相对湿度均表现为均匀分布处理(A)明显高于随机分布处理(R)。

|
| 图 7 各行距内随机分布处理冠层下部和冠层上部相对湿度的变化趋势 Fig.7 Accumulation and translocation of dry matter in different treatments (2012—2013) |

|
| 图 8 各行距内随机(R)与均匀(A)分布处理冠层下部和冠层顶部相对湿度的变化趋势 Fig.8 Variation trend of relative humidity at the bottom of canopy for random and uniform distribution treatments in different row spacing (2012—2013) |
不同行间和同一行不同行段间最终穗数的差异性可反映群体分布均衡性。成熟期在每个处理小区分别调查6个1 m长的行段内实际穗数,结果表明,虽然不同处理的群体平均穗数并没有显著差异,但样段间成穗数的变异程度存在较大差异(表4),随着行距增加而增大,即:10 cm<15 cm<20 cm;在3种行距内,样段内成穗个数的变异系数随着行内落种均匀度的提高而相应减少,即3种行距内的两种分布形式的变异系数比较结果是:A<R。
| 处理 Treatment |
2011—2012 | 2012—2013 | ||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | CV/% | 1 | 2 | 3 | 4 | 5 | 6 | CV/% | |
| 10R | 76 | 72 | 86 | 76 | 70 | 88 | 9.5 | 87 | 83 | 78 | 74 | 88 | 62 | 12.4 |
| 10A | 82 | 78 | 74 | 81 | 79 | 74 | 4.4 | 79 | 80 | 78 | 76 | 70 | 85 | 6.3 |
| 15R | 104 | 130 | 116 | 105 | 132 | 115 | 10.2 | 117 | 112 | 106 | 100 | 143 | 124 | 13.0 |
| 15A | 120 | 121 | 114 | 124 | 126 | 106 | 6.2 | 128 | 123 | 109 | 110 | 112 | 120 | 6.7 |
| 20R | 144 | 183 | 160 | 130 | 164 | 155 | 11.6 | 120 | 151 | 130 | 176 | 158 | 164 | 14.1 |
| 20A | 168 | 165 | 140 | 150 | 160 | 153 | 6.7 | 152 | 128 | 163 | 134 | 170 | 159 | 11.0 |
不同行、同一行不同行段内植株间农艺性状的差异性可反映群体内个体生长的均衡性。不同处理间单株性状指标的比较结果(表5)中,株高和单株干重均表现为10A显著高于20R和20A处理,其它处理间没有显著差异;穗粒重处理间差异显著,随着行距变窄、行内种子分布均匀度提高而显著提高。在单株性状指标中,穗粒重和单株干重的变异程度较大,处理20R的变异系数分别为34.7%和31.6%。对于同一行距内的两种分布形式,均匀分布处理(A)的各项单株指标值均高于对应的随机分布处理(R)。总的看来,随着行距的增加,各个单株指标的变异程度增加,提高行内植株分布的均匀度会降低相应行距内各个指标的变异度。综合分析穗粒重与各项指标变异系数的相关性(表6)表明,穗粒重与所测各指标的变异系数呈显著或极显著负相关,说明单株指标变异程度大小会显著影响个体的单株生产能力。
| 处理 Treatment |
株高 Plant height/cm |
穗长 Spike length/cm |
小穗数 Spikelet number per ear |
穗粒数 Grains per ear |
穗粒重 Grain weight per ear/g |
单株干重 Single plant weight/g |
||||||
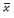 |
CV/% | |
CV/% | |
CV/% | |
CV/% | |
CV/% | |
CV/% | |
| 10R | 71.1 ab | 6.7 | 7.8 a | 8.9 | 15.2 ab | 12.3 | 34.0 ab | 23.6 | 1.12 b | 31.1 | 2.4 ab | 26.6 |
| 10A | 72.6 a | 5.8 | 7.9 a | 8.8 | 16.0 a | 11.8 | 35.1 ab | 23.3 | 1.18 a | 29.1 | 2.5 a | 26.3 |
| 15R | 71.1 ab | 7.7 | 7.8 a | 9.6 | 15.1 ab | 13.5 | 33.1 b | 26.1 | 1.08 c | 32.7 | 2.3 ab | 30.1 |
| 15A | 71.2 ab | 7.1 | 7.9 a | 9.5 | 15.2 ab | 13.0 | 34.2 ab | 24.5 | 1.13 b | 31.6 | 2.3 ab | 29.3 |
| 20R | 69.6 b | 7.9 | 7.6 a | 9.8 | 15.0 ab | 14.9 | 33.0 b | 27.7 | 1.04 c | 34.7 | 2.2 b | 31.6 |
| 20A | 70.0 b | 6.0 | 7.8 a | 9.7 | 15.0 ab | 13.7 | 34.0 ab | 25.6 | 1.11 b | 34.2 | 2.3 b | 29.6 |
| 性状指标Trait index | CV 株高 | CV 穗长 | CV 小穗数 | CV 穗粒数 | CV 穗粒重 | CV 单株干重 |
| CV 株高 | 1 | |||||
| CV 穗长 | 0.2 | 1 | ||||
| CV 小穗数 | 0.12 | 0.42 ** | 1 | |||
| CV 穗粒数 | 0.25 | 0.35 ** | 0.38 ** | 1 | ||
| CV 穗粒重 | 0.18 | 0.35 ** | 0.39 ** | 0.44 ** | 1 | |
| CV 单株干重 | 0.37 ** | 0.29 * | 0.47 ** | 0.53 ** | 0.54 ** | 1 |
| 穗粒重Grain weight per ear | -0.32 * | -0.47 ** | -0.49 ** | -0.51 ** | -0.55 ** | -0.55 ** |
考察不同处理的物质积累与运转(表7),可见,不同处理成熟期的总生物量表现为10A>10R>15A>15R>20A>20R,10A、10R、15A处理间没有显著差异,但均显著高于15R、20R、20A处理。花前物质积累量在不同处理间的差异性与总生物量的差异性表现相似,花前贮藏物质向籽粒的转运量和对籽粒贡献率则以10A、10R显著低于其他处理。花后物质积累量以10A最高,各处理依次为10A>10R>15A>15R>20A>20R,处理间差异达到显著水平。此结果说明,缩小行距、增加行内均匀度可以增加群体物质生产量,特别是显著增加了花后物质积累量及其对产量的贡献率。
| 处理 Treatment |
总生物量 Total biomass/ (kg/hm 2) |
花前物质 Dry matter before anthesis |
花后物质 Dry matter after anthesis |
籽粒干物重 Grain dry matter/ (kg/hm 2) |
|||
| 积累量 Accumulation Amount/ (kg/hm 2) |
转运量 Translocation amount/ (kg/hm 2) |
对籽粒贡献率 Contribution rate to grain weight/% |
积累量 Accumulation amount/ (kg/hm 2) |
对籽粒贡献率 Contribution rate to grain weight/% |
|||
| 10R | 18423 a | 12576 a | 2147 b | 27 | 5846 b | 73 | 7993 a |
| 10A | 19110 a | 12648 a | 2065 b | 25 | 6262 a | 75 | 8327 a |
| 15R | 16646 b | 12006 b | 2552 a | 35 | 4640 d | 65 | 7192 c |
| 15A | 17162 a | 12181 a | 2475 a | 32 | 5182 c | 68 | 7656 b |
| 20R | 16350 b | 11801 b | 2565 a | 36 | 4469 e | 64 | 7034 d |
| 20A | 16636 b | 11955 b | 2622 a | 35 | 4682 d | 65 | 7403 bc |
两年的产量结果(表8)表明,在常规随机播种方式下,不同行距的籽粒产量表现为10R >15R >20R,行距间产量差异达显著水平;在均匀播种方式下,不同行距的产量表现为10A >15A >20A,2011—2012年度10A 与 15A处理产量差异不显著,2012—2013年度,10A处理产量显著高于15A和20A处理。在各行距下两种播种方式间比较,产量皆表现为均匀分布高于随机非均匀分布方式,特别是在15 cm和20 cm行距下,两个年度两种播种方式间产量差异均达显著水平。此结果说明,适当缩小行距和提高行内种子分布均匀性皆有利于提高群体产量,在较大行距下比在较小行距下提高行内播种均匀性的增产效果更明显。从产量构成因素分析,不同处理间产量的差异主要由于平均穗粒重的差异,特别是粒重的差异。
| 年度 Year |
处理 Treatment |
穗数 Ear number/ (10 4穗/hm 2) |
穗粒数 Grains per ear |
千粒重 10 3 grain weight/g |
理论产量 Theoretical Yield/ (kg/hm 2) |
实际产量 Actual yield/ (kg/hm 2) |
| 2011—2012 | 10 R | 775 a | 29.0 a | 41.1 ab | 9237 | 8045 b |
| 10 A | 778 a | 31.0 a | 42.3 a | 10201 | 8602 a | |
| 15 R | 771 a | 27.0 b | 40.5 bc | 8431 | 7663 b | |
| 15 A | 781 a | 29.0 a | 41.0 ab | 9286 | 8379 a | |
| 20 R | 766 a | 26.0 b | 39.6 c | 7887 | 6462 c | |
| 20 A | 772 a | 27.0 ab | 40.2 bc | 8379 | 7979 b | |
| 2012—2013 | 10 R | 786 a | 34.0 ab | 38.0 ab | 10155 | 9311 a |
| 10 A | 782 a | 35.0 a | 38.9 a | 10620 | 9519 a | |
| 15 R | 774 a | 33.0 b | 37.7 b | 9757 | 8114 cd | |
| 15 A | 780 a | 34.0 ab | 38.6 ab | 10130 | 8650 b | |
| 20 R | 747 a | 33.0 b | 36.5 c | 8998 | 7754 d | |
| 20 A | 755 a | 34.0 ab | 37.5 b | 9626 | 8391 bc |
播种量、行距和播种方式是建立小麦群体结构的基本措施,也是影响群体发展和群体生产效率的基础因素。在一定的群体规模下,个体的质量决定了群体产量,而个体质量与个体在群体内的分布形式有很大关系,因此,在一定的播期播量下,合理的行距配置和合适的植株排布方式对构建高产高效群体十分重要。前人针对行距与产量的关系有较多研究,而对行内种子分布形式对产量的影响研究较少[17]。通常认为,增大行距会有利于增加群体的通风透光性能,但较大的行距也会增大漏光损失,特别是在小麦生育后期,光截获量与产量密切相关[18, 19],此期宽行距的劣势愈加明显。在晚播小麦生产中,由于播种量较大,个体性能受到群体的限制程度也较大,对行距和播种方式的要求更为严格。本研究表明,在华北平原干旱区小麦晚播、大播量条件下,与常规的20 cm行距相比较,行距缩小到15 cm、特别是缩小到10 cm,群体的结构发生了明显变化,表现在叶面积指数增加,冠层叶倾角减小,基部漏光损失减少,对光能的截获增加。群体环境的动态测定表明,随着行距缩小,冠层下部行间温度降低,特别是中午高温时段降温明显,而一日中相对湿度增加。这一趋势性结果与其他学者相关的报道相似[18, 19, 20],均说明通过缩小行距增加行数,减少了行内种子量,个体的生育微环境可以得到均衡改善。缩小行距使群体内温度降低、湿度增加的同时,也减少了行间土壤水分的蒸发[21],这对小麦节水栽培具有重要意义。本试验结果还表明,行内植株的排布方式对群体结构和环境也有很大影响,在不同的行距下,行内植株等距离均匀化分布处理与随机化分布处理相比,上述群体结构和微环境指标也均得到有效改善,且行距越大,其效果越明显,说明在较大行距下,行内种子量较大,随机播种个体分布不均匀,容易造成局部株间过于紧密,营养和环境条件不均衡,群体质量不佳。调查发现,较小行距和均匀分布处理不同行段间成穗数趋向一致,穗层均匀性明显高于较大行距和随机分布处理。分析群体内个体生长性状的变异性表明,缩小行距和提高播种均匀性使株高、单株重、穗粒数和穗粒重等性状的株间变异程度降低,而个体性状变异系数的降低是与群体平均穗粒重的提高密切相关的,也即提高群体内个体的均匀性可以提高个体平均产量。这一结果也给我们重要启示,在当前高产田穗数已达饱和的条件下,如何通过提高穗粒重来突破产量限制?从本试验看,提高群体内个体分布的均匀性是提高群体平均穗粒重的重要途径。
株行空间配置影响群体结构和环境,必然影响群体物质生产性能和最终产量形成。从群体的物质生产性能看,缩行扩株和均匀播种使小麦花前和花后物质积累量增加,特别是花后物质积累量明显增加。小麦的产量物质一方面来自花后物质生产,另一方面来自于花前贮藏物质的再运转[22]。许多研究表明,花后群体物质积累量越大,产量越高[23, 24]。晚播小麦尽管由于花期推迟,开花—成熟期较短,开花前的贮藏物质对产量有较大贡献[25],但从本试验看,产量物质仍主要来自于花后物质积累,不同处理花后物质对产量的贡献率为64%—75%。随着行距缩小,花后物质积累及其对产量的贡献增加,在同一行距下均匀分布处理花后物质及贡献率高于随机分布处理。后期物质积累的差异导致了最终穗粒重的差异,最终导致产量的差异。不同处理最终产量表现为10A>10R>15A>15R>20A>20R,相对于较大行距和随机种子分布处理,较小行距和均匀分布处理显示出明显的增产效应,特别是10A处理的产量优势最为突出。这一结果清楚地表明,小行距和匀株距的结合是晚播小麦大群体栽培的重要技术改良方向。当然,本研究结果是在播种量和播种方式严格控制的试验条件下获得的,可为生产实践合理调控株群配置提供理论依据,但实际的生产应用需要配套解决整地播种环节诸多技术问题。近年来部分地区推广应用“小麦宽幅精播技术”[13],通过农机农艺结合和扩大行距、加宽播幅的形式实现了均匀播种,也证明具有明显增产效应。但在晚播小麦大群体栽培条件下,如何实现机械化窄行匀播仍需要深入研究。
3.2 结论华北平原晚播小麦在高密度播种条件下,适当缩小行距、增加行内种子分布的均匀度,可以优化冠层结构,增加群体光截获率并降低冠层内部的温度,从而改善群体的微环境;可以提高穗群整齐度,减小个体农艺性状的变异度,从而提高个体穗粒重;可以增加群体总生物量,并增加花后物质生产对产量的贡献率,从而提高群体产量。因此,窄行匀播是提高华北平原晚播小麦产量的重要途径。
| [1] | 付雪丽, 张惠, 贾继增, 杜立丰, 付金东, 赵明. 冬小麦-夏玉米"双晚"种植模式的产量形成及资源效率研究. 作物学报, 2009, 35(9): 1708-1714. |
| [2] | 王志敏, 王璞, 李绪厚, 李建民, 鲁来清. 冬小麦节水省肥高产简化栽培理论与技术. 中国农业科技导报, 2006, 8(5): 38-44. |
| [3] | 高海涛, 王育红, 孟战赢, 席玲玲, 段国辉, 温红霞. 小麦-玉米双晚种植对周年产量和资源利用的影响. 麦类作物学报, 2012, 32(6): 1102-1106. |
| [4] | Teich A H, Welacky T, Hamill A, Smid A. Row-spacing and seed-rate effects on winter wheat in Ontario. Canadian Journal of Plant Science, 1993, 73(1): 31-35. |
| [5] | 赵虹, 杨兆生, 闫素红, 王俊娟, 陈长海, 梁文科, 戚廷香. 播种方式对不同类型小麦品种产量性状的影响. 华北农学报, 2000, 15(2): 100-105. |
| [6] | Johnson J W, Hargrove W L, Moss R B. Optimizing row spacing and seeding rate for soft red winter wheat. Agronomy Journal, 1988, 80(2): 164-166. |
| [7] | Rasmussen I A. The effect of sowing date, stale seedbed, row width and mechanical weed control on weeds and yields of organic winter wheat. Weed Research, 2004, 44(1): 12-20. |
| [8] | Frederick J R, Marshall H G. Grain yield and yield components of soft red winter wheat as affected by management practices. Agronomy Journal, 1985, 77(3): 495-499. |
| [9] | Marshall G C, Ohm H W. Yield responses of 16 winter wheat cultivars to row spacing and seeding rate. Agronomy Journal, 1987, 79(6): 1027-1030. |
| [10] | 郭天财, 刘胜波, 冯伟, 杨文平, 王永华, 韩巧霞. 不同种植行距的大穗型小麦品种'兰考矮早八'中几种与旗叶衰老有关的生理指标变化. 植物生理学通讯, 2008, 44(1): 33-36. |
| [11] | 朱云集, 郭汝礼, 郭天财, 张庆友, 王之杰, 王永华. 行距配置与密度对兰考906群体 质量及产量的影响. 麦类作物学报, 2001, 21(2): 62-66. |
| [12] | 赵竹, 曹承富, 乔玉强, 杜世州, 张耀兰, 张明杰. 机播条件下行距与密度对小麦产量和品质的影响. 麦类作物学报, 2011, 31(4): 714-719. |
| [13] | 余松烈, 于振文, 董庆裕, 王东, 张永丽, 姚德常, 王坚强. 小麦亩产789.9kg高产栽培技术思路. 山东农业科学, 2010, (4): 11-12. |
| [14] | 韩惠芳, 赵丹丹, 沈加印, 郎坤, 刘泉汝, 李全起. 灌水量和时期对宽幅精播冬小麦产量及品质特性的影响. 农业工程学报, 2013, 29(14): 109-114. |
| [15] | 张胜全, 方保停, 王志敏, 周顺利, 张英华. 春灌模式对晚播冬小麦水分利用及产量形成的影响. 生态学报, 2009, 29(4): 2035-2044. |
| [16] | 陈素英, 张喜英, 毛任钊, 王彦梅, 孙宏勇. 播期和播量对冬小麦冠层光合有效辐射和产量的影响. 中国生态农业学报, 2009, 17(4): 681-685. |
| [17] | 张维城, 王绍中, 李春喜. 小麦植株分布状况对干物质积累和产量的影响. 河南农业科学, 1995, (6): 1-6. |
| [18] | 陈雨海, 余松烈, 于振文. 小麦生长后期群体光截获量及其分布与产量的关系. 作物学报, 2003, 29(5): 730-734. |
| [19] | 闫长生, 肖世和, 张秀英, 海林. 冬小麦冠层内的光分布. 华北农学报, 2002, 17(3): 7-13. |
| [20] | 杨文平, 郭天财, 刘胜波, 王晨阳, 王永华, 马冬云. 行距配置对'兰考矮早八'小麦后期群体冠层结构及其微环境的影响. 植物生态学报, 2008, 32(2): 485-490. |
| [21] | 陈素英, 张喜英, 陈四龙, 裴冬, 张清涛. 种植行距对冬小麦田土壤蒸发与水分利用的影响. 中国生态农业学报, 2006, 14(3): 86-89. |
| [22] | 郭文善, 封超年, 严六零, 彭永欣, 朱新开, 宗爱国. 小麦开花后源库关系分析, 作物学报, 1995, 21(3): 334-340. |
| [23] | 董树亭. 高产冬小麦群体光合能力与产量关系的研究. 作物学报, 1991, 17(6): 461-469. |
| [24] | 曾浙荣, 赵双宁, 李青. 北京地区高产小麦品种的冠层形成、光截获和产量. 作物学报, 1991, 17(3): 161-170. |
| [25] | 屈会娟, 李金才, 沈学善, 魏凤珍, 王成雨, 郅胜军. 种植密度和播期对冬小麦品种兰考矮早八干物质和氮素积累与转运的影响. 作物学报, 2009, 35(1): 124-131. |