 2015, Vol. 35
2015, Vol. 35文章信息
- 郭婧, 喻林华, 方晰, 项文化, 邓湘雯, 路翔
- GUO Jing, YU Linhua, FANG Xi, XIANG Wenhua, DENG Xiangwen, LU Xiang
- 中亚热带4种森林凋落物量、组成、动态及其周转期
- Litter production and turnover in four types of subtropical forests in China
- 生态学报, 2015, 35(14): 4668-4677
- Acta Ecologica Sinica, 2015, 35(14): 4668-4677
- http://dx.doi.org/10.5846/stxb201312052896
-
文章历史
- 收稿日期:2013-12-05
- 修订日期:2014-09-09






2. 南方林业生态应用技术国家工程实验室, 长沙 410004
2. State Key Laboratory of Ecological Applied Technology in Forest Area of South China, Changsha 410004, China
森林凋落物是森林生态系统养分循环、碳流动的重要环节,作为养分的基本载体,是连接植物与土壤的“纽带”[1],森林植物吸收的养分中,90%以上的氮和磷、60%以上的矿质元素都来自于凋落物归还土壤的养分再循环[2],凋落物的量和分解速率构成森林生态系统的功能过程之一,对森林土壤肥力的维持及其生态恢复和更新具有重要作用[3]。在当今全球变化背景下,凋落物也是森林碳库的重要组成部分,是研究森林生态系统与大气间碳交换的基本参数[4]。因此,深入研究各类森林凋落物的特征对正确认识全球森林碳循环具有重要意义[5, 6, 7, 8]。近十几年来,随着我国对天然林保护的高度重视,亚热带地区形成了多种次生林,树种组成及结构趋于复杂。同时为了满足社会经济快速发展对木材生产的需求,人工林也成为了该地区主要森林类型。国内已开展了寒温带(如小兴安岭[9, 10])、温带(如长白山[4, 11])、暖温带(如太行山[12])、北亚热带(如神农架[13])、中亚热带(如天童山[14]、井冈山[15])和南亚热带(如鼎湖山[3, 7, 16, 17, 18])以及西北地区(如兴隆山[19]、贺兰山[20])乃至西南喀斯特地区(如茂兰[21])的主要森林类型凋落物量及其动态、凋落物分解及其影响因子、养分归还及其水文生态效应等方面的研究,揭示了我国不同气候带主要森林生态系统凋落物量、分解和养分归还动态及其影响因子等规律和机制。但是,由于森林凋落物的影响因素较多,许多研究结果仍缺乏可比性。此外,次生林与人工林在林龄、抚育方式等不同,凋落物特征也不同。对亚热带次生林和人工林凋落物量动态及其分解率和周转期的比较研究仍不多见。
湖南省长沙县大山冲森林公园经过50多年封山育林,保存了由不同演替阶段树种组成的多种次生林,还有营造的杉木人工林,这些林分的环境条件(母岩、土壤)相似,为比较不同次生林和人工林的生态功能过程提供了良好条件。本研究以3 种次生林:马尾松(Pinus massonana)+石栎(Lithocarpus glaber)针阔混交林(PM)、南酸枣(Choerospondia axillaris)落叶阔叶林(CA)、石栎+青冈(Cyclobalanopsis glauca)常绿阔叶林(LG)和杉木(Cunninghamia lanceolata)人工林(CL)为对象,比较不同森林类型及其优势树种凋落物量、组分动态及其分解率和周转期的差异,剖析次生林、人工林养分循环过程、维持地力能力的特征,揭示亚热带天然林保护和森林树种组成的差异对森林生态系统养分循环、碳流动的影响机制,为森林科学经营提供理论依据。
1 研究地概况研究地设在湖南省长沙县路口镇大山冲湖南省森林公园(东经113°17′—113°19′,北纬28°23′—28°24′),海拔高度在55—350 m之间,属于典型低山丘陵和亚热带大陆型季风湿润气候,年均温度16.7—17.6 ℃,极端高温40 ℃,极端低温-11 ℃,雨量充沛,相对湿度较大,年降雨量在1412—1559 mm之间,2010—2012 年月平均降水量和月平均气温如图 1所示。土壤以红壤为主,是由板岩和页岩发育而成,地带性植被为亚热带常绿阔叶林,属于湘中湘东山丘盆地栲(Castanopsis fargesii)林、马尾松林、毛竹(Phyllostachys heterocycla)林、油茶(Camellia oleifera)林及农田植被区——幕阜、连云山山地丘陵植被小区。

|
| 图1 研究地2012 年至2013 年月平均降水量和月平均气温 Fig.1 Monthly precipitation and air temperature in years of 2012 to 2013 |
2 009 年在杉木人工林和3 种具有代表性的次生林:马尾松+石栎针阔混交林、南酸枣落叶阔叶林、石栎+青冈常绿阔叶林分别建立面积为1 hm2的固定样地,各林分的固定样地均分为100 个10 m×10 m小样地,对样地内胸径大于1 cm的植物测定胸径、树高、枝下高和冠幅,记录植物种类,用Shannon-Wiener多样性指数计算各林分树种多样指数。各林分的基本特征如表 1所示。
| 森林类型 Forest type | 优势树种 Dominant species | 树种数 Species number | 密度 Density / (株/hm2) | 多样性指数 Diversity index | 平均胸径 Average DBH/cm | 平均树高 Average tree H/m | 海拔 Elevation/m | 坡向 Slope aspect | 坡度/(°) Slope |
| CL | 杉木Cunninghamia lanceolata | 1 | 625 | 0 | 23.5(4.0—35.0) | 19.5(4.5—27.0) | 223—258 | 东南 | 24 |
| PM | 整个林分 Whole stand | 19 | 2492 | 0.746 | 11.5(4.0—37.2) | 10.0(0.3—26.5) | 220—262 | 西南 | 15 |
| 马尾松Pinus massoniana | 816 | 17.1 | 13.5 | ||||||
| 石栎Lithocarpus glaber | 1340 | 8.0 | 8.0 | ||||||
| 南酸枣Choerospondias axillaries | 76 | 21.1 | 12.3 | ||||||
| 枫香Liquidambar formosana | 40 | 13.8 | 11.7 | ||||||
| 浙江柿Diospyros glaucifolia | 40 | 8.6 | 8.1 | ||||||
| CA | 整个林分 Whole stand | 32 | 1696 | 1.104 | 8.8(4.0—53.5) | 6.4(1.4—28.1) | 245—321 | 西 | 35 |
| 南酸枣Choerospondias axillaries | 184 | 27.5 | 15.9 | ||||||
| 檵木Loropetalum chinensis | 568 | 5.1 | 4.8 | ||||||
| 四川山矾Symplocos setchuensis | 308 | 6.3 | 5.1 | ||||||
| 毛豹皮樟Litsea coreana | 92 | 8.0 | 5.7 | ||||||
| 千年桐Vernicia fordii | 44 | 11.5 | 9.3 | ||||||
| LG | 整个林分 Whole stand | 23 | 1340 | 1.001 | 12.7(4.0—37.6) | 10.30(1.5—21.0) | 225—254 | 西北 | 22 |
| 石栎Lithocarpus glaber | 552 | 12.7 | 10.5 | ||||||
| 青冈Cyclobalanopsis Glauca | 188 | 13.9 | 11.4 | ||||||
| 马尾松Pinus massoniana | 140 | 17.6 | 14.0 | ||||||
| 檫木Sassafras tzumu | 68 | 19.9 | 14.1 | ||||||
| 南酸枣Choerospondias axillaries | 36 | 23.3 | 13.5 | ||||||
| 括号内的数据为树木胸径或树高的变化范围,多样性指数为Shannon-Wiener 指数; CL: 杉木人工林Cunninghamia lanceolata plantation forest; PM: 马尾松+石栎针阔混交林Pinus massoniana and Lithocarpus glaber mixed forest; CA: 南酸枣落叶阔叶林Choerospondias axillaries deciduous broadleaved forest; LG: 石栎+青冈常绿阔叶林L. glaber and Cyclobalanopsis glauca evergreen broadleaved forest | |||||||||
用凋落物收集器直接收集凋落物。用直径4.06 mm的圆形铁丝做成面积为1 m2圆形,然后将40目尼龙网缝制在圆形铁丝圈上,制作成锥形的收集袋,即收集器。2012 年7 月在4 种林分的固定样地内沿上坡、中坡、下坡分别安装3 个收集器,每个林分类型共9个作为重复。收集器用4根1 m长的PVC管固定,收集器最低端离地面60 cm。每月收集凋落物1次,到2013 年止,共收集了12 个月。每次收集到的各收集器的凋落物分别按树种分类,再按落叶、枯枝、落果、碎屑分类,在80 ℃恒温下烘至恒重后称重,4 种林分均取9 个收集器各树种不同组分的平均值作为该林分各组分、各树种各组分的月凋落物量。
2.3 林地凋落物层现存量的测定2011 年12 月下旬,在4 种林分固定样地沿上坡、中坡、下坡分别设置6 个小样方(1.0 m×1.0 m),根据凋落物层分层标准[22],按未分解层、半分解层和已分解层收集小样方内的凋落物,带回实验室,置于80 ℃下烘干至恒重后称重,4 种林分用6 个小样方的平均值估算各林地凋落物层各分解层的现存量及各分解层现存量占林地凋落物层现存总量的百分比。
2.4 凋落物周转期、分解率的估算凋落物周转期和分解率(或周转率)用以下的公式来计算[23, 24]:
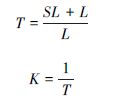
用SPSS10.0软件中单因素方差(One-way ANOVA)计算平均值和标准差,比较不同森林之间凋落物量差异和显著性,用回归分析方法分析林分凋落物量与林分密度、树种多样性指数、降水量、气温之间的相关性,用Excel软件绘制图表。
3 结果与分析 3.1 凋落物量及其组成从表 2可以看出,4种林分年凋落物总量在414.40—818.22 g m-2 a-1之间,大小顺序为PM>CA>LG>CL,其中CL与CA、LG之间差异显著(P<0.05),与PM之间差异极显著(P<0.01),但PM、CA、LG两两之间差异不显著(P>0.05)。凋落物主要由落叶(包括针叶和阔叶)、落枝、落果、碎屑(动物残体及其粪便、芽鳞、落花、树皮等统称)组分组成。不同林分凋落物的各组分量及其占林分凋落物总量百分比不同,CL为:落叶>枯枝>落果>碎屑,PM为:落叶>碎屑>枯枝>落果,CA和LG为:落叶>枯枝>碎屑>落果。4 种林分凋落物均以落叶为主,占林分凋落物总量的59.92%—66.62%,CL落叶最低,与PM、CA、LG差异显著(P<0.05),但PM、CA和LG两两之间差异不显著(P>0.05)。枯枝占林分凋落物总量的13.92%—25.25%,以CA最高,其次是CL,PM最低,但4种林分两两之间差异均不显著(P>0.05)。落果占林分凋落物总量的1.31%—10.09%,以CL落果及其占林分凋落物总量的百分比最高,其次是LG,PM、CA最低,且CL与PM、LG差异显著(P<0.05),与CA差异极显著(P<0.01),PM、CA、LG两两之间差异不显著(P>0.05)。碎屑占林分凋落物总量的1.78%—24.28%,以PM碎屑及其占林分凋落物总量的百分比最高,其次是CA、LG,CL最低,且CL与PM、CA、LG之间的差异,PM与CA之间的差异均极显著(P<0.01),与LG差异显著(P<0.05),但CA与LG差异不显著(P>0.05)。
| 林分类型 Forest type | 落叶 Leaf | 枯枝 Branch | 落果 Fruit | 碎屑 Chippings | 合计 Total |
| 杉木人工林 CL | 265.5±14.1 (64.1) Aa | 99.8±5.7 (24.1) Aa | 41.8±2.6 (10.1) Aa | 7.4±1.1 (1.8) Aa | 414.4±21.1 (100) Aa |
| 马尾松+石栎针阔混交林 PM | 490.3±22.0 (59.9) Ba | 114.0±13.6 (13.9) Aa | 15.3±1.14 (1.9) Bab | 198.7±8.9 (24.3) Bbd | 818.2±34.4 (100) Bb |
| 南酸枣落叶阔叶林 CA | 510.6±45.7 (64.3) Ba | 192.7±27.8 (24.3) Aa | 10.4±1.3 (1.3) Bb | 80.9±6.2 (10.2) Dc | 794.6±49.1 (100) Bab |
| 石栎+青冈常绿阔叶林 LG | 482.1±25.6 (66.6) Ba | 110.3±14.5 (15.2) Aa | 24.6±2.3 (3.4) Bab | 106.7±5.4 (14.7) Dcd | 723.7±30.7 (100) Bab |
| 括号内的数据为百分数(%),同列不同大写字母为差异显著(P<0.05),不同小写字母为差异极显著(P<0.01) | |||||
4种林分凋落物总量、落叶均呈现出明显的月变化特征,CL凋落物总量、落叶均呈双峰模式,峰值均在3 月和10 月,且10 月峰值高于3 月;CA凋落物总量、落叶也均呈双峰模式,第1峰值分别在4 月和3 月,第2个峰值均在10 月;PM凋落物总量、落叶均为不规则模式,出现多个峰值,其中凋落物总量在6 月和9 月明显增多,落叶在6、9、11 月也明显增多;LG凋落物总量为不规则模式,出现3个峰值,分别在3、6、9 月,落叶呈双峰模式,峰值在3 月和9 月(图 2)。

|
| 图2 4种林分凋落物总量及各组分的月动态 Fig.2 Monthly dynamics in total litter fall production and its components in the four forests |
枯枝、落果和碎屑的月变化特征不如凋落物总量、落叶的明显。其中CL枯枝有2个峰值,在4月和11月,CA枯枝峰值在12月,PM、LG枯枝峰值均在6 月。CL落果集中在2—4月和9—12月,PM在6月和11月,CA在7—8月,LG在9—8月。CL碎屑无明显的季节变化,PM生长季(4—9 月)碎屑较多,CA在4月较多,LG在3—10月较多(图 2)。此外,CL凋落物总量、落叶、枯枝的月变化较为平缓,月标准差较低(表 2),而PM、AC、LG月变化波动较大,月标准差较高,尤其是AC(表 2)。
3.3 优势树种凋落物总量、落叶及其月动态各林分的优势树种凋落物总量、落叶占其林分凋落物总量百分比表现为杉木凋落物总量、落叶占林分凋落物总量的百分比最高,分别为93.46%和61.25%,马尾松、石栎两者的凋落物总量、落叶分别占69.82%和54.52%,南酸枣分别占68.67%和53.46%,石栎、青冈两者的凋落物总量、落叶分别占51.20%和34.11%。4种林分优势树种凋落物总量、落叶占林分凋落物总量的百分比基本上随林分的树种多样性增加呈下降趋势。各优势树种落叶占相应树种凋落物总量的64.75%—81.21%之间,表明各优势树种凋落物量也是以落叶为主(表 3)。
| 项目Item | 杉木人 工林LC | 马尾松+石栎针阔混交林PM | 南酸枣落叶 阔叶林CA | 石栎+青冈常绿阔叶林LG | ||
| 杉木 C.lanceolata | 马尾松 P.massonana | 石栎 L.glaber | 南酸枣 C.axillaris | 石栎 L.glaber | 青冈 C.glauca | |
| 树种凋落物总量/(g m-2 a-1) Total litter fall quantity of tree species | 387.3±22.2 (93.5) | 385.5±21.0 (47.1) | 185.8±13.6 (22.7) | 545.7±50.9 (68.67) | 188.5±18.8 (26.0) | 182.1±19.0 (25.2) |
| 落叶Leaf/(g m-2 a-1) | 253.8±14.1 (61.3) | 313.0±18.6 (38.3) | 133.0±9.5 (16.3) | 424.8±48.1 (53.5) | 122.0±8.4 (16.7) | 124.8±18.2 (17.3) |
| 树种落叶占树种凋落物总量百分比/% The percentage of leaf to total litter fall of tree species | 65.5 | 81.2 | 71.6 | 77.8 | 64.8 | 68.6 |
同一优势树种凋落物总量、落叶的月动态基本一致,但不同树种模式不同,其中杉木、青冈均为双峰模式,第1个峰值均在3月,但第2个峰值杉木在10月,青冈在9月;南酸枣、石栎均为单峰模式,但两者峰值时间不同,南酸枣在9月,石栎在6月且在PM、LG中均表现一致;马尾松为不规则模式,但6月和9月凋落物量明显增多(图 3)。从图 2和图 3的比较可以看出,杉木、马尾松凋落物总量的月动态均与其林分凋落物总量的月动态基本一致,但南酸枣、青冈、石栎不一致。

|
| 图3 4种林分优势树种凋落物总量及其落叶量的月动态 Fig.3 Monthly dynamics in total litter fall and leaf of dominant species in the four forests |
从表 4可以看出,4种林分凋落物层现存总量依次为:LG>PM>CA>CL,且各林分类型之间差异极显著(P<0.01)。各分解层现存量及其占林分凋落物层现存总量的百分比因林分不同而异,尽管未分解层现存量依次为:PM>CA>CL>LG,且两两之间差异显著(P<0.05)或极显著(P<0.01),但CL占其林分凋落物层现存总量的百分比为最高,其次为CA,LG最低;半分解层现存量以CL最低,其次是PM、CA,LG最高,且4种林分两两间差异均极显著(P<0.01),但4种林分半分解层现存量占其林分凋落物层现存总量的百分比相当;LG已分解层现存量及其占林分凋落物层现存总量的百分比最高,其次是PM,CA最低,且4种林分已分解层现存量两两间差异均极显著(P<0.01)。不同分解层现存量占林分凋落物层现存总量的百分比均以半分解层为最低,LG已分解层现存量明显高于未分解层、半分解层现存量,PM已分解层现存量与未分解层现存量相近。
4 种林分凋落物分解率为0.31—0.45之间,周转期在2.2—3.2a之间,CA凋落物分解率最大(0.45),周转期最短(2.2a),其次是PM和LG,CL凋落物分解率最低,为0.31,周转期最长,为3.2a(表 4)。
| 林分类型 Forest type | 现存量The existing storage of litter/(g/m2) | 分解率 Decomposition rate | 周转期/a turnover period | |||
| 未分解层 Litter | 半分解层 Fermentation | 已分解层 Humus | 总计 Total | |||
| 杉木人工林 CL | 453±1.2 (50.8) Aa | 138±0.5 (15.5) Aa | 301±0.6 (33.7) Aa | 892±1.8 (100) Aa | 0.31 | 3.2 |
| 马尾松+石栎针阔混交林 PM | 484±0.9 (41.6) Bb | 198±0.4 (17.0) Bb | 484 ±2.2 (41.6) Bb | 1166±2.2 (100) Bb | 0.41 | 2.4 |
| 南酸枣落叶阔叶林 CA | 456±1.4 (50.0) Ca | 196±0.6 (21.5) Cc | 260±1.0 (28.5) Cc | 912±2.3 (100) Cc | 0.45 | 2.2 |
| 石栎+青冈常绿阔叶林 LG | 326±0.8 (27.1) Dc | 218±0.6 (18.1) Dd | 659±3.0 (54.7)Dd | 1203±3.6 (100)Dd | 0.40 | 2.5 |
| 括号内的数据为百分数(%),同列不同大写字母为差异显著(P<0.05),不同小写字母为差异极显著(P<0.01) | ||||||
同一气候条件下,森林类型是影响凋落物量的主要因素,凋落物量随树种组成、密度的不同而变化[25]。本研究中,CL凋落物总量、落叶显著或极显著低于3种次生林(PM、CA、LG),与现有的研究结果[26, 27]一致。与CL相比,3种次生林组成树种多样且林分密度大,次生林阔叶树的叶面积和质量均远大于CL的针叶,这是次生林凋落物量、落叶显著高于CL的原因。表明次生林组成树种复杂多样,凋落物量大,更有利于养分归还和地力的维持。此外,LG凋落物量、落叶低于PM、CA,但差异不显著,可能是由于:①3种次生林密度不同(表 1),其中PM密度最大(2493株/hm2),其次是CA(1696株/hm2),而LG(1340株/hm2),林分凋落物量、落叶与林分密度呈显著正相关(r=0.2877—0.3497,n=48,P<0.05);②3种次生林树种多样性不同,CA树种多样性最大(1.104),其次是LG,PM最低,林分凋落物量、落叶与林分树种多样性指数呈显著线性正相关(r=0.3033—0.3497,n=48,P<0.05)。表明林分凋落物量、落叶随林分密度、树种多样性增加而增大,林分密度和树种多样性是导致林分凋落物量、落叶差异的主要原因。此外,同一地区不同林分组成树种的生物学和生态学特性的差异,也是林分凋落物量、落叶差异的重要影响因素。
森林主要树种的生物学特性不仅制约着林分凋落物量,还影响到凋落方式和凋落器官的比例[28]。本研究中,不同林分凋落物各组分的量及其占林分凋落物量的百分比均不同,但4种林分凋落物量均以落叶为主体,落叶占林分凋落物量的59.9%—66.6%之间,与郭剑芬等[26]研究结果(62%—69%)接近,表明落叶在林分凋落物量中占据关键地位。研究表明,杉木针叶在秋季少雨月份(10—12月)枯黄,之后连同小枝一起脱落,落叶与枯枝之间存在极显著的正相关,而阔叶树则在叶片凋落前,离层细胞间的中层粘液化、分解,使叶柄自离层处折断,叶片脱落,枝条则在以后凋落,其叶、枝的凋落量不存在任何相关[28],导致枯枝也成为CL凋落物的主要部分,占林分凋落物总量的24.1%,明显高于PM、LG。由于常绿针叶树用于繁殖的器官资源分配较大,阔叶树的养分主要用于营养器官的生长,其果实小、质量轻[9],导致CL的落果量及其占林分凋落物总量的百分比显著高于PM、CA、LG。碎屑也是凋落物中不可忽视的部分,PM、CA、LG极显著高于CL,PM也极显著或显著高于CA、LG,表明次生林中鸟类、昆虫等动物比较丰富,尤其是PM,可能是由于马尾松抗虫害能力弱,易受松毛虫危害,虫粪多是马尾松林凋落物组成的一个显著特点[29]。相关分析结果表明,林分枯枝、落果、碎屑与林分密度、树种多样性指数均不存在显著相关(P>0.05),表明林分密度及其树种多样性不是影响林分枯枝、落果、碎屑的主要因素。
林分凋落物特征是林分树种共同形成的,林分优势树种的改变将会影响林分凋落物特征[30],而各树种生物量最能直观地反映该树种对林分结构的贡献[31]。本研究中,各林分优势树种凋落物量、落叶分别占其林分凋落物量、落叶的51.2%和34.1%以上,但随着林分树种多样性的提高呈下降趋势。表明无论是次生林还是杉木人工林的凋落物量、落叶均以其优势树种凋落物量、落叶为主,但优势树种对林分凋落物量的贡献随林分树种数量的增加而下降。表明次生林组成树种多样复杂,明显改变了林分凋落物组成的特征。
4.2 不同林分凋落物量峰值的形成森林凋落物量随气候因子的季节变化而呈现明显的季节变化。本研究中,落叶是林分、各树种凋落物量主体,对林分凋落物量贡献了1/2以上,无论是林分凋落物量的月动态还是各林分优势树种凋落物量的月动态都主要受控于落叶的月动态。
森林凋落物量的月动态模式有单峰型、双峰型或不规则型,主要取决于林分树种的生物学和生态学特性以及气候因素的综合影响[17],亚热带不同地区常绿阔叶林凋落物量季节动态并不一致,多数是双峰,少数是单峰,与林分树种组成有关[17, 18]。本研究中,杉木、马尾松凋落物量的月动态与其林分凋落物量的月动态基本一致,而南酸枣、青冈、石栎与其林分没有一致的变化趋势。CL、杉木凋落物量、落叶均呈双峰型,是由于CL是由杉木单一树种组成,因此CL凋落物量、落叶的月动态是由杉木凋落物量、落叶所控制。研究表明,3—4月是常绿针叶树换叶季节,此时大量老叶凋落形成第1个峰值,杉木针叶在少雨秋季枯黄,连同小枝一起脱落而出现第2个峰值[28]。PM、马尾松凋落物量、落叶的月动态为不规则型,是由于PM仍以马尾松为绝对优势树种,林分凋落物量的月动态仍取决于马尾松凋落物量的动态,3—4月是马尾松换叶季节,此时有较多老叶脱落,出现第1个峰值,6月是马尾松针叶凋落高峰而形成第2个峰值,秋季(9—11月)少雨干旱促使马尾松针叶大量脱落以减少水分的消耗导致第3个峰值出现。CA凋落物量、落叶呈双峰型,而南酸枣凋落物量、落叶呈单峰型,是由于CA中除了南酸枣、檵木、千年桐等落叶树种外,还有四川山矾、毛豹皮樟等常绿阔叶树,尽管南酸枣1—4月萌发新芽叶,此时南酸枣凋落物量几乎为零,但3—4月是常绿阔叶树换叶,老叶脱落[18]季节,因此CA凋落物量仍然出现一个较小峰值;秋冬季节(9—12月)是南酸枣等落叶树种落叶高峰[27, 32],也是常绿阔叶树部分叶子完成使命后开始脱落,因此9—11月CA、南酸枣出现落叶峰值。LG凋落物量为不规则型,落叶呈双峰型,而青冈凋落物量、落叶呈双峰型,石栎呈单峰型,是由于大多常绿阔叶树在春夏(3—5月)期间新叶萌发,生长旺盛,促使衰老的叶子脱落,此时凋落物明显增多而出现第1个峰值,秋季(9—11月)干旱少雨、温度下降,大多常绿阔叶树果实成熟脱落而结束一个繁殖周期,部分叶子完成使命开始脱落而出现第2个高峰,因此大多常绿阔叶树凋落物量、落叶月动态为双峰型。也有少数常绿阔叶树凋落物量、落叶仅在5—6月新叶长成后出现一个高峰值,在秋冬季节没有明显的变化,不出现第2个峰值。表明青冈属于大多常绿阔叶树凋落物量、落叶月动态为双峰型,而石栎属于少数常绿阔叶树凋落物量、落叶月动态为单峰型。此外,由于6月强度大的降雨频率高,枯枝凋落量明显增加,导致LG凋落物量在6月出现峰值。相关分析结果表明,4种林分月凋落物量、月落叶量与月降水量之间不存在显著相关(相关系数分别为0.065和-0.056,P>0.05,n=48),与月平均温度之间呈显著的线性正相关(相关系数分别为0.3086和0.2932,P<0.05,n=48),表明林分凋落物量、落叶的月动态与林分树种生物学特性和当地气温的季节变化密切相关。
4.3 不同林分凋落物层现存量和周转期的差异林地凋落物层是凋落物逐渐积累而形成,并处于不断分解和聚积的动态变化中,林地凋落物层的积累量随林分密度增大而增加[24],林地凋落物层的数量和组成反映了凋落物的产生和分解过程[30]。本研究中,4 种林分凋落物层现存量变化趋势为:LG>PM>CA>CL,与郑路等[22]的研究结果基本一致。由于CL凋落物难分解,PM、CA、LG凋落物易于分解,而CA在9—11月为落叶高峰,在12月测定时,林地未分解的凋落物仍较多,导致CL、CA未分解层现存量占林分凋落物层现存量的百分比较高于PM、LG,而半分解层和已分解层所占百分比较低于PM、LG。相关分析结果表明,凋落物层现存量、各分解层现存量与林分树种多样性指数、密度呈线性正相关,但未达到显著水平(P>0.05),表明林分树种数量、密度不是林地调落物层现存量主要影响因子。
本研究中,CL凋落物分解率最低(0.31),周转期最长(3.2a),与广西不同密度湿地松凋落物的平均分解速率和周转期[24]、浙江天童不同演替阶段常绿阔叶林凋落物周转期[14]相近。CA凋落物分解率最高(0.45),周转期最短(2.2 a),导致CA凋落物层现存量,尤其是已分解层现存量显著低于LG、PM,而LG凋落物层现存量最高,可能与其年凋落物量较多,已分解层现存量明显增加有关。理论上CL凋落物层现存量应较高,但CL树种单一,林分密度低,又是针叶林,导致CL的凋落物层现存量最低。相关分析表明,林分凋落物的周转期与林分树种多样性指数呈显著的线性负相关(P<0.05),与林分密度呈线性负相关,但未达到显著水平(P>0.05),表明林分树种、密度增加,不仅增加了凋落物量,而且改变了凋落物的组成,加快凋落物的分解和周转。
总之,次生林比杉木人工林年凋落量大,且分解率高,周转期短,群落结构稳定,能更好地进行养分归还,具有良好维持土壤肥力的能力。因此保护次生林,促进其向顶极常绿阔叶林群落演替,对提高整个森林生态系统的碳汇功能和生态恢复有深远意义。
| [1] | 林波,刘庆,吴彦,何海.森林凋落物研究进展.生态学杂志,2004,23(1):60-64. |
| [2] | Chapin F S III, Matson P A, Mooney H A. Principles of Terrestrial Ecosystem Ecology. New York: Springer-Verlag, 2002. |
| [3] | Zhou GY, Guan L L, Wei X H, Zhang D Q, Zhang Q M, Yan J H, Wen D Z, Liu J X, Liu S G, Huang Z L, Kong G H, Mo J M, Yu Q F. Litterfall production along successional and altitudinal gradients of subtropical monsoon evergreen broadleaved forests in Guangdong, China. Plant Ecology, 2007, 188(1):77-89. |
| [4] | 刘颖,韩士杰,林鹿.长白山四种森林类型凋落物动态特征.生态学杂志,2009,28(1):7-11. |
| [5] | Janzen H H. Carbon cycling in earth systems: A soil science perspective. Agriculture, Ecosystems and Environment, 2004, 104(3): 399-417. |
| [6] | Liski J, Nissinen A, Erhard M, Taskine O. Climate effects on litter decomposition from arctic tundra to tropical rainforest. Global Change Biology, 2003, 9(4): 575-584. |
| [7] | 邓琦,刘世忠,刘菊秀,孟泽,张德强.南亚热带森林凋落物对土壤呼吸的贡献及其影响因素.地球科学进展,2007,22(9):976-986. |
| [8] | 吴雅琼,刘国华,傅伯杰,郭玉华.中国森林生态系统土壤CO2释放分布规律及其影响因子.生态学报,2007,27(5):2126-2135. |
| [9] | 侯玲玲,毛子军,孙涛,宋元.小兴安岭十种典型森林群落凋落物生物量及其动态变化.生态学报,2013,33(6):1994-2002. |
| [10] | 陈金玲,金光泽,赵凤霞.小兴安岭典型阔叶红松林不同演替阶段凋落物分解及养分变化.应用生态学报,2010,21(9):2209-2216. |
| [11] | 郭忠玲,郑金萍,马元丹,李庆康,于贵瑞,韩士杰,范春楠,刘万德.长白山各植被带主要树种凋落物分解速率及模型模拟的试验研究.生态学报,2006,26(4):1037-1046. |
| [12] | 赵勇,吴明作,樊巍,高喜荣.太行山针阔叶森林凋落物分解及养分归还比较.自然资源学报,2009,24(9):1616-1624. |
| [13] | 刘蕾,申国珍,陈芳清,罗璐,谢宗强,喻杰.神农架海拔梯度上4种典型森林凋落物现存量及其养分循环动态.生态学报,2012,32(7):2142-2149. |
| [14] | 张庆费,徐绒娣.浙江天童常绿阔叶林演替过程的凋落物现存量.生态学杂志,1999,18(2):17-21. |
| [15] | 李海涛,于贵瑞,李家永,梁涛,陈永瑞.井冈山森林凋落物分解动态及磷钾释放速率.应用生态学报,2007,18(2):233-240. |
| [16] | 官丽莉,周国逸,张德强,刘菊秀,张倩媚.鼎湖山南亚热带常绿阔叶林凋落物量20年动态研究.植物生态学报,2004,28(4):449-456. |
| [17] | 翁轰,李志安,屠梦照,姚文华.鼎湖山森林凋落物量及营养元素含量研究.植物生态学与地植物学报,1993,17(4):299-304. |
| [18] | 屠梦照,姚文华,翁轰,李志安.鼎湖山南亚热带常绿阔叶林凋落物的特征.土壤学报,1993,30(1):34-41. |
| [19] | 魏强,凌雷,张广忠,闫沛斌,陶继新,柴春山,薛睿.甘肃兴隆山主要森林类型凋落物累积量及持水特性.应用生态学报,2011,22(10):2589-2598. |
| [20] | 赵晓春,刘建军,任军辉,刘斌,母吉君.贺兰山4种典型森林类型凋落物持水性能研究.水土保持研究,2011,18(2):107-111. |
| [21] | 俞国松,王世杰,容丽,冉景丞.茂兰喀斯特森林主要演替群落的凋落物动态.植物生态学报,2011,35(10):1019-1028. |
| [22] | 郑路,卢立华.我国森林地表凋落物现存量及养分特征.西北林学院学报,2012,27(1):63-69. |
| [23] | 蒋有绪.川西亚高山冷杉林枯枝落叶层的群落学作用.植物生态学与地植物学丛刊,1981,5(2):8-998. |
| [24] | 项文化,田大伦,蔡宝玉,钟建德.不同密度湿地松林凋落物量及养分特性的研究.林业科学,1997,33(S.2):175-180. |
| [25] | Facelli J M, Pickett S T A. Plant litter: its dynamics and effects on plant community structure. The Botanical Review, 1991, 57(1): 1-32. |
| [26] | 郭剑芬,陈光水,钱伟,杨少红,杨玉盛,郑群瑞.万木林自然保护区2种天然林及杉木人工林凋落量及养分归还.生态学报,2006,26(12):4091-4098. |
| [27] | Lian Y W, Zhang Q S. Conversion of a natural broad-leafed evergreen forest into pure and mixed plantation forests in a subtropical area: effects on nutrient cycling. Canadian Journal of Forest Research, 1998, 28(10): 1518-1529. |
| [28] | 温远光,韦炳二,黎洁娟.亚热带森林凋落物产量及动态的研究.林业科学,1989,25(6):542-548. |
| [29] | 杨会侠,汪思龙,范冰,张伟东.不同林龄马尾松人工林年凋落量与养分归还动态.生态学杂志,2010,29(12): 2334-2340. |
| [30] | 胡灵芝,陈德良,朱慧玲,陈永华,丁炳扬.百山祖常绿阔叶林凋落物凋落节律及组成.浙江大学学报:农业与生命科学版,2011,37(5):533-539. |
| [31] | 方炜,彭少麟.常绿阔叶林中种群在群落中地位测度指标的探讨.生态学报,1996,16(1):111-112. |
| [32] | 樊后保,李燕燕,孙新,林德喜.马尾松纯林及其与阔叶树混交林的凋落量与养分通量.应用与环境生物学报,2005,11(5):521-527. |