 2015, Vol. 35
2015, Vol. 35文章信息
- 孟莹莹, 周莉, 周旺明, 于大炮, 包也, 王晓雨, 郭焱, 代力民
- MENG Yingying, ZHOU Li, ZHOU Wangming, YU Dapao, BAO Ye, WANG Xiaoyu, GUO Yan, DAI Limin
- 长白山风倒区植被恢复26年后物种多样性变化特征
- Characteristics of plant species diversity in a windthrow area on Changbai Mountain after 26 years of natural recovery
- 生态学报, 2015, 35(1): 142-149
- Acta Ecologica Sinica, 2015, 35(1): 142-149
- http://dx.doi.org/10.5846/stxb201405301109
-
文章历史
- 收稿日期:2014-05-30
- 修订日期:2014-11-06






2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
生物多样性是人类赖以生存的条件,也是经济得以持续发展的基础[1]。物种多样性是生物多样性的重要组成部分,是衡量某一地区生物多样性状况和一个群落结构和功能复杂性的重要指标之一[2]。植物群落物种多样性作为生态系统多样性最直接和最易于观察研究的一个层次,一直受到专家学者的重视[3]。物种多样性受许多因素的影响[4, 5, 6, 7, 8],而干扰被广泛地认为是影响物种多样性的主要因素之一[9]。风干扰是森林生态系统自然干扰中较常见的一种[10],通过研究风干扰对植物多样性的影响,不仅可以全面评价生态系统植被恢复程度,而且有利于促进干扰后生态系统的更新恢复。因此,研究风倒区植物多样性特征具有重要的意义。
长白山位于吉林省的东部,是我国东北地区海拔最高的山脉。20世纪80年代,长白山西坡和西南坡遭受了罕见的台风袭击,使得保护区植被遭到极大破坏。经过近30a的自然恢复,与原始植被群落相比,其风倒区的植被群落类型发生了变化。但以往有关长白山风干扰后植被恢复多样性的研究多是针对乔木层物种多样性变化[11],而系统开展植被恢复过程中乔木层、灌木层和草本层的植被多样性变化研究还较少。因此,本文通过对长白山自然保护区风灾迹地3种典型森林类型的乔木、灌木和草本恢复群落的多样性进行全面的调查研究,旨在揭示长白山风灾迹地植被群落恢复状况,认清群落的结构、类型及群落的演替趋势,以期为风灾迹地的植被恢复和受损生态系统的恢复提供理论根据。
1 研究区域长白山自然保护区风灾区位于北纬41°52′ 40″—42°01′10″,东经 127°53′37″—128°02′00″,南北长为15.75km,东西宽为11.75km,形成从海拔 1050—1700m的阔叶红松林、云冷杉林至岳桦林的风倒带[12]。3个林带的土壤分别为暗棕色森林土、棕色针叶林土和亚高山草甸土[13]。年平均气温3.3℃,最热月平均气温19.3℃,最冷月平均气温为-16.2℃,年降水量800mm,生长季(5—9月)降水量616mm,占全年降水量的76.9%[14]。风灾区内光照强度增大,晴天比林内高100—200lx;风灾区内温差增大,晴天比林内气温高1—2℃,阴天比林内气温约低1℃[12]。
2 研究方法 2.1 植被调查2012年9月对长白山西坡风倒区进行植被调查,采用典型的群落取样方法[15, 16],按照海拔由低到高分别调查了3种典型的森林类型(阔叶红松林,云冷杉林和岳桦林)的乔木、灌木和草本群落(表 1),并选择立地条件基本一致的未受风干扰影响的样地作为对照。共调查样地22块(风倒区与对照区各11块),其中阔叶红松林和云冷杉林风倒区与对照区各4块,岳桦林风倒区与对照区各3块。调查面积乔木样方为30m×30m,灌木样方为5m×5m;草本样方为1m×1m。将每块乔木样方划分成9个10m×10m的小样方进行调查,在每块乔木样方内,按对角线设置3—5块灌木和草本样方。调查项目主要包括:乔木树种的胸径和树高;灌木和草本植物的高度、盖度和株数;同时记录各样方的经纬度、海拔、坡度、坡向和坡位。样地基本概况见表 1。
| 森林类型 Forest Types | 海拔 Altitude/m | 干扰类型 Disturbance types | 物种种类Species | 主要植被类型Main Vegetation Type | ||||
| 乔木层 | 灌木层 | 草本层 | 乔木层 Tree layer | 灌木层 Shrub layer | 草本层 Herb layer | |||
| 阔叶红松林 | 1100—1300 | 对照区 | 21 | 9 | 20 | 臭冷杉Abies nephrolepis | 刺五加Acanthopanax senticosus | 山茄子Scopolia acutangula |
| BKPF | 红松Pinus koraiensis | 小楷槭Acer tschonoskii | 美汉草Meehania uriticifolia | |||||
| 紫椴Tilia amurensis | 花楷槭Acer ukurunduense | 三花拉拉藤Galium triflorum Michx. | ||||||
| 蒙古栎Quercus mongolica | 黄花忍冬Lonicera chrysantha | 东北羊角芹AegopodiumsalpestreLedeb | ||||||
| 风倒区 | 24 | 11 | 31 | 白桦B.platyphylla | 花楷槭Acer ukurunduense | 二叶舞鹤草Maianthemum bifolium | ||
| 落叶松Larix olgensis | 狗枣猕猴桃Actinidia kolomikta | 山酢浆草Oxalis acetosella | ||||||
| 臭冷杉Abies nephrolepis | 青楷槭Acer tegmentosum | 卵果蕨Phegopteris polypodioides | ||||||
| 山杨Populus davidiana | ||||||||
| 云冷杉林 | 1300—1500 | 对照区 | 12 | 9 | 17 | 臭冷杉Abies nephrolepis | 笃斯越橘Vaccinium uliginosum | 小叶章Calamagrostis angustifolia |
| SFF | 落叶松Larix olgensis | 蓝靛果忍冬Lonicera edulis | 二叶舞鹤草Maianthemum bifolium | |||||
| 鱼鳞云杉Picea jezoensis | 东北羊角芹AegopodiumsalpestreLedeb | |||||||
| 风倒区 | 5 | 4 | 25 | 臭冷杉Abies nephrolepis | 库页悬钩子Rubus matsumuranus | 小叶章Calamagrostis angustifolia | ||
| 岳桦Betula ermanii | 深山蔷薇Rosa marretii | |||||||
| 鱼鳞云杉Picea jezoensis | 蓝靛果忍冬Lonicera edulis | |||||||
| 岳桦林 | 1500—1700 | 对照区 | 1 | 2 | 10 | 岳桦Betula ermanii | 蓝靛果忍冬Lonicera edulis | 小叶章Calamagrostis angustifolia |
| EBF | 落叶松Larix olgensis | 库页悬钩子Rubus matsumuranus | 贝加尔野豌豆 baicalensis (Turcz.)B Fedtsch | |||||
| 风倒区 | 4 | 5 | 29 | 岳桦Betula ermanii | 笃斯越橘Vaccinium uliginosum | 小叶章Calamagrostis angustifolia | ||
| 蓝靛果忍冬Lonicera edulis | ||||||||
| 库页悬钩子Rubus matsumuranus | ||||||||
| 金老梅Potentilla fruticosa | ||||||||
物种多样性指标的测定方法参考马克平等[17]、郝占庆等[18]。
Patrick物种丰富度指数:
R=S
Shannon-Wiener多样性指数:
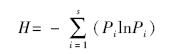
Simpson 优势度指数:
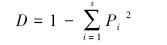
Pielou均匀度指数:
E=H/lnS
式中,S为物种数,Pi为第i个种的个体数占所有种株数的比例。
重要值的计算:
乔木树种的重要值=(相对多度+相对显著度+相对频度)/3
灌木树种的重要值=(相对多度+相对显著度+相对频度)/3
草本物种的重要值=(相对多度+相对频度+相对盖度)/3
2.3 数据统计与分析用Excel 2007和SPSS16.0软件分别进行数据处理和统计分析,采用t检验法检验两个变量之间的差异显著性(α=0.05),origin8.5软件绘图。
3 结果与分析 3.1 长白山风倒区3种主要森林类型乔木树种多样性恢复特征通过对比风倒区和对照区主要树种重要值可知,风倒区恢复近30a,原阔叶红松林风倒迹地已发展成以白桦和山杨为主的杨桦次生林(表 2);相对于对照区而言,云冷杉林风倒区树种组成比较简单,只有6种树(表 3);而对照区中岳桦树种重要值是100,表明对照区是岳桦纯林,风倒区中乔木有4种,说明风倒区乔木群落组成较对照区复杂(表 4)。
| 阔叶红松林对照区 Control area of BKPF | 阔叶红松林风倒区 Windthrow area of BKPF | ||
| 树种 Tree species | 重要值 Importance value | 树种 | 重要值 |
| 假色槭 | 16.51±8.92 | 白桦 | 25.29±9.23 |
| 红松 | 14.68±5.54 | 山杨 | 22.64±12.65 |
| 鱼鳞云杉 | 9.91±4.11 | 色木槭 | 12.01±0.74 |
| 紫椴 | 9.24±1.65 | 小叶杨 | 9.43±4.13 |
| 蒙古栎 | 8.82±5.24 | 臭冷杉 | 8.17±2.90 |
| 簇毛槭 | 8.72±4.13 | 青楷槭 | 6.83±2.62 |
| 臭冷杉 | 5.13±4.44 | 大叶杨 | 6.55±2.27 |
| 色木槭 | 5.10±2.52 | 稠李子 | 4.98±3.04 |
| 花楷槭 | 4.56±0.99 | 紫椴 | 3.62±2.25 |
| 青楷槭 | 4.28±3.83 | 枫桦 | 3.45±1.21 |
| 云冷杉林对照区 Control Area of SFF | 云冷杉林风倒区 Windthrow Area of SFF | ||
| 树种 | 重要值 | 树种 | 重要值 |
| 鱼鳞云杉 | 38.78±12.00 | 鱼鳞云杉 | 55.93±4.70 |
| 臭松 | 19.54±1.40 | 臭松 | 24.28±8.74 |
| 色木 | 14.28±5.19 | 落叶松 | 9.37±5.46 |
| 岳桦 | 12.47±7.00 | 岳桦 | 5.91±4.23 |
| 红松 | 6.24±5.52 | 枫桦 | 3.34±2.78 |
| 水曲柳 | 3.42±1.35 | 白桦 | 1.17±1.03 |
| 白桦 | 2.32±1.03 | ||
| 小叶杨 | 1.09±0.49 | ||
| 杨树 | 1.05±0.61 | ||
| 枫桦 | 0.62±0.08 | ||
| 岳桦林对照区 Control Area of EBF | 岳桦林风倒区 Windthrow Area of EBF | ||
| 树种 | 重要值 | 树种 | 重要值 |
| 岳桦 | 100±0.00 | 岳桦 | 55.39±13.30 |
| 落叶松 | 39.18±8.35 | ||
| 白桦 | 3.48±1.03 | ||
| 臭松 | 1.95±0.38 | ||
比较长白山风倒区与对照区3种主要林型乔木树种多样性指数(图 1),云冷杉林对照区的物种丰富度显著高于风倒区,而阔叶红松林和岳桦林对照区的物种丰富度均小于风倒区,但差异不显著。阔叶红松林对照区与风倒区之间,Shannon-Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数差异均不显著。而云冷杉林对照区的Shannon-Wiener多样性指数和Simpson优势度指数均显著高于风倒区,Pielou均匀度指数差异不显著;岳桦林对照区的Shannon-Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数均显著高于风倒区。随着海拔的升高,对照区和风倒区乔木物种丰富度和α多样性均呈逐渐降低的趋势。

|
| 图 1 3种林型对照区与风倒区乔木树种物种丰富度和α多样性指数对比 Fig. 1 The contrast of species richness and α diversity index of tree layer in windthrow area and control area of three forest types |
阔叶红松林对照区灌木群落物种丰富度显著高于风倒区,岳桦林对照区灌木群落物种丰富度显著低于风倒区,云冷杉林差异不显著。阔叶红松林灌木群落α多样性对照区与风倒区之间差异均不显著;云冷杉林灌木群落Shannon-Wiener多样性指数和Simpson优势度指数对照区均显著低于风倒区;岳桦林灌木群落只有Shannon-Wiener多样性指数是对照区显著低于风倒区,其它两指标差异不显著(图 2)。

|
| 图 2 3种林型对照区与风倒区灌木物种丰富度和α多样性指数对比 Fig. 2 The contrast of species richness and α diversity index of shrub layer in windthrow area and control area of three forest types |
3种林型中草本群落对照区与风倒区之间物种丰富度均没有显著差异。阔叶红松林中草本群落Shannon-Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数均是对照区显著高于风倒区;云冷杉林中草本群落对照区与风倒区之间α多样性差异不显著;岳桦林中草本群落只有Simpson优势度指数对照区显著高于风倒区(图 3)。

|
| 图 3 3种林型对照区与风倒区草本植物物种丰富度和α多样性指数对比 Fig. 3 The contrast of species richness and α diversity index of herb layer in windthrow area and control area of three forest types |
干扰是生态系统多样性改变的重要原因[19],干扰自身只能减少或维持群落的物种多样性而不能增加群落的物种多样性[20]。本研究中风干扰后,其乔木层的物种丰富度和α多样性均没有显著变化,但优势树种的组成有极大的不同:对照区阔叶红松林以红松、紫椴等珍贵硬木为主,风倒区阔叶红松林以阳性树种白桦、山杨为主。有研究表明,皆伐后阔叶红松林的演替过程为:白桦作为先锋树种首先占据主要位置,然后被阔叶树种群取代,最后将达到以红松为主要树种的顶级群落[21],本研究表明风干扰26年后阔叶红松林风灾迹地目前属于演替的第一阶段,要演替到干扰前的状态仍需较长时间。云冷杉林和岳桦林风灾迹地乔木层的物种丰富度和α多样性指数均降低,且更新演替较差,更新树种类数量较少,但与对照区的树种组成差别不大,云冷杉林对照区与风倒区均是以鱼鳞云杉、臭松等针叶树种和阔叶树岳桦为主,岳桦林风倒区则形成以散生于杂草丛中的岳桦单优种群。乔木层物种丰富度、Shannon-Wiener多样性指数和 Simpson优势度指数的排序结果为:阔叶红松林>云冷杉林>岳桦林,表明干扰26年后,就乔木层而言,阔叶红松林更新最快,云冷杉林其次,岳桦林演替最慢。云冷杉林和岳桦林乔木层物种多样性较灾前显著降低,要想演替到灾前水平需要更长时间或有效的人为措施促进植被恢复。
风干扰后,阔叶红松林风灾迹地灌木层物种丰富度和α多样性均降低;云冷杉林风灾迹地灌木层物种丰富度降低了,α多样性升高了;岳桦林风灾迹地灌木层二者均升高了。灌木层Shannon-Wiener多样性指数和 Simpson优势度指数的排序结果为:云冷杉林>岳桦林>阔叶红松林。阔叶红松林风灾迹地草本层Simpson指数和均匀度指数显著降低;云冷杉林草本层丰富度和物种多样性均降低了,且除了物种丰富度外差异均达到显著水平;岳桦林物种丰富度和α多样性指数没有显著变化。干扰后灌木层和草本层多样性的这种变化,主要是由于阔叶红松林风灾迹地乔木群落更新较好,森林郁闭度较大,林下光照较差,抑制林下灌木和草本植被的生长;而云冷杉林风干扰后更新较差,郁闭度较小,为灌木和草本植被的生长提供了充足的阳光,但是由于小叶章具有紧密丛生的特点,使其具有竞争优势,不利于其它草本植物的生长,导致云冷杉林风灾迹地草本植被以小叶章为主,多样性较低。岳桦林受灾程度与云冷杉林相近,但是由于岳桦林所处的高海拔地区,物种多样性本来就较低[22],所以风倒后差异不显著。
当一个群落演替到顶级阶段时,其构成群落的种类之间往往达到某种协调[23],风倒区植被恢复26a后,物种组成和多样性与对照区还存在较大差异,26a相对于漫长的森林演替过程还只能说是森林演替的初期阶段,全面揭示从风干扰后开始的次生林恢复演替到地带性顶级群落的整个恢复过程中物种多样性的变化规律还有待进一步深入研究。
5 结论风干扰后经过26a的自然恢复,与对照区相比,阔叶红松林风倒区乔木层物种丰富度和α多样性差异已不显著,然而云冷杉林和岳桦林均显著降低;阔叶红松林灌木层物种丰富度和α多样性均呈降低趋势,且丰富度显著降低,云冷杉林和岳桦林α多样性显著升高;阔叶红松林和云冷杉林草本层α多样性显著降低,岳桦林差异不显著。表明阔叶红松林乔木层更新恢复的最好,灌木层和草本层更新较差;云冷杉林和岳桦林均是乔木层更新较差,灌木层更新较好;但是云冷杉林草本层物种较单一,岳桦林变化不大。
| [1] | 樊正球, 陈鹭真, 李振基. 人为干扰对生物多样性的影响. 中国生态农业学报, 2001, 9(2): 31-34. |
| [2] | 谢晋阳, 陈灵芝. 暖温带落叶阔叶林的物种多样性特征. 生态学报, 1994, 14(4): 337-344. |
| [3] | 李秋年. 高寒草甸退化草地植物群落结构特征及物种多样性的初步分析. 青海环境, 2004, 14(1): 30-33. |
| [4] | Palmer M W. Variation in species richness: towards a unification of hypotheses. Folia Geobotanica & Phytotaxonomica, 1994, 29(4): 511-530. |
| [5] | 于立忠, 朱教君, 孔祥文, 胡万良, 谭学仁. 人为干扰(间伐)对红松人工林林下植物多样性的影响. 生态学报, 2006, 26(11): 3757-3764. |
| [6] | 孙家宝, 张海林, 胡海清. 火干扰强度对兴安落叶松林物种组成及多样性的影响. 森林工程, 2009, 25(6): 1-5. |
| [7] | 吕浩荣, 刘颂颂, 叶永昌, 朱剑云, 莫罗坚, 陈红跃. 人为干扰对风水林群落树种组成及多样性的影响. 生态学杂志, 2009, 28(4): 613-619. |
| [8] | 李新彬, 王襄平, 罗菊春, 牟长城, 林英建, 夏新莉. 干扰对长白山北坡次生林群落结构和树种多样性的影响. 林业科学, 2006, 42(2): 105-110. |
| [9] | Mackey R L, Currie D J. The diversity-disturbance relationship: Is it generally strong and peaked?. Ecology, 2001, 82(12): 3479-3492. |
| [10] | 陈利顶, 傅伯杰. 干扰的类型、特征及其生态学意义. 生态学报, 2000, 20(4): 581-586. |
| [11] | 郭利平, 姬兰柱, 张伟东, 张悦, 薛俊刚. 长白山西坡风灾区森林恢复状况. 应用生态学报, 2010, 21(6): 1381-1388. |
| [12] | 赵晓飞, 牛丽君, 陈庆红, 张植, 栾艳新. 长白山自然保护区风灾干扰区生态系统的恢复与重建. 东北林业大学学报, 2004, 32(4): 38-40. |
| [13] | 薛俊刚. 吉林长白山国家级自然保护区风灾区植被恢复情况调查. 国土与自然资源研究, 2009, (1): 95-96. |
| [14] | 侯向阳, 韩进轩. 长白山红松阔叶混交林风灾迹地的更新与恢复. 林业科学, 1996, 32(5): 419-425. |
| [15] | 代力民, 谷会岩, 邵国凡, 王庆礼. 中国长白山阔叶红松林. 沈阳: 辽宁科学技术出版社, 2004. |
| [16] | 代力民, 邵国凡. 森林经营决策-理论与实践. 沈阳: 辽宁科学技术出版社, 2005. |
| [17] | 马克平, 刘玉明. 生物群落多样性的测度方法Ⅰα多样性的测度方法(下). 生物多样性, 1994, 2(4): 231-239. |
| [18] | 郝占庆, 郭水良, 曹同. 长白山植物多样性及其格局. 沈阳: 辽宁科学技术出版社, 2002. |
| [19] | 段仁燕. 干扰对生态系统中生物多样性的影响. 安徽农学通报, 2008, 14(9): 57-58. |
| [20] | 贺金生, 陈伟烈, 江明喜, 金义兴, 胡东, 路鹏. 长江三峡地区退化生态系统植物群落物种多样性特征. 生态学报, 1998, 18(4): 399-407. |
| [21] | Chen X W, Li B L, Lin Z S. The acceleration of succession for the restoration of the mixed-broadleaved Korean pine forests in Northeast China. Forest Ecology and Management, 2003, 177(1/3): 503-514. |
| [22] | 郝占庆, 于德永, 杨晓明, 丁之慧. 长白山北坡植物群落α多样性及其随海拔梯度的变化. 应用生态学报, 2002, 13(7): 786-789. |
| [23] | 郭忠玲, 马元丹, 郑金萍, 刘万德, 金哲峰. 长白山落叶阔叶混交林的物种多样性、种群空间分布格局及种间关联性研究. 应用生态学报, 2004, 15(11): 2013-2018. |