 2014, Vol. 34
2014, Vol. 34文章信息
- 郝建锋, 王德艺, 李艳, 姚小兰, 张逸博, 詹美春, 齐锦秋
- HAO Jianfeng, WANG Deyi, LI Yan, YAO Xiaolan, ZHANG Yibo, ZHAN Meichun, QI Jinqiu
- 人为干扰对川西金凤山楠木次生林群落结构和物种多样性的影响
- Effects of human disturbance on species diversity of Phoebe zhennan communitis in Jinfengshan Moutain in western Sichuan
- 生态学报, 2014, 34(23): 6930-6942
- Acta Ecologica Sinica, 2014, 34(23): 6930-6942
- http://dx.doi.org/10.5846/stxb201401140103
-
文章历史
- 收稿日期:2013-01-14
- 修订日期:2014-04-04






2. 水土保持与荒漠化防治重点实验室, 雅安 625014;
3. 木材工业与家具工程重点实验室, 雅安 625014
2. Soil and Water Conservation, Desertification Combating of MOE Sichuan Provincial Colleges and Universities Key Laboratory, Ya'an 625014, China;
3. Wood Industry and Furniture Engineering Sichuan Provincial Colleges and Universities Key Laboratory, Ya'an 625014, China
生态系统功能和服务的稳定与持续是人类生存的基础,生物多样性是影响生态系统功能和服务发挥的最关键因素之一[1, 2, 3],在全球变化的背景下,生物多样性与生态系统的稳定性成为国际生态学界关注的热点[1, 4]。人为干扰在全球气候和环境的重大变化中起着主导作用,是近百年来全球物种多样性的锐减的主要原因[5, 6]。国内外学者对放牧、采伐、旅游活动等人为干扰对物种多样性的影响做过大量的研究[4, 5, 7, 8, 9, 10],研究范围遍及热带森林、草原、湿地、农业及城市等景观类型[7, 8, 9, 10, 11, 12],但仍以草地为主[5, 7, 8, 9],对陆地生态系统中生产力最高、组成最复杂的森林生态系统的研究却为之不多[1],对人为干扰下森林群落的多样性与稳定性的关系很少有研究涉及[13]。关于人为干扰强度的很多研究认为,中度干扰下的物种多样性有增加的趋势[5, 9, 14],即“中度干扰假说”;关于干扰对植物群落层次的影响的很多研究认为,各层次对干扰的反应存在差异,但关于干扰对群落哪一层次影响最大仍无定论[3, 5, 14, 15]。中度干扰能否增加森林群落的物种多样性水平?森林群落各层次对干扰的反应如何?这些问题都未能得到很好的解决。
2013年结束的第八次全国森林资源清查显示,我国森林覆盖率提高到21.63%;但是普遍存在的以获取林产品为导向、经济利益为重的森林经理方式却不甚合理,森林经营者广植马尾松(Pinus massoniana)[16]、杉木(Cunninghamia lanceolata)[17]、桉(Eucalyptus robusta)[18]等速生丰产林。但是,人工纯林的整体功能远不及天然林和次生林,物种多样性剧烈下降、病虫害大爆发等问题使人们更多地关注人工林经营的可持续性。如何调控森林经理方式才能达到森林生产力的持续与物种多样性的保护和生态系统整体功能的维持之间的平衡?这是走林业可持续发展道路的亟待解决的问题。近年来,鉴于人工纯林暴露的诸多不足,国家开始倡导培育珍稀名贵树种,以期获取比速生树种更高经济效益,并且实现恢复和保护物种多样性的目的。楠木(Phoebe zhennan)[19],樟科楠木属,国家二级保护渐危种,俗称“金丝楠木”,天然分布极少,具有极高的经济和景观价值,所以受到众多森林经营者的青睐,但楠木群落环境和物种多样性现状却十分堪忧。现存的关于楠木研究多集中在楠木人工林群落生物量、凋落物等方面[19],因此,本研究以金凤山旅游区的楠木次生林群落为对象,探讨以选择性采伐和旅游活动为主的人为干扰对其物种组成、群落结构和物种多样性的影响,以期为该旅游区的楠木次生林群落物种多样性保护、可持续经营和发挥生态系统整体功能提供科学依据。
1 研究区域与研究方法 1.1 研究区概况研究区位于四川省雅安市雨城区的金凤山(30°04′47″—30°04′59″N,103°02′50″—103°02′57″E),地质构造为川西沉降褶皱带,地形呈西南东北走向,最高峰海拔1000 m;亚热带季风性湿润气候,年均温16.1 ℃,年极端平均最高气温16.9 ℃,年极端平均最低气温15.4 ℃,大于10 ℃的积温约为5000 ℃,全年日照时数约1000 h;位于华西雨屏区的雅安素有“天漏”之称[20],雨量充沛,年降水量约为1700 mm,某些年份高达2000 mm;土壤为紫色土,成土母质为第三系名山群钙质紫色砂、泥岩的残基物、坡基物和洪基物。本研究区属于亚热带季风性湿润气候常绿落叶混交林区,多为常绿樟栎林、针叶次生林等,森林覆被率达90%。
坐落于金凤山的金凤寺是川藏线上的著名寺庙,金凤寺的僧人对楠木的栽植始于唐初,由于年数久远,僧人抚育得当,楠木人工林至今已演替为物种丰富度高、林相丰富、林分郁闭度高的楠木次生林群落。然而,近年来金凤寺的旅游活动日益频繁,观赏树形良好、经济价值极高的楠木古树是游客主要的旅游动机之一,楠木次生林受到人为践踏、折枝采叶等不同程度的旅游干扰;此外,经营者因楠木巨大的景观和经济价值而视其为特殊保护对象,选择性剔除其他树种,楠木次生林群落的林分趋于单一、林相遭受破坏。森林采伐和旅游活动成为影响楠木次生林群落物种多样性和生态系统稳定性的主要人为干扰形式,并且与金凤寺核心景区距离越近,选择性伐除越明显,旅游活动越频繁,人为干扰强度越大。
1.2 研究方法 1.2.1 样地的选择与设置参照鲁庆彬[5]、吴甘霖[14]等对人为干扰强度的划分方法,根据与金凤寺核心景区距离的远近来划分人为干扰强度,距离越近,选择性采伐和旅游活动对楠木次生林群落的干扰强度越大;距离越远,则反之。为研究不同强度人为干扰对楠木次生林群落物种多样性的影响,实地考察发现,远离核心景区120m以外的林区很少受到选择性采伐和旅游活动为主的人为干扰,因此本研究将人为干扰强度划分为以下3级:重度干扰(紧靠核心景区0—40 m:选择性采伐明显,林分单一,楠木占据乔木层的大部分空间,少见其他树种;旅游活动频繁,游客进入量大,草本层植被遭受严重践踏,普见人为折枝痕迹等);中度干扰(距离核心景区40—80 m:选择性采伐较弱,有较多其他树种存在;游客进入量较少,草本层植被遭受轻度践踏,人为折枝痕迹较少等);轻度干扰(远离核心景区80—120 m:选择性采伐不明显,林相丰富,有较多的其他树种存在;几乎无游客进入,草本层植被很少遭到践踏,几乎无人为折枝的痕迹等)。在充分考察的基础上,设置9块20 m×20 m林相整齐、林层丰富、具有代表性的样地(表 1),其中轻度、中度和重度干扰样地各3块。将每个样地等分为4个10 m×10 m的乔木样方,并在每个样地中沿对角线选取6个5 m×5 m的灌木样方和12个1 m×1 m的草本样方进行调查,乔木层、灌木层、草本层样方总数分别为36个、54个、108个。
| 样地编号 Sample plot No. | 海拔 Altitude/m | 坡度 Slope/(°) | 坡向 Aspect/(°) | 平均胸径 Average diameter/cm | 平均高 Average height/m | 密度 Density/ (株/hm2) | 郁闭度 Canopy | 干扰强度 Disturbance intensity |
| 1 | 788 | 17 | NW50 | 11.3 | 9.4 | 1850 | 0.8 | 轻度 |
| 2 | 785 | 19 | NE45 | 11.5 | 9.9 | 1655 | 0.9 | 轻度 |
| 3 | 783 | 19 | NE45 | 10.1 | 8.9 | 1675 | 0.8 | 轻度 |
| 4 | 788 | 20 | ES45 | 10.9 | 8.4 | 1200 | 0.8 | 中度 |
| 5 | 787 | 20 | ES45 | 9.2 | 8.7 | 1300 | 0.8 | 中度 |
| 6 | 785 | 20 | ES45 | 10.4 | 8.9 | 1150 | 0.7 | 中度 |
| 7 | 787 | 19 | WN30 | 16.4 | 10. 8 | 1300 | 0.8 | 重度 |
| 8 | 788 | 23.5 | WN30 | 21.5 | 13.8 | 1225 | 0.7 | 重度 |
| 9 | 790 | 21 | WN50 | 23.4 | 15.3 | 1450 | 0.7 | 重度 |
测定和统计内容有
(1)乔木层:测定树高≥3 m的所有植株,记录其种类、胸径、高度和冠幅;(2)灌木层:测定所有树高<3 m的木本个体,包括乔木幼苗和幼树,记录其种类、株数(丛数)、高度和冠幅;(3)草本层:统计包括草质藤本和蕨类植物,但大型木质藤本按胸径大小分别计入乔木层或灌木层,记录其种类、株数(丛数)、高度和盖度。
1.2.2 楠木次生林群落结构依据孟祥楠[21]等对乔木树种胸径和高度的划分方法,测定楠木次生林群落中高度不小于3 m乔木个体的胸径和高度。将乔木个体的胸径分为9个径级,每4 cm为一个径级,依次为:胸径Ⅰ 0—4 cm,胸径Ⅱ 4—8 cm,胸径Ⅲ 8—12 cm,胸径Ⅳ 12—16 cm,胸径Ⅴ 16—20 cm,胸径Ⅵ 20—24 cm,胸径Ⅶ 24—28 cm,胸径Ⅷ 28—32 cm,胸径Ⅸ 32 cm及以上;统计不同强度人为干扰下楠木次生林群落的乔木层中各径级的个体数,并绘制径级结构图(图 1);将乔木个体的高度分为9个高度级,每2 m为一个高度级,依次为:高度Ⅰ 3—5 m,高度Ⅱ 5—7 m,高度Ⅲ 7—9 m,高度Ⅳ 9—11 m,高度Ⅴ 11—13 m,高度Ⅵ 13—15m,高度Ⅶ 15—17 m,高度Ⅷ 17—19 m,高度Ⅸ 19 m及以上;统计受不同强度人为干扰群落乔木层中各高度级的个体数,并绘制高度级结构图(图 1)。

|
| 图 1 不同强度人为干扰下楠木次生林群落乔木层的径级和高度级结构 Fig. 1 The diameter class and height class in tree layer of Phoeloe zhennan community under different level of disturbance 不同小写字母表示不同径级间、高度级间的数据差异显著(P<0.05) |
根据样地资料,计算不同物种的相对密度、相对显著度(相对盖度)和相对频度,计算每个样地不同物种的重要值(IV),并以物种的重要值为基础,计算物种多样性指数。本文采用物种丰富度指数S值、Shannon-Wiener多样性指数H值、Simpson优势度指数D值和Pielou均匀度指数Jsw值来综合评价楠木次生林群落的物种多样性:
相对密度=某个种的株数/所有种的总株数
相对显著度=某个种的胸高断面积/所有种的胸高断面积之和
相对频度=某个种在样方中出现的次数/所有种出现的总次数
相对盖度=某个种的盖度/所有种盖度之和
重要值:乔木层
IV=(相对密度+相对显著度+相对频度)/3
灌木层、草本层
IV=(相对密度+相对盖度+相对频度)/3
丰富度指数S=出现在样方的物种数
Simpson优势度指数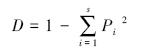
Shannon-Wiener多样性指数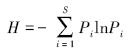
Pielou均匀度指数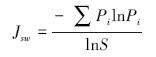
式中,Pi为第i种的个体数ni占所有种个体总数n的比例,ni为第i种的个体数,n为所有种的个体总数,即Pi=ni/n;i=1,2,3,…,S,S为物种数。
1.2.4 数据处理文中所有数据采用SPSS17.0统计软件、Origin8.0软件和Excel进行统计分析,采用单因素方差分析(one-Way ANOVA)和最小显著差异法(LSD)检验群落各层次多样性指数、高度级和径级数据的显著性差异(P=0.05)。
2 结果与分析 2.1 不同强度人为干扰对楠木次生林群落结构的影响 2.1.1 群落径级结构植物群落的径级结构是最基本的群落结构,是植物生长与环境关系的综合反映,成为评价植物群 落受干扰程度、稳定性、生长发育状况、预测群落结构发展的重要指标[22]。从图 1可以看出,不同强度人为干扰下楠木次生林群落的径级结构差异显著,乔木个体在各高度级的数量分布不同;轻度和中度干扰下乔木层的径级都在径级Ⅱ(径级4—8 cm)处出现数量的峰值,分别为64株和67株,并向两侧呈递减趋势,林分存在大量幼树个体,说明林分具有潜在的天然更新能力;受重度干扰的乔木个体的多分布在径级Ⅰ(胸径0—4 cm)、径级Ⅱ(胸径4—8 cm)和径级Ⅸ(胸径32 cm及以上),大径级和小径级个体多,这是因为楠木受到特殊保护,人为选择性采伐其他树种,楠木易形成大径级个体,但过分对楠木的保护抑制了其他树种的长势,其他树种多为小径级个体。
2.1.2 群落高度级结构植物群落高度级结构不仅能反映群落垂直结构的物种组成性状,还能体现出垂直空间层次上的物种多样性和动态性[3],定量分析和评价植物群落高度级结构有助于掌握群落结构的复杂性、发展阶段和稳定程度。从图 1可以看出,不同人为干扰强度对楠木次生林群落乔木层的高度级结构的影响差异显著,乔木个体在各高度级的数量分布不同;轻度和中度干扰下乔木层的高度级的峰值都出现在高度级Ⅲ(高度7—9 m),个体数分别为55株和60株,并向两侧递减,低高度级(高度3—9 m)和中高度级(高度9—15 cm)的乔木个体多,高高度级(高度15 m及以上)个体分布稀少,这是因为群落受人为干扰的影响较轻,群落结构在垂直方向上趋于稳定;受重度干扰群落的乔木个体呈分散分布,在低高度级和高高度级较多,中高度级个体数少,说明受重度干扰的群落垂直分布结构不稳定,群落垂直空间层次上的物种多样性低。
2.2 不同强度人为干扰对楠木次生林群落物种组成的影响 2.2.1 科属种组成分析在总面积为3600 m2的9个样地中,共记录到维管束植物155种,隶属于69科136属,以禾本科(Gramineae)、樟科(Lauraceae)、菊科(Compositae)、荨麻科(Urticaceae)、蔷薇科(Rosaceae)、壳斗科(Fagaceae)、百合科(Liliaceae)等为主。不同群落层次的物种组成不同(图 2),乔木层有31种,隶属于19科28属,以樟科、壳斗科、大戟科(Euphorbiaceae)为主;灌木层有51种,隶属于27科42属,以樟科、壳斗科、蔷薇科、桑科(Moraceae)为主;草本层有87种,隶属于42科79属,以禾本科、菊科、荨麻科、百合科居多;可见草本层对物种多样性的贡献最大,灌木层次之,乔木层物种组成比较单一。不同强度人为干扰下楠木次生林群落的物种组成不同(图 2),轻度干扰样地有113种,隶属于86属57科,中度干扰样地有88种,隶属于63属51科,重度干扰样地有77种,隶属于62属51科,物种数、属数和科数随着干扰强度的增加而降低。

|
| 图 2 不同人为干扰强度和群落层次科、属、种的数量 Fig. 2 The statistics of the number of Family,Genus,Species of Phoeloe zhennan community in different layer and under different level of disturbance |
重要值是反映某个物种在森林群落中作用和地位的综合数量指标,通过分析重要值可以找出群落中的主要优势种,调查结果表明:不同强度人为干扰下楠木次生林群落各层次物种组成和优势种不同。
表 2表明,不同强度人为干扰下乔木层的优势种的数量排序为:轻度干扰>中度干扰>重度干扰,轻度干扰下乔木层的优势种有青冈(Cyclobalanopsis glauca)、楠木、银杏(Ginkgo biloba)、润楠(Machilus pingii)等10种,楠木的重要值为0.1115,仅次于青冈(0.1173);中度干扰下乔木层的优势种有青冈、润楠、楠木、柳杉(Cryptomeria fortunei)等9种,楠木的重要值为0.1335,次于青冈(0.1432)和润楠(0.1341);重度干扰下乔木层的有楠木、枫香树(Liquidambar formosana)等6种,楠木的重要值为0.5095,是绝对优势种,居于其次的枫香树,其重要值急剧下降为0.0816,说明重度干扰使物种组成发生急剧变化,这是因为人为选择性剔除其他树种,对楠木进行特殊保护使楠木占据了林地大部分空间,致使其他树种与楠木竞争林地资源空间的能力下降。
| 物种 Species name | 重要值 Important value | ||
| A | B | C | |
| A:轻度干扰Slight disturbance;B:中度干扰Medium disturbance;C:重度干扰Severe disturbance | |||
| 青冈Cyclobalanopsis glauca | 0.1173 | 0.1432 | |
| 楠木Phoebe zhennan | 0.1115 | 0.1335 | 0.5095 |
| 银杏Ginkgo biloba | 0.1109 | ||
| 润楠Machilus pingii | 0.1078 | 0.1341 | 0.0399 |
| 杉木Cunninghamia lanceolata | 0.0967 | 0.1114 | |
| 野桐Mallotus japonicas var. floccosus | 0.0773 | 0.0847 | |
| 柳杉Cryptomeria fortunei | 0.0773 | 0.0931 | |
| 枹栎Quercus serrate | 0.0462 | 0.0515 | 0.0735 |
| 罗汉松Podocarpus macrophyllus | 0.0461 | ||
| 灯台树Bothrocaryum controversum | 0.0407 | 0.0472 | |
| 毛脉南酸枣Choerospondias axillaris var. pubinervis | 0.0400 | ||
| 枫香树Liquidambar formosana | 0.0816 | ||
| 亮叶猴耳环Pithecellobium lucidum | 0.0796 | ||
| 栲Castanopsis fargesii | 0.0488 | ||
| A:其余11种 The 11 remaining species belonging to A | 0.1681 | ||
| B:其余10种 The 10 remaining species belonging to B | 0.1614 | ||
| C:其余9种 The 9 remaining species belonging to C | 0.1672 | ||
| 合计 Total | 1.0000 | 1.0000 | 1.0000 |
表 3表明,不同强度人为干扰下灌木层的优势种的数量排序为:中度干扰>轻度干扰>重度干扰,中度干扰下灌木层的优势种最多,为水麻(Debregeasia orientalis)、紫麻(Oreocnide frutescens)、五加(Acanthopanax gracilistylus)等15种;轻度干扰下灌木层的优势种数量居于其次,有细叶楠(Phoebe hui)、火棘(Pyracantha fortuneana)、柳杉等13种;受重度干扰群落的灌木层优势树种为铁仔(Myrsine africana)、火棘等11种。在重度干扰的乔木层中,占据绝对优势地位的楠木可能抑制林下灌木的生长,不同植物对环境的适应能力有差异,只有火棘、铁仔、棕竹(Rhapis excelsa)、慈竹(Neosinocalamus affinis)等耐阴性、抗逆性强树种的生长,耐受性差的物种被淘汰。此外,在楠木次生林群落中未见楠木幼树幼苗,说明楠木几乎无天然更新能力,但在乔木层和灌木层中常见细叶楠、润楠、黑壳楠(Lindera megaphylla)、水青冈(Fagus longipetiolata)等乡土树种,这些树种具有潜在的天然更新能力。
| 物种 Species name | 重要值 Important value | ||
| A | B | C | |
| A:轻度干扰Slight disturbance;B:中度干扰Medium disturbance;C:重度干扰Severe disturbance | |||
| 细叶楠Phoebe hui | 0.1526 | 0.0463 | 0.0407 |
| 火棘Pyracantha fortuneana | 0.1152 | 0.1155 | |
| 柳杉Cryptomeria fortunei | 0.1029 | ||
| 水麻Debregeasia orientalis | 0.0816 | 0.1290 | |
| 黄牛奶树Symplocos laurina | 0.0696 | ||
| 冬青Ilex chinensis | 0.0522 | ||
| 铁仔Myrsine Africana | 0.0437 | 0.0622 | 0.2848 |
| 山茶Camellia japonica | 0.0390 | ||
| 水青冈Fagus longipetiolata | 0.0363 | 0.0409 | |
| 棕竹Rhapis excels | 0.0363 | 0.0395 | |
| 异叶榕Ficus heteromorpha | 0.0288 | 0.0472 | |
| 杉木Cunninghamia lanceolata | 0.0276 | ||
| 悬钩子属Rubus | 0.0258 | 0.0632 | |
| 紫麻Oreocnide frutescens | 0.1183 | ||
| 五加Acanthopanax gracilistylus | 0.0688 | ||
| 润楠Machilus pingii | 0.0476 | 0.0466 | |
| 枸杞Lycium chinense | 0.0442 | ||
| 庐山梣Fraxinus mariesii | 0.0354 | ||
| 假豪猪刺Berberis soulieana | 0.0322 | ||
| 黑壳楠Lindera megaphylla | 0.0279 | 0.0363 | |
| 小果蔷薇Rosa cymosa | 0.0259 | ||
| 花椒Zanthoxylum bungeanum | 0.0220 | ||
| 栲Castanopsis fargesii | 0.1142 | ||
| 慈竹Neosinocalamus affinis | 0.0469 | ||
| 柃木Eurya japonica | 0.0373 | ||
| 白花泡桐Paulownia fortunei | 0.0297 | ||
| 毛豹皮樟Litsea coreana var. lanuginosa | 0.0295 | ||
| A:其余18种 The 18 remaining species belonging to A | 0.1883 | ||
| B:其余17种 The 17 remaining species belonging to B | 0.1888 | ||
| C:其余16种 The 16 remaining species belonging to C | 0.1791 | ||
| 合计 Total | 1.0000 | 1.0000 | 1.0000 |
表 4表明,不同强度人为干扰下草木层的优势种的数量排序为:轻度干扰>中度干扰>重度干扰,受轻度干扰的草本层的优势种有渐尖毛蕨(Cyclosorus acuminatus)、里白(Hicriopteris glauca)、乌蕨(Stenoloma chusanum)、水竹叶(Murdannia triquetra)、 冷水花(Pilea notate)等34种,而受中度和重度干扰的草本层优势种仅为18种和16种。轻度干扰下草本层物种丰富度最高可能有以下两方面的原因:一是草本层植被主要受到人为践踏等旅游活动的干扰,受轻度干扰的草本层距离核心景区远,少有游客进入,几乎无人为践踏等干扰;二是轻度干扰下整个群落环境较少受到人为干扰的影响弱,群落结构和功能比较健全,草本植物生长条件良好。
| 物种 Species name | 重要值 Important value | ||
| A | B | C | |
| A:轻度干扰Slight disturbance;B:中度干扰Medium disturbance;C:重度干扰Severe disturbance | |||
| 渐尖毛蕨Cyclosorus acuminatus | 0.0778 | 0.0379 | |
| 里白Hicriopteris glauca | 0.0768 | ||
| 乌蕨Stenoloma chusanum | 0.0548 | 0.0745 | 0.1058 |
| 水竹叶Murdannia triquetra | 0.0385 | ||
| 冷水花Pilea notate | 0.0365 | 0.0782 | |
| 长叶苎麻Boehmeria penduliflora | 0.0354 | ||
| 皱叶狗尾草Setaria plicata | 0.0327 | ||
| 长柱头薹草Carex teinogyna | 0.0307 | 0.0456 | |
| 荩草Arthraxon hispidus | 0.0291 | 0.0752 | 0.0397 |
| 薄叶卷柏Selaginella delicatula | 0.0288 | 0.0312 | 0.0740 |
| 石海椒Reinwardtia indica | 0.0273 | ||
| 华中瘤足蕨Plagiogyria euphlebia | 0.0249 | 0.0537 | 0.0760 |
| 问荆Equisetum arvense | 0.0244 | ||
| 南方复叶耳蕨Arachniodes australis | 0.0221 | 0.1544 | |
| 鸢尾Iris tectorum | 0.0206 | 0.0185 | |
| 紫苏Perilla frutescens | 0.0198 | ||
| 野茼蒿Crassocephalum crepidioides | 0.0179 | ||
| 马唐Digitaria sanguinalis | 0.0176 | ||
| 吉祥草Reineckia carnea | 0.0171 | 0.0235 | 0.0307 |
| 蛇莓Duchesnea indica | 0.0166 | ||
| 海金沙Lygodium japonicum | 0.0159 | 0.0258 | |
| 画眉草Eragrostis pilosa | 0.0134 | ||
| 白酒草Conyza japonica | 0.0132 | ||
| 鹅观草Roegneria kamoji | 0.0127 | ||
| 三脉紫菀Aster ageratoides | 0.0126 | ||
| 小花叶底红Phyllagathis fordii var. micrantha | 0.0114 | ||
| 蒲公英Taraxacum mongolicum | 0.0112 | 0.0185 | |
| 平车前Plantago depressa | 0.0103 | ||
| 通泉草Mazus pumilus | 0.0099 | ||
| 积雪草Centella asiatica | 0.0098 | ||
| 一年蓬Erigeron annuus | 0.0098 | ||
| 酢浆草Oxalis corniculata | 0.0096 | 0.0263 | 0.0167 |
| 土牛膝Achyranthes aspera | 0.0096 | ||
| 马兰Kalimeris indica | 0.0090 | ||
| 姜花Hedychium coronarium | 0.0376 | 0.1075 | |
| 盾蕨Neolepisorus ovatus | 0.0294 | ||
| 薯蓣Dioscorea opposite | 0.0271 | 0.0186 | |
| 麦冬Ophiopogon japonicus | 0.0235 | 0.0346 | |
| 蕺菜Houttuynia cordata | 0.0207 | ||
| 变豆菜Sanicula chinensis | 0.0194 | ||
| 鸭跖草Commelina communis | 0.1527 | ||
| 八角莲Dysosma versipellis | 0.0322 | ||
| 光头稗Echinochloa colonum | 0.0318 | ||
| 白车轴草Trifolium repens | 0.0282 | ||
| 虎耳草Saxifraga stolonifera | 0.0226 | ||
| 铜锤玉带草Pratia nummularia | 0.0199 | ||
| A:其余36种 The 36 remaining species belonging to A | 0.1920 | ||
| B:其余27种 The 27 remaining species belonging to B | 0.1976 | ||
| C:其余26种 The 26 remaining species belonging to C | 0.1904 | ||
| 合计 Total | 1.0000 | 1.0000 | 1.0000 |
总体上看,楠木次生林群落的物种丰富度指数S值、Shannon-Wiener多样性指数H值、Simpson优势度指数D值和均匀度指数Jsw值随着人为干扰强度的增加而降低(表 5),物种多样性水平逐渐下降。重度干扰样地紧邻金凤寺核心景区,因大量种植楠木而选择性采伐其他树种,旅游活动频繁,人为干扰对生境的破坏度高,一些耐受性差的物种因不适应生境的剧烈变化而被淘汰,所以S值最低;轻度干扰样地与金凤寺核心景区距离最远,较少受到人为干扰的影响,所以S值最大;中度干扰样地的S值居中。本研究中的H值与S值的变化趋势一致,重度干扰下群落的H值最低,中度干扰次之,轻度干扰下群落的H值最高,说明楠木次生林群落的物种多样性水平随着干扰强度的增加而降低。均匀度指数Jsw反映群落中物种分布的均匀程度,由H值和S值共同决定,一般H值和S值越大,Jsw值越大[3];轻度和中度干扰下群落的Jsw值较大,这因为其H值和S值都较大,而重度干扰下群落的H值和S值最小,因此其Jsw值最小,说明重度干扰使物种在生境中分布趋于不均匀。
由上述结果可知,随着人为干扰强度增加,群落物种多样性整体呈下降趋势:轻度干扰>中度干扰>重度干扰,人为干扰对楠木次生林群落的物种多样性产生负面影响明显。干扰强度与金凤寺核心景区的距离有关,表现为:距离越近,旅游活动越频繁,选择性地伐除除楠木以外的其他树种的行为越明显,楠木的生态优势明显,物种在群落生境中分布不均匀,一些耐受性差的物种消失,物种丰富度指数、Shannon-Wiener多样性指数和均匀度指数均下降;距离越远,群落林分较少受到选择性采伐和旅游活动的影响,更多的物种能在群落环境中生存,物种多样性指数逐渐增加。
2.3.2 植物群落层次物种多样性植物群落层次物种多样性反映了植物的立体结构和群落中乔灌草的组合状况,良好的群落层次结构有利于形成稳定的群落环境。通过对乔木层、灌木层和草本层多样性指数的计算(表 5),轻度、中度和重度人为干扰下群落层次多样性差异显著,物种丰富度指数S值、Shannon-Wiener多样性指数H值、Simpson优势度指数D值一般为草本层>灌木层>乔木层,这是因为草本植物、灌木植物、乔木植物对干扰环境的适应能力依次降低所致[3]。受轻度、中度和重度干扰的乔木层和草本层的各项指数均为轻度干扰>中度干扰>重度干扰;灌木层的各项指数均为中度干扰>轻度干扰>重度干扰,中度干扰下灌木层的物种多样性有增加的趋势。Andreas Mlder[23]等认为乔木层的物种组成会通过营养循环、透光率、土壤水分供应等生态过程对林下植被产生影响,杨梅[15]等研究发现林下层的物种更新也会受到乔木层的影响。在本研究中,受重度干扰的乔木层楠木的重要值高达0.5095,优势度极高,乔木层物种组成单一,进而影响林下灌木和草本的生长,所以受重度干扰的楠木次生林群落各层次物种多样性都处于最低水平;受中度干扰的乔木层物种组成较为丰富,群落环境条件较好,灌木层和草本层的物种多样性维持在较高水平;受轻度干扰群落远离金凤寺核心景区,生境较少遭受人为干扰,乔木层物种组成丰富,所以轻度干扰下各层次物种多样性指数都维持在较高的水平。
| 干扰强度 Disturbance intensity | 植物层 Plant layer | S | D | H | Jsw |
| 不同小写字母表示不同群落层次间的数据差异显著(P<0.05) The difference normal letters in different plant layer indicate significant differences at P<0.05;S:物种丰富度指数The richness index;D:Simpson优势度指数Simpson index;H:Shannon-Wiener多样性指数Shannon-Wiener index;Jsw:Pielou均匀度指数Pielou index | |||||
| 轻度干扰 | 乔木 | 13.3333±0.3333b | 0.8540±0.0012a | 2.2187±0.0297b | 0.8567±0.0038a |
| Slight disturbance | 灌木 | 18.6667±2.6667ab | 0.8207±0.0580a | 2.2657±0.2090ab | 0.7777±0.0563a |
| 草本 | 28.3333±3.8442a | 0.9167±0.0085a | 2.7997±0.0575a | 0.8083±0.0212a | |
| 中度干扰 | 乔木 | 12.6667±0.6667b | 0.7667±0.0657a | 1.9247±0.1601b | 0.7620±0.0771a |
| Medium disturbance | 灌木 | 20.6667±1.3333a | 0.9053±0.0081a | 2.6707±0.0738a | 0.8830±0.0136a |
| 草本 | 23.3333±0.8819a | 0.8970±0.0120a | 2.5427±0.0998a | 0.8073±0.0287a | |
| 重度干扰 | 乔木 | 8.6667±1.7638b | 0.6850±0.0801a | 1.5617±0.2603a | 0.7270±0.0612a |
| Medium disturbance | 灌木 | 14.6667±1.2019ab | 0.7260±0.0526a | 1.8350±0.2002a | 0.6817±0.0537a |
| 草本 | 20.3333±1.2019a | 0.8573±0.0069a | 2.2663±0.0676a | 0.7530±0.0101a | |
植物群落结构是个体对外界干扰强度、立地条件优劣、环境适应性和多样性的反映,被认为是影响群落物种组成和多样性的最重要因素之一[3, 24]。金凤山楠木次生林群落的乔木层在轻度和中度人为干扰下,径级和高度级结构均呈近正态分布,幼树较多,说明其群落结构稳定且自然更新能力强;受重度干扰的乔木层的径级和高度级分布参差不齐,中径级和中高度级的个体少,两极分布明显,群落垂直结构不稳定。通过分析重要值发现,受重度干扰的乔木层中楠木的重要值高达0.5095,楠木是绝对优势种,林分中未见楠木幼树幼苗,说明重度干扰导致楠木丧失自然更新能力。Anitha K[11]等研究发现持续的重度干扰会改变群落结构、植被类型、群落演替方向等,因此,笔者认为若选择性采伐和高强度旅游活动等人为干扰持续作用于楠木次生林群落,将会对群落结构造成不可逆的破坏,致使楠木次生林群落退化为人工林。
金凤山楠木次生林群落的物种丰富度指数S值和Shannon-Wiener指数H值随着人为干扰强度的增加而降低,重度干扰对群落物种多样性的负面影响最大,中度干扰次之,轻度干扰最小,这与许多研究认为中度干扰增加物种多样性的结论不一致[5, 9, 14]。一些研究认为人为干扰对灌木层的负面影响最大[5],本研究结果却与之相反,受中度干扰的灌木层的S值和H值均高于受轻度和重度干扰的灌本层,物种多样性水平呈增加的趋势;鲁庆彬[5]、杨梅[15]等研究发现不同人为干扰强度下群落各层次所呈现的物种多样性特征不同,对干扰的反应不一致,可能与各层次物种的耐受性、种群的繁殖策略等生态习性有关。综上研究,认为楠木次生林群落各层次的多样性特征不同可能与人为干扰的作用范围和不同层次植物的生态习性有关。选择性采伐和旅游活动这两种主要的干扰形式起作用的群落层次不同,选择性采伐主要影响乔木层,旅游活动对草本层的影响最严重,而居于二者中间的灌木层受到这两种干扰的影响相对较弱,此外,乔木和草本的长势因干扰而被削弱,为灌木的更好地利用生境资源提供了契机,所以灌木层的多样性水平较高。
生物多样性的生态系统功能是生物多样性科学领域的热点问题之一,人为干扰导致物种丧失,对植物群落的整体功能和服务产生重要影响[1]。经营者为追求楠木巨大的景观和经济价值,不惜选择性剔除其他树种,并开展不加控制的旅游活动,导致金凤山楠木次生林群落物种多样性水平显著降低(表 5)。Miyoshi在1925年曾预测纯粹地对日本红枫(Acer pycnanthum)进行特殊保护、选择性采伐其他树种会引起群落环境的衰退,后来Ikuyo Saeki的研究也证实了人为干扰对红枫群落造成的恶果[10];大量研究表明旅游活动带来的踩踏、折枝、游道等人为干扰对森林群落环境造成严重破坏[4, 5]。如何化解人为干扰与保护物种多样性的冲突?是否要将森林经营目标转移到生态保护上,摒弃森林的经济效益?Evan Davies[25]等研究发现我国海南省传统的营林理念完全以生态保护为重心,扼制了当地获取林产品的经济需要和黎族农户的“靠山吃山”生活需求,农户造林和护林的积极性受挫,盗取木材等现象频发。因此,笔者认为森林经营既要满足人类对森林的物质需求又要能实现森林的永续利用,实现森林多功能经营。在欧美发达国家实践成功的“近自然林业”理念[23, 26, 27]为实现森林多功能经营提供了一条可行途径,近自然林经营并不排斥获取森林的生产力,与传统森林经营理论相比,近自然经营理论认为只有实现最合理的近自然状态的森林才能实现经济利益最大化,在发挥森林的经济功能的同时,不偏废其生态功能和社会功能。因此,金凤山楠木次生林群落的恢复和营造可借鉴“异龄-混交-复层-多树种”的近自然经营方式,降低旅游活动和选择性采伐干扰的强度,栽植具有潜在更新能力的乡土树种,如细叶楠、润楠、青冈等;对林下幼苗进行抚育更新,实行混交造林;重视群落垂直方向上的多样性,适度间伐,调节林分结构,构造异龄林和复层林,最终营造物种多样性水平高、群落结构稳定、整体功能发挥良好的群落环境,使楠木次生林群落发挥最优的经济和生态功能。
| [1] | Ma K P. Studies on biodiversity and ecosystem function via manipulation experiments. Biodiversity Science, 2013, 21(3): 247-248. |
| [2] | Lu X L, Liang G F, Tang Q, Ding S Y, Li Q X, Zhang X Q. Plant species of the non-agricultural habitats in the lower reaches of the Yellow River plain agro-landscape. Acta Ecologica Sinica, 2014, 34(4): 789-797. |
| [3] | Yu W, Wan D, Wang S L, Yang D S. Community structure and species diversity of debris flow deposition area in southeast of Tibet, China. Journal of Mountain Research, 2013, 31(1): 120-126. |
| [4] | Suriguga, Zhang J T, Wang Y X. Species diversity of forest communities and its forecasting by neural network in the Songshan National Nature Reserve, Beijing. Acta Ecologica Sinica, 2013, 33(11): 3394-3403. |
| [5] | Lu Q B, You W Y, Zhao C J, Wang X W, Meng X X. Effects of tourism disturbance on plant diversity in Qingshan Lake scenic area of Zhejiang Province. The Journal of Applied Ecology, 2011, 22(2): 295-302. |
| [6] | Santangeli A, Wistbacka R, Hanski I K, Laaksonen T. Ineffective enforced legislation for nature conservation: A case study with Siberian flying squirrel and forestry in a boreal landscape. Biological Conservation, 2013, 157: 237-244. |
| [7] | Chen F R, Cheng J M, Liu W, Zhu R B, Yang X M, Zhao X Y, Su J S. Effects of different disturbances on diversity and biomass of communities in the typical steppe of loess region. Acta Ecologica Sinica, 2013, 33(9): 2856-2866. |
| [8] | Wang H M, Li Z H, Wang Z. Effects of climate and grazing on the vegetation cover change in Xilinguole League of Inner Mongolia, North China. Chinese Journal of Applied Ecology, 2013, 24(1): 156-160. |
| [9] | Zhou L, Shangguan T L, Guo D G, Gao H W, Wang Z. Response of species diversity in Caragana korshinskii communities to climate factors and grazing disturbance in Shanxi, Shaanxi, Ningxia and Inner Mongolia. Acta Ecologica Sinica, 2012, 32(1): 111-122. |
| [10] | Saeki I. Effects of tree cutting and mowing on plant species composition and diversity of the wetland ecosystems dominated by the endangered maple, Acer pycnanthum. Forest Ecology and Management, 2007, 242(2/3): 733-746. |
| [11] | Anitha K, Joseph S, Chandran R J, Ramasamy E V, Prasad S N. Tree species diversity and community composition in a human-dominated tropical forest of Western Ghats biodiversity hotspot, India. Ecological Complexity, 2010, 7(2): 217-224. |
| [12] | Muthulingam U, Thangavel S. Density, diversity and richness of woody plants in urban green spaces: A case study in Chennai metropolitan city. Urban Forestry and Urban Greening, 2012, 11(4): 450-459. |
| [13] | Ma K P. A mini review on the advancement of biodiversity research in China in 2012. Biodiversity Science, 2013, 21(1): 1-2. |
| [14] | Wu G L, Huang M Y, Duan R Y, Zhao K. Disturbing effects of tourism on species diversity in Pinus taiwanensis communities. Acta Ecologica Sinica, 2006, 26(12): 3924-3930. |
| [15] | Yang M, Lin S Z, Cao G Q. Species diversity of Castanopsis eyrei communities under different human-caused disturbance. Journal of Northeast Forestry University, 2009, 37(7): 30-32. |
| [16] | Li R X, Min J G, Peng T T, Liu N, Hao J P, Wang D, Guan Q W. Effects of thinning on understory species diversities of Pinus massoniana forests. Journal of Northwest A&F University: Natural Science, 2013, 41(3): 61-68. |
| [17] | Xu J L, Mao Y M, Zheng C Z, Fan Y D, Zhou S S, Chen Y H, Cheng X Y, Yu M K. Effect of thinning on growth and timber outturn in Cunninghamia lanceolata plantation. Forest Research, 2014, 27(1): 99-107. |
| [18] | Wang Z C, Du A M, Chen S X. Current situation and sustainable management strategy of Eucalypt plantations in China. Eucalypt Science and Technology, 2012, 29(4): 58-62. |
| [19] | Ma M D, Jiang H, Liu Y J. Biomass, carbon content, carbon storage and their vertical distribution of Phoebe bourmei artificial stand. Scientia Silvae Sinicae, 2008, 44(3): 34-39. |
| [20] | Zhang Z C, Chen C H, Yu Z G, Qi G N. Research on the planning of tea garden in Yaan. Journal of Sichuan Agricultural University, 2004, 22(3): 228-232. |
| [21] | Meng X N, Zhao Y S, Zheng L, Xin Y. Population structure and understory species diversity of different aged Pinus sylvestris var. mongolica plantations in Nenjiang Sandy Land of Northeast China. The Journal of Applied Ecology, 2012, 23(9): 2332-2338. |
| [22] | Xie X K, Su D K, Liu Z G, Yu D P, Zhou L, Dai L M. Modeling diameter distribution of primary broad leaf-Korean pine mixed forest in Changbai Mountains. Chinese Journal of Ecology, 2010, 29(8): 1477-1481. |
| [23] | Mölder A, Streit M, Schmidt W. When beech strikes back: How strict nature conservation reduces herb-layer diversity and productivity in Central European deciduous forests. Forest Ecology and Management, 2014, 319: 51-61. |
| [24] | Wehenkel C, Corral-Rivas J J, Gadow K V. Quantifying differences between ecosystems with particular reference to selection forests in Durango, Mexico. Forest Ecology and Management, 2014, 316: 117-124. |
| [25] | Davies E G R, Wismer S K. Sustainable forestry and local people: the case of Hainan's Li Minority. Human Ecology, 2007, 35(4): 415-426. |
| [26] | Zhang C Y, Zhao X H, Gadow K V. Analyzing selective harvest events in three large forest observational studies in North Eastern China. Forest Ecology and Management, 2014, 316: 100-109. |
| [27] | Schou E, Jacobsen J B, Kristensen K L. An economic evaluation of strategies for transforming even-aged into near-natural forestry in a conifer-dominated forest in Denmark. Forest Policy and Economics, 2012, 20: 89-98. |
| [1] | 马克平. 生物多样性与生态系统功能的实验研究. 生物多样性, 2013, 21(3): 247-248. |
| [2] | 卢训令, 梁国付, 汤茜, 丁圣彦, 李乾玺, 张晓青. 黄河下游平原农业景观中非农生境植物多样性. 生态学报, 2014, 34(4): 789-797. |
| [3] | 喻武, 万丹, 汪书丽, 杨东升. 藏东南泥石流沉积区植物群落结构和物种多样性特征. 山地学报, 2013, 31(1): 120-126. |
| [4] | 苏日古嘎, 张金屯, 王永霞. 北京松山自然保护区森林群落物种多样性及其神经网络预测. 生态学报, 2013, 33(11): 3394-3403. |
| [5] | 鲁庆彬, 游卫云, 赵昌杰, 王向伟, 孟秀祥. 旅游干扰对青山湖风景区植物多样性的影响. 应用生态学报, 2011, 22(2): 295-302. |
| [7] | 陈芙蓉, 程积民, 刘伟, 朱仁斌, 杨晓梅, 赵新宇, 苏纪帅. 不同干扰对黄土区典型草原物种多样性和生物量的影响. 生态学报, 2013, 33(9): 2856-2866. |
| [8] | 王海梅, 李政海, 王珍. 气候和放牧对锡林郭勒地区植被覆盖变化的影响. 应用生态学报, 2013, 24(1): 156-160. |
| [9] | 周伶, 上官铁梁, 郭东罡, 高洪文, 王赞. 晋、陕、宁、蒙柠条锦鸡儿群落物种多样性对放牧干扰和气象因子的响应. 生态学报, 2012, 32(1): 111-122. |
| [13] | 马克平. 2012年中国生物多样性研究进展简要回顾. 生物多样性, 2013, 21(1): 1-2. |
| [14] | 吴甘霖, 黄敏毅, 段仁燕, 赵凯. 不同强度旅游干扰对黄山松群落物种多样性的影响. 生态学报, 2006, 26(12): 3924-3930. |
| [15] | 杨梅, 林思祖, 曹光球. 不同人为干扰强度下甜槠林群落物种多样性比较分析. 东北林业大学学报, 2009, 37(7): 30-32. |
| [16] | 李瑞霞, 闵建刚, 彭婷婷, 刘娜, 郝俊鹏, 王东, 关庆伟. 间伐对马尾松人工林植被物种多样性的影响. 西北农林科技大学学报: 自然科学版, 2013, 41(3): 61-68. |
| [17] | 徐金良, 毛玉明, 郑成忠,范荣德, 周世水, 陈永辉, 成向荣, 虞木奎. 抚育间伐对杉木人工林生长及出材量的影响. 林业科学研究, 2014, 27(1): 99-107. |
| [18] | 王志超, 杜阿朋, 陈少雄. 我国桉树人工林现状及可持续经营对策研究. 桉树科技, 2012, 29(4): 58-62. |
| [19] | 马明东, 江洪, 刘跃建. 楠木人工林生态系统生物量、碳含量、碳贮量及其分布. 林业科学, 2008, 44(3): 34-39. |
| [20] | 张泽岑, 陈昌辉, 余宗国, 齐桂年. 雅安市茶叶区划设计研究. 四川农业大学学报, 2004, 22(3): 228-232. |
| [21] | 孟祥楠, 赵雨森, 郑磊, 辛颖. 嫩江沙地不同年龄樟子松人工林种群结构与林下物种多样性动态. 应用生态学报, 2012, 23(9): 2332-2338. |
| [22] | 谢小魁, 苏东凯, 刘正纲, 于大炮, 周莉, 代力民. 长白山原始阔叶红松林径级结构模拟. 生态学杂志, 2010, 29(8): 1477-1481. |