 2014, Vol. 34
2014, Vol. 34文章信息
- 景慧娟, 凡强, 王蕾, 廖文波, 陈春泉, 彭少麟
- JING Huijuan, FAN Qiang, WANG Lei, LIAO Wenbo, CHEN Chunquan, PENG Shaolin
- 江西井冈山地区沟谷季雨林及其超地带性特征
- The ravine monsoon rain forest in Jinggangshan of Jiangxi Province and its super zonal characteristics
- 生态学报, 2014, 34(21): 6265-6276
- Acta Ecologica Sinica, 2014, 34(21): 6265-6276
- http://dx.doi.org/10.5846/stxb201405291106
-
文章历史
- 收稿日期:2014-5-29
- 修订日期:2014-9-28






2. 首都师范大学资源环境与旅游学院, 北京 100048;
3. 江西井冈山国家级自然保护区管理局, 井冈山 343600
2. College of Resource Environment and Tourism, Capital Normal University, Beijing 100048, China;
3. Administration Bureau of Jinggangshan National Nature Reserve, Jinggangshan 343600, China
井冈山地区位于北南走向的罗霄山脉的中段,向南延伸与南岭垂直。井冈山地貌复杂,沟壑纵横,相对海拔差高达1920m(全区海拔最低为200m,主峰南风面高达2120.4m)。井冈山地处中国大陆东部湿润季风气候区,中亚热带的南缘,水、热条件充沛,是亚洲东部生物多样性高丰度区。地带性植被以中亚热带常绿阔叶林为主[1]。在低海拔的沟谷地段,分布有具南亚热带季风常绿林与中亚热带常绿阔叶林过渡特征的中亚热带沟谷雨林,在植被演替和植物区系区划上具有重要意义[2]。
亚热带沟谷雨林是随着温度条件的下降,由热带雨林向亚热带常绿阔叶林过渡的居间类型,是受制于温度因子的纬度地带性植被类型[3]。属于热带雨林的超地带性扩展形成的独特类型,体现为物种数目减少,特别是乔木种减少,亚热带及温带成分增加,特有的各种热带外貌特征减弱,但本质上又很相似的一种独立的群系类型[4]。国际上关于亚热带雨林的提法和研究也有报道[5, 6, 7, 8],这种植被类型在我国也有出现,主要分布在低海拔沟谷潮湿生境中。在云南称为“亚热带常绿栎林”,在台湾[9, 10, 11]和福建[12, 13, 14, 15]称为“亚热带雨林”,在广东等南亚热带地区常称为“季风常绿阔叶林”[1],宋永昌[16, 17]将这一植被类型称为亚热带适雨常绿阔叶林。
因此,研究井冈山地区的季风常绿阔叶林,对探讨植被区系沿南岭向罗霄山脉地区的扩散或退缩具有重要的生态地理学意义。
1 研究区域自然概况井冈山地区属中亚热带湿润季风气候区。水热条件充沛,年平均气温为17.1℃,最热7月平均气温为23.9℃,7月极端最高气温36.7℃;最冷1月平均气温为3.4℃,1月极端最低气温为-11.0℃。年平均降水量为1889.8mm,最大降水量为2878.8mm。多年平均蒸发量为978.8mm。相对湿度85%。年辐射总量为85—105 MJ/cm2。历年年日照时数约1365.5h。海拔800m以下的土壤主要为山地红壤和山地黄壤。井冈山地貌是岭、沟纵横错节的中山地貌,在保护区核心区,发育有3条主要沟谷流域,为向西南流的行洲河 (河西垄为其上游段)、向西流的湘洲河 (锡坪河为其支流),以及向北流的石溪河 (龙潭为其上游河段)。
井冈山沟谷常绿阔叶林类群包括12个主要群系。本次研究选择分布在海拔650m以下的行洲河和湘洲河流域沟谷低地的5个群丛,以及1个常绿落叶混交林——宜昌润楠群丛作为研究对象。6个样地面积均为1600m2。样地基本情况如表 1所示。
2 调查与数据分析方法[18](1)样地调查
采用常规单株记帐调查法。将每块1600m2样地,划成16个10m×10m方格,进行单株记帐调查,记录树种名称、树高、胸径数据;在每一个10m×10m样方内取得一个2m×2m的小样方,调查乔木幼树及灌木、草本,记录物种、高度和盖度。统计各物种的相对多度(RF),相对频度(RA),相对显著度(RD),相对高度(RH),相对盖度(RC)。其中乔木层重要值(IV)=(RF+ RA+RD)/3,灌草层重要值 (IV) =(RH+ RA+RC)/3。
(2)群落物种多样性分析
分别采用Shannon-Wiener多样性指数 (SW)、Pielou均匀度 (Jsw)和Simpson优势度指数 (λ),计算式如下:
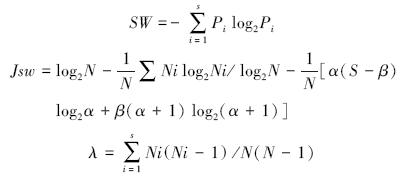
式中,Pi=Ni/N,Ni表示第i个物种的个体数,N表示群落中所有个体数 (或重要值)之和,S表示样方总物种数,β是N被S整除以外的余数 (0≤β<N);α= (N-β)/S。
(3)频度 (R)分析
按Raunkiaer原理,划分为5个等级,即:频度1%—20%为A级,21%—40%为B级,41%—60%为C级,61%—80%为D级,81%—100%为E级。
3 井冈山沟谷季雨林群落的特征 3.1 群落外貌和种类组成表 1是在井冈山沟谷、低地选定的群落概况,按 海拔高度从低到高。根据对6个低地群落的分析, 整体上可以看出井冈山沟谷群落的外貌和组成特点。
| 样地排序/群落名称 Running No./ Community | 郁闭度 Canopy closure | 海拔 Altitude/ m | 蕨类植物 (科/属/种) Ferns (Family/ Genus/ Species) | 种子植物 (科/属/种) Seed plants (Family/Genus/ Species) | 地理位置 Geo coordinates | 样地号/地点 Plot No. /Location |
| P1./杨梅叶蚊母树+米槠群落 Ass. Distylium myricoides +Castanopsis carlesii | 0.80 | 367 | 10/13/14 | 60/91/123 | 26°30′45.44″N 114°13′01.75″E | S5/利洲 |
| P2./鹿角锥+蕈树群落 Ass. Castanopsis lamontii + Altingia chinensis | 0.80 | 376 | 4/5/5 | 35/54/78 | 26°36′01.96″N 114°15′39.64″E | Y5/湘洲 |
| P3./鹿角锥+观光木群落 Ass. Castanopsis lamontii + Michelia odora | 0.85 | 384 | 3/3/5 | 34/68/95 | 26°36′05.93″N 114°15′37.81″E | S21/湘洲 |
| P4./乐昌含笑+钩锥群落 Ass. Michelia chapensis + Castanopsis tibetana | 0.90 | 495 | 6/12/14 | 49/75/104 | 26°35′28.25″N 114°14′34.86″E | Y4/锡坪 |
| P5./大果马蹄荷群落 Ass. Exbucklandia tonkinensis | 0.85 | 618 | 5/5/6 | 45/51/91 | 26°34′53.63″N 114°14′00.40″E | S20/锡坪 |
| P6./宜昌润楠+赤杨叶群落 Ass. Machilus ichangensis + Alniphyllum fortune | 0.75 | 561 | 13/18/22 | 45/70/98 | 26°31′11.22″N 114°10′11.25″E | S10/河西垄 |
(1) 群落外貌
冠层高低起伏明显,呈淡黄绿色、淡绿色至浓绿色,不同季节色相也略有不同,但四季常绿,郁闭度为0.75-0.90。
(2) 乔木层常为多优势种
乔木层常分为2亚层。第一亚层高20—30m,第二亚层14—20m。优势种主要是壳斗科的鹿角锥Castanopsis lamontii、钩锥Castanopsis tibetana、栲Castanopsis fargesii、甜槠Castanopsis eyrei,樟科的宜昌润楠Machilus ichangensis、绒毛润楠Machilus velutina、黄樟Cinnamomum parthenoxylon,杜英科的日本杜英Elaeocarpus japonicus、杜英Elaeocarpus decipiens、猴欢喜Sloanea sinensis,山茶科的厚皮香Ternstroemia gymnanthera,以及金缕梅科的大果马蹄荷Exbucklandia tonkinensis、蕈树Altingia chinensis,木兰科的乐昌含笑Michelia chapensis、观光木Tsoongiodendron odorum,藤黄科的多花山竹子Garcinia multiflora,漆树科的南酸枣Choerospondias axillaris等。上述树种多为泛热带性树种,是群落的特征种,显示一种热带沟谷林性质。
(3) 灌草层较丰富,盖度30%—40%
灌木类以山茶科的山茶属、柃属、杨桐属,樟科的山胡椒属,桃金娘科的赤楠Syzygium buxifolium,紫金牛科的金花树Blastus dunnianus等为常见属、种。草本层,种类较丰富,如狗脊Woodwardia japonica、线蕨Colysis elliptica、珍珠茅Scleria hebecarpa、 草珊瑚Sarcandra glabra、攀援星蕨Microsorium buergerianum、 野苎麻Boehmeria siamensis、蔓赤车Pellionia scabra、无盖鳞毛蕨Dryopteris scottii、狭翅铁角蕨Asplenium wrightii、淡竹叶Lophatherum gracile等。
(4) 层间植物较为丰富
攀援状灌木如番荔枝科的瓜馥木Fissistigma oldhamii在多个沟谷林群落中形成优势层片;常见粗大的木质藤本还有钩藤Uncaria rhynchophylla、野木瓜Stauntonia chinensis、亮叶鸡血藤 Callerya nitida等,其它有常春藤Hedera nepalensis var. sinensis、链珠藤Alyxia sinensis、酸藤子Embelia laeta、菝葜Smilax china、流苏子Coptosapelta diffusa、木通Akebia quinata等。附生小型蕨类也较丰富。林中偶见杜英科、壳斗科等老树板根现象。
3.2 优势科属与重要值分析井冈山沟谷林群落的物种组成丰富,科属的组成复杂,其中乔木层有60科105属222种,从属种的丰富度来说,在6个群落中(表 2)含有5种及以上的科有14科,共53属和150种,占乔木层科、属、种总数的23.33%、50.48%、67.57%。无疑,它们是各群落的优势科,又以樟科、壳斗科、山茶科、冬青科、山矾科、蔷薇科等占较大的比例,也是热带性很强的科[19],主要为泛热带分布的科。从重要值计算看,壳斗科、樟科占较大优势,金缕梅科、山茶科、紫金牛科、安息香科、冬青科等所占比重也较大。
| 主要优势科 Dominant families | P1 | P2 | P3 | P4 | P5 | P6 | Total | 个体数 IN | 重要值 IV | |||||||
| GN | SN | GN | SN | GN | SN | GN | SN | GN | SN | GN | SN | GN | SN | |||
| 所选重要科为6样地中所含种数>5的科,“%” 表示各样地中乔木层主要科的属种之和分别占6样地乔木层所有属(105)种(222)的百分比,GN为属数,SN为种数,IN为个体数,IV为重要值; In the table only shows the family with more than 5 species,“%”mains the percentage of genera/ species of dominant families of every community in the total genera (105) and species (222) of 6 communities. “GN” Number of Genera,“SN” Number of Species,“IN” Individual Number,“IV” Important Value | ||||||||||||||||
| 壳斗Fagaceae | 3 | 9 | 3 | 5 | 3 | 6 | 3 | 8 | 4 | 11 | 3 | 5 | 4 | 18 | 463 | 149.18 |
| 樟科Lauraceae | 4 | 9 | 6 | 10 | 5 | 8 | 3 | 8 | 4 | 11 | 4 | 11 | 7 | 27 | 423 | 114.02 |
| 金缕梅科Hamamelidaceae | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 1 | 5 | 5 | 124 | 93.99 |
| 山茶科Theaceae | 5 | 5 | 4 | 5 | 4 | 6 | 4 | 6 | 6 | 9 | 5 | 9 | 7 | 17 | 295 | 58.65 |
| 紫金牛科Myrsinaceae | 2 | 3 | 2 | 6 | 3 | 3 | 1 | 1 | 3 | 5 | 2 | 2 | 3 | 8 | 106 | 44.31 |
| 安息香科Styracaceae | 3 | 4 | 2 | 2 | 2 | 2 | 1 | 1 | 0 | 0 | 1 | 1 | 3 | 5 | 75 | 39.63 |
| 冬青科Aquifoliaceae | 1 | 4 | 1 | 4 | 1 | 8 | 1 | 5 | 1 | 7 | 1 | 4 | 1 | 15 | 130 | 37.12 |
| 木兰科Magnoliaceae | 1 | 2 | 1 | 3 | 1 | 2 | 2 | 4 | 1 | 3 | 2 | 2 | 2 | 7 | 169 | 36.12 |
| 杜英科Elaeocarpaceae | 1 | 3 | 2 | 4 | 2 | 6 | 1 | 2 | 1 | 2 | 2 | 3 | 2 | 7 | 125 | 29.59 |
| 茜草科Rubiaceae | 6 | 6 | 2 | 2 | 4 | 4 | 3 | 3 | 1 | 1 | 3 | 3 | 7 | 8 | 116 | 21.71 |
| 杜鹃花科Ericaceae | 1 | 3 | 1 | 3 | 1 | 2 | 1 | 1 | 2 | 7 | 1 | 1 | 2 | 9 | 101 | 20.11 |
| 山矾科Symplocaceae | 1 | 3 | 1 | 2 | 1 | 3 | 0 | 0 | 1 | 2 | 1 | 4 | 1 | 10 | 50 | 15.33 |
| 蔷薇科Rosaceae | 3 | 4 | 2 | 3 | 1 | 2 | 2 | 2 | 1 | 2 | 1 | 1 | 5 | 9 | 65 | 14.95 |
| 大戟科Euphorbiaceae | 2 | 2 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 2 | 2 | 4 | 5 | 7 | 2.36 |
| 总计Total | 34 | 58 | 28 | 50 | 30 | 55 | 24 | 44 | 26 | 62 | 29 | 49 | 53 | 150 | 2249 | 677.07 |
| 属种占总数的百分比/% % of the total genera and species | 32.7 | 42.3 | 33 | 51.9 | 42.3 | 55 | 27.6 | 37.3 | 50.5 | 45.6 | 62.6 | 64.1 | 40.8 | 67.6 | 77.6 | 80.6 |
优势种 (或建群种)对说明群落的生境、外貌、组成、演替特征有重要意义[20]。通过对样地数据统计,6个样地合计有357种,其中乔木层有190种,灌草层129种,藤本层39种,分别占总种数的53.2%、36.7%、10.1%;各层相对重要值分别为62.4%、29.3%、8.2%。从表 3表明,重要值IV≥5优势种(除蕈树外)很丰富,达41种,而主要建群种(IV≥10)有20种。同时,可将6样方分为2组,P1、P4、P5为第一组,P2、P3、P6为第二组,而且第一组的建群种其泛热带性特征更强。
| 种类Species | 重要值 (IV)* | |||||||
| P1 | P2 | P3 | P4 | P5 | P6 | Total | % | |
| *表中所列的种类为乔木层中总重要值≥5的树种;Total均包括该种在6个样地的重要值之和;%代表某一物种重要值在6个样地总重要值 (600)的比例 | ||||||||
| 杨梅叶蚊母树Distylium myricoides | 47.83 | 47.83 | 7.98 | |||||
| 鹿角椎Castanopsis lamontii | 15.81 | 23.68 | 1.17 | 40.66 | 6.78 | |||
| 大果马蹄荷Exbucklandia tonkinensis | 1.08 | 34.75 | 35.83 | 5.98 | ||||
| 赤杨叶Alniphyllum fortunei | 0.33 | 1.14 | 3.03 | 12 | 18.13 | 34.63 | 5.78 | |
| 甜槠Castanopsis eyrei | 3.14 | 11.35 | 0.19 | 0.28 | 11.21 | 26.17 | 4.36 | |
| 绒毛润楠Machilus velutina | 3.62 | 5.8 | 2.2 | 3.6 | 9.2 | 24.42 | 4.08 | |
| 乐昌含笑Michelia chapensis | 21.82 | 21.82 | 3.64 | |||||
| 广东山胡椒Lindera kwangtungensis | 3.6 | 18.19 | 21.79 | 3.63 | ||||
| 细枝柃Eurya loquaiana | 0.6 | 0.65 | 6.59 | 1.91 | 3.28 | 5.63 | 18.66 | 3.12 |
| 宜昌润楠Machilus ichangensis | 0.41 | 17.97 | 18.38 | 3.06 | ||||
| 钩锥Castanopsis tibetana | 15.49 | 0.33 | 15.82 | 2.64 | ||||
| 日本杜英Elaeocarpus japonicus | 1.44 | 2.02 | 3.63 | 0.65 | 3 | 4.48 | 15.22 | 2.54 |
| 赤楠Syzygium buxifolium | 4.14 | 6.52 | 1.36 | 0.28 | 0.83 | 0.72 | 13.85 | 2.31 |
| 格药柃Eurya muricata | 1.97 | 1.34 | 1.94 | 0.87 | 7.1 | 13.22 | 2.20 | |
| 树参Dendropanax dentiger | 12.65 | 12.65 | 2.11 | |||||
| 黄丹木姜子Litsea elongata | 0.45 | 4.87 | 1.34 | 0.34 | 3.55 | 2.04 | 12.59 | 2.10 |
| 栲Castanopsis fargesii | 3.82 | 4.34 | 0.37 | 3.13 | 11.66 | 1.94 | ||
| 杉木Cunninghamia lanceolata | 0.27 | 2.48 | 1.44 | 1.51 | 2.78 | 3.08 | 11.56 | 1.92 |
| 香楠Aidia canthioides | 2.92 | 1.9 | 0.67 | 4.11 | 1.54 | 11.14 | 1.86 | |
| 米槠Castanopsis carlesii | 10.97 | 10.97 | 1.82 | |||||
| 罗浮栲Castanopsis fabri | 0.44 | 2.81 | 3.82 | 2.9 | 9.97 | 1.66 | ||
| 密花山矾Symplocos congesta | 2.9 | 7.03 | 9.93 | 1.66 | ||||
| 木姜叶柯Lithocarpus litseifolius | 0.69 | 1.93 | 2.95 | 4.21 | 9.78 | 1.64 | ||
| 毛冬青Ilex pubescens | 2.95 | 0.19 | 3.26 | 1.3 | 1.52 | 0.52 | 9.74 | 1.62 |
| 南酸枣Choerospondias axillaris | 0.34 | 4.1 | 5.23 | 9.67 | 1.61 | |||
| 多花山竹子Garcinia multiflora | 4.27 | 1.4 | 3.77 | 9.44 | 1.58 | |||
| 台湾冬青Ilex formosana | 0.41 | 2.65 | 3.21 | 0.57 | 2.24 | 9.08 | 1.52 | |
| 鼠刺Itea chinensis | 0.38 | 6.05 | 0.52 | 1.05 | 8 | 1.34 | ||
| 深山含笑Michelia maudiae | 0.22 | 4.05 | 0.22 | 0.68 | 1.48 | 0.97 | 7.62 | 1.28 |
| 红楠Machilus thunbergii | 0.51 | 0.19 | 1.27 | 3.19 | 1.53 | 6.69 | 1.12 | |
| 厚皮香Ternstroemia gymnanthera | 0.97 | 3.4 | 0.82 | 0.29 | 0.82 | 0.23 | 6.53 | 1.08 |
| 枫香Liquidambar formosana | 3.1 | 1.87 | 0.48 | 0.83 | 6.28 | 1.04 | ||
| 虎皮楠Daphniphyllum oldhamii | 1.17 | 1.8 | 2.56 | 0.25 | 0.29 | 6.07 | 1.02 | |
| 罗浮柿Diospyros morrisiana | 0.69 | 4.55 | 0.52 | 5.76 | 0.96 | |||
| 密花树Myrsine seguinii | 0.2 | 4.7 | 0.16 | 0.59 | 5.65 | 0.94 | ||
| 杨桐Adinandra millettii | 0.19 | 1.29 | 1.87 | 0.88 | 1.36 | 5.59 | 0.93 | |
| 马银花Rhododendron bachii | 0.45 | 3.08 | 0.5 | 1.1 | 0.26 | 5.39 | 0.90 | |
| 杜英Elaeocarpus decipiens | 2.63 | 2.07 | 0.6 | 5.30 | 0.88 | |||
| 黄樟Cinnamomum parthenoxylon | 0.63 | 2.33 | 0.57 | 1.75 | 5.28 | 0.88 | ||
| 云山青冈Cyclobalanopsis sessilifolia | 1.24 | 4.03 | 5.27 | 0.88 | ||||
| 桃叶石楠Photinia prunifolia | 1.94 | 1.15 | 0.43 | 0.89 | 0.74 | 5.15 | 0.86 | |
| 蕈树Altingia chinensis | 3.81 | 3.81 | 0.64 | |||||
在6个样地中许多藤本植物属于热带、亚热带分布属(表 4),如钩藤、藤黄檀、菝葜、瓜馥木等为泛热带分布属,网脉酸藤子等为旧世界热带属,链珠藤为热带亚洲至大洋洲分布属,雀梅藤为热带亚洲至热带美洲间断分布属,常春藤为热带亚洲至热带非洲分布属[21, 22]。
| 种名Species | 重要值IV | |||||||
| P1 | P2 | P3 | P4 | P5 | P6 | Total | % | |
| 表中所列为重要值大于1的物种;%代表某一物种在6个样地重要值之和占6个样地中总物种重要值之和 (600)的百分比 | ||||||||
| 瓜馥木Fissistigma oldhamii | 3.43 | 1.7 | 7.97 | 9 | 22.1 | 3.6 | ||
| 链珠藤Alyxia sinensis | 0.62 | 7.5 | 1.03 | 0.7 | 9.85 | 1.6 | ||
| 菝葜Smilax china | 6.09 | 1.21 | 0.64 | 1.11 | 9.05 | 1.6 | ||
| 雀梅藤Sageretia thea | 3.85 | 4.03 | 7.88 | 1.4 | ||||
| 常春藤Hedera nepalensis var. sinensis | 0.59 | 0.37 | 4.84 | 5.8 | 1 | |||
| 密齿酸藤子Embelia vestita | 3.93 | 1 | 0.84 | 5.77 | 1 | |||
| 藤黄檀Dalbergia hancei | 1.17 | 0.59 | 1.95 | 3.71 | 0.6 | |||
| 钩藤Uncaria rhynchophylla | 3.32 | 3.32 | 0.6 | |||||
| 酸藤子Embelia laeta | 3.02 | 3.02 | 0.6 | |||||
| 三叶崖爬藤Tetrastigma hemsleyanum | 2.56 | 2.56 | 0.4 | |||||
| 流苏子Coptosapelta diffusa | 1.06 | 0.67 | 0.82 | 2.55 | 0.4 | |||
| 千金藤Stephania japonica | 0.19 | 1.98 | 2.17 | 0.4 | ||||
| 木通Akebia quinata | 0.73 | 0.91 | 0.37 | 2.01 | 0.4 | |||
| 土茯苓Smilax glabra | 0.82 | 1.07 | 1.89 | 0.4 | ||||
| 粗叶悬钩子Rubus alceifolius | 1.62 | 1.62 | 0.2 | |||||
| 亮叶鸡血藤Callerya nitida | 1.6 | 1.6 | 0.2 | |||||
| 圆苞杜根藤Justicia championii | 1.43 | 1.43 | 0.2 | |||||
| 野木瓜Stauntonia chinensis | 0.19 | 1.12 | 1.31 | 0.2 | ||||
| 崖爬藤Tetrastigma obtectum | 1.31 | 1.31 | 0.2 | |||||
| 暗色菝葜Smilax lanceifolia var. opaca | 0.53 | 0.64 | 1.17 | 0.2 | ||||
| 酸味子Antidesma japonicum | 0.66 | 0.39 | 1.05 | 0.2 | ||||
| 玉叶金花Mussaenda pubescens | 0.63 | 0.39 | 1.02 | 0.2 | ||||
| 清香藤Jasminum lanceolarium | 1.01 | 1.01 | 0.2 | |||||
| 锈毛莓Rubus reflexus | 1 | 1 | 0.2 | |||||
统计表明,6个样地合计藤本植物36种中,粗大木质藤本共26种,如野木瓜、木通、鸡血藤、钩藤、瓜馥木等,占72.2%,草质藤本为10种,占27.8%。整体来看,热带性的藤本植物属,与南亚热带沟谷林相当。相对而言,样地P1、P4、P6中,藤本的丰度、热带性较其它3个样地更强,又以样地P6的热带性最丰富,含6种。
3.5 群落草本植物与重要值分析统计6个样地共有草本植物93种,其中蕨类植物43种(表 5),在多个样地中同时出现的主要种类有草珊瑚,狗脊,宽叶薹草,鳞毛蕨,淡竹叶,赤车,凤丫蕨,江南卷柏,石韦,阴地蕨,冷水花等,按草本层重要值排序,重要值最大的为狗脊(36.98),其次为草珊瑚(17.9)、鳞毛蕨(13.95)、凤丫蕨(10.37)、冷水花(9.59)、江南卷柏(9.56)、福建观音莲座(9.23)、芒萁(8.35)、披针骨牌蕨(7.84)、攀援星蕨(6.83)、里白(6.01)、珍珠茅(5.58)、深绿卷柏(5.34)、线蕨(5.15)。其中,草本层中多为喜温喜湿种,主要为蕨类植物,其以泛热带和热带亚洲分布为多,如狗脊、凤丫蕨、福建观音莲座等,其中,粗齿黑桫椤等为热带林常见的大型树蕨,其他如鳞毛蕨、江南卷柏等为世界广布种[23, 24]。另外,冷水花也为泛热带分布,草珊瑚、淡竹叶、赤车等为亚热带分布[20]。
| 种名Species | 重要值IV | |||||||
| P1 | P2 | P3 | P4 | P5 | P6 | Total | % | |
| 表中所列为重要值大于3的物种;%代表某物种在6个样地中重要值之和占6个样地中总物种重要值之和(600)的百分比 | ||||||||
| 狗脊蕨Woodwardia japonica | 6.03 | 2.2 | 11.6 | 5.2 | 9.71 | 2.24 | 36.98 | 6.2 |
| 草珊瑚Sarcandra glabra | 6 | 1.34 | 4.83 | 2.39 | 1.99 | 1.35 | 17.9 | 3 |
| 中华鳞毛蕨Dryopteris chinensis | 7.08 | 1.07 | 1.06 | 0.42 | 5.38 | 15.01 | 2.6 | |
| 凤丫蕨Coniogramme japonica | 8.42 | 0.64 | 1.31 | 10.37 | 1.8 | |||
| 冷水花Pilea notata | 0.14 | 9.45 | 9.59 | 1.6 | ||||
| 江南卷柏Selaginella moellendorfii | 0.69 | 7.89 | 0.98 | 9.56 | 1.6 | |||
| 福建莲座蕨Angiopteris fokiensis | 8.07 | 1.16 | 9.23 | 1.6 | ||||
| 芒萁Dicranopteris pedata | 8.35 | 8.35 | 1.4 | |||||
| 披针叶骨牌蕨Lepidogrammitis diversa | 7.84 | 7.84 | 1.4 | |||||
| 攀援星蕨Microsorum brachylepis | 5.97 | 0.86 | 6.83 | 1.2 | ||||
| 里白Diplopterygium glaucum | 6.01 | 6.01 | 1 | |||||
| 黑鳞珍珠茅Scleria hookeriana | 0.52 | 5.06 | 5.58 | 1 | ||||
| 深绿卷柏Selaginella doederleinii | 5.34 | 5.34 | 0.8 | |||||
| 线蕨Colysis elliptica | 5.15 | 5.15 | 0.8 | |||||
| 石韦Pyrrosia lingua | 2.15 | 0.99 | 1.74 | 4.88 | 0.8 | |||
| 扯根菜Penthorum chinense | 4.84 | 4.84 | 0.8 | |||||
| 楼梯草Elatostema involucratum | 4.25 | 4.25 | 0.8 | |||||
| 蔓赤车Pellionia scabra | 2.62 | 1.6 | 4.22 | 0.8 | ||||
| 卵叶盾蕨Neolepisorus ovatus | 0.55 | 3.61 | 4.16 | 0.6 | ||||
| 水龙骨Polypodiodes nipponica | 0.32 | 3.64 | 3.96 | 0.6 | ||||
| 淡竹叶Lophatherum gracile | 0.25 | 2.57 | 0.73 | 3.55 | 0.6 | |||
| 赤车Pellionia radicans | 0.6 | 1.4 | 1.36 | 3.36 | 0.6 | |||
| 中华里白Diplopterygium chinense | 3.34 | 3.34 | 0.6 | |||||
| 扁穗莎草Cyperus compressus | 0.97 | 2.34 | 3.31 | 0.6 | ||||
| 星毛冠盖藤Pileostegia tomentella | 3.09 | 3.09 | 0.6 | |||||
针对各群落的物种组成,按吴征镒所划分的15个分布区类型,分析种子植物属的地理成分[21, 22, 25]。结果表明(表 6),井冈山沟谷林群落均以热带性成分占绝对优势,主要优势属:蚊母树属、马蹄荷属、含笑属、柃木属、树参属、安息香属、杜英属、山矾属、紫金牛属、冷水花属等均为泛热带分布,也有部分温带区系成分的侵入[26, 27],如栲属、鼠刺属等。其热带性成分达64.71%—77.94%,较之海南尖峰岭的2个热带雨林群落[28]的热带性成分83.15%—85.00%稍低,但两个区域相差8个纬度,热带成分仅高5.21%。而与南亚热带季风气候区的鼎湖山[29]比较,热带性成分76.38%相接近,甚至略高。与三清山[30]的典型亚热带常绿阔叶林比较,明显具有较高的热带性成分。王希华[31]对中国典型常绿阔叶林的分析表明,其热带性成分占56.05%,温带性成分占43.95%(含中国特有成分6.37%)。从井冈山沟谷林看,具有从热带雨林向亚热带常绿阔叶林过渡的南亚热带区系特点,与亚热带地带性的典型常绿阔叶林植被有明显区别[26]。
| 分布区类型 Areal types | 尖峰 岭[28] | 鼎湖 山[29] | 井冈山,江西 | 三清 山[30] | 典型常绿 阔叶[31] Typical EBLF | ||||||
| P1 | P2 | P3 | P4 | P5 | P6 | ||||||
| -: 比例中扣除广布种 | |||||||||||
| 1.广布 Cosmopolitan | |||||||||||
| 2. 泛热带Pan tropic | 21.35 | 18.33 | 28 | 28.57 | 26.47 | 28.09 | 21.43 | 21.57 | 23.38 | 7.69 | 17.85 |
| 3. 东亚及热带南美间断 Trop. Asia & Trop. Amer. Disjuncted | 6.74 | 6.67 | 4.4 | 3.36 | 8.82 | 10.11 | 6.12 | 3.92 | 6.49 | 15.38 | 2.5 |
| 4. 旧世界热带Old world tropics | 11.24 | 11.67 | 10.64 | 10.08 | 8.82 | 4.49 | 8.16 | 7.84 | 6.49 | 3.85 | 6.87 |
| 5. 热带亚洲至热带大洋洲 Trop.Asia to Trop. Australasia | 8.99 | 8.33 | 8.95 | 4.2 | 2.94 | 13.48 | 6.12 | 1.96 | 6.49 | 3.85 | 4.49 |
| 6. 热带亚洲至热带非洲 Trop. Asia to Trop. Africa | 4.49 | 5 | 6.74 | 4.2 | 4.41 | 0 | 4.08 | 1.96 | 7.79 | 0 | 4.74 |
| 7. 热带亚洲 Trop.Asia (Indo-Malaysia) | 30.34 | 35 | 17.64 | 21.85 | 26.47 | 15.73 | 24.49 | 27.45 | 25.97 | 19.23 | 19.6 |
| 8. 北温带North Temperate | 3.25 | 3.33 | 7.9 | 10.08 | 7.35 | 5.62 | 6.12 | 11.76 | 10.39 | 7.69 | 12.73 |
| 9. 东亚及北美间断 E. Asia & N. Amer. Disjuncted | 8.99 | 8.33 | 3.5 | 9.24 | 5.88 | 14.61 | 12.24 | 11.76 | 9.09 | 26.92 | 7.24 |
| 10. 旧世界北温带 Old World temperate | 0 | 0 | 2.9 | 0 | 1.47 | 0 | 0 | 1.96 | 0 | 0 | 2 |
| 11. 温带非洲 Temp. | 0 | 0 | 0.5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.75 |
| 12. 地中海区、西亚至中亚 Mediterranea W. Asia to C. Asia | 1.12 | 1.67 | 0.13 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.37 |
| 13. 中亚C. Asia | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 14. 东亚E. Asia | 2.25 | 1.67 | 6.6 | 5.88 | 5.88 | 5.62 | 10.2 | 7.84 | 2.6 | 15.38 | 14.48 |
| 15. 中国特有Endemic to China | 2.25 | 0 | 2.1 | 2.52 | 1.47 | 2.25 | 1.02 | 1.96 | 1.3 | 0 | 6.37 |
| 热带分布总计Total in Tropic | 83.15 | 85 | 76.38 | 72.27 | 77.94 | 71.91 | 70.41 | 64.71 | 76.62 | 50 | 56.05 |
| 温带分布总计Total in Temperate | 16.85 | 15 | 23.62 | 27.73 | 22.06 | 28.09 | 29.59 | 35.29 | 23.38 | 50 | 43.95 |
物种多样性指数、生态优势度、群落均匀度,可以从3个不同的角度表征群落组成和结构水平[32]。表 7表明,井冈山沟谷群落的Shannon-Wiener多样性指数为4.44—5.46,略低于海南岛尖峰岭的3个典型热带山地雨林,后者为5.02—6.05。与其它热带山地雨林群落[28, 33, 34]相比,其Shannon-Wiener多样性指数也稍低。与鼎湖山典型的南亚热带季风常绿阔叶林[32, 35] (4.12—4.57)、福建南靖的亚热带雨林[36] (4.55)相比,其物种多样性指数接近,甚至更高;而比三清山典型的亚热带常绿阔叶林群落[30, 37]的物种多样性指数 (2.77—4.2)要高很多。另外,井冈山沟谷群落的生态优势度和群落均匀度数据分别为0.04—0.102和0.73—0.86。整体来说,井冈山沟谷低地群落的物种多样性指数、生态优势度和群落均匀度指标,均与广东鼎湖山季风常绿林和福建南靖南亚热带雨林群落相一致,其群落的组成结构水平相似;且远超过同纬度或相近纬度中亚热带地区的其它独立山体,个别甚至与热带地区的常绿阔叶林群落相似。
| 地点 Site | 群落名称 Names of association | 面积Area/ m2 | 种数 Species numbers | 物种多样 性指数 Species diversity index | 生态优势 度指数 Ecological dominance index | 均匀度指数 Community evenness index |
| 尖峰岭,海南 | 倒卵阿丁枫+丛花厚壳桂+托盘青冈群落Ass. Altingia chinensis+Cryptocarya densiflora + Cyclobalanopsis patelliformis | 2500 | 125 | 5.92 | 0.025 | 0.85 |
| 小叶白椎+海南黄叶树+紫树群落Ass. Castanopsis fabri+Xanthophyllum hainanense+Nyssa sinensis | 2500 | 120 | 6.05 | 0.044 | 0.88 | |
| 藜蒴+米花木+灰木群落Ass. Castanopsis fissa+Saurauia tristyla + Symplocos sumuntia | 2500 | 71 | 5.02 | 0.082 | 0.82 | |
| 鼎湖山,广东 | 锥栗+黄果厚壳桂群落Ass. Castanea enryi+Cryptocarya concinna | 1200 | 71 | 4.57 | 0.078 | 0.79 |
| 藜蒴+厚壳桂群落Ass. Castanopsis fissa+Cryptocarya chinensis | 1200 | 61 | 4.12 | 0.086 | 0.69 | |
| 南靖,福建 | 红栲群落Ass. Castanopsis fargesii | 3000 | - | 4.55 | 0.81 | |
| 井冈山,江西 | P1 | 1600 | 137 | 5.46 | 0.102 | 0.83 |
| P2 | 1600 | 83 | 5.2 | 0.040 | 0.86 | |
| P3 | 1600 | 100 | 4.86 | 0.043 | 0.81 | |
| P4 | 1600 | 118 | 4.48 | 0.076 | 0.74 | |
| P5 | 1600 | 97 | 4.92 | 0.096 | 0.82 | |
| P6 | 1600 | 120 | 4.44 | 0.066 | 0.73 | |
| 三清山,江西 | 钩锥+山腊梅群落Ass. Castanopsis tibetana +Chimonanthus nitens | 1200 | 82 | 2.77 | - | 0.86 |
| 长柄双花木群落Ass. Disanthus cercidifolius subsp. longipes | 1200 | 56 | 4.2 | - | 0.79 |
频度 (Frequency)表示某一种群的个体在群落中水平分布的均匀程度[18]。按照Raunkiaer的5个频度等级对井冈山沟谷低地群落进行频度分析(图 1)。

|
| 图 1 井冈山典型群落的频度分析 Fig. 1 Frequency analysis of the ravine communities in Mount Jinggangshan |
(1)各频度级关系为:A>B>C>D<E
由图 1可知,群落P1、P4、P6的频度级关系与Raunkiaer频度规律 (A>B>C≥D<E)相似。由于群落中组成种群较多,大量偶见种的出现使A级物种比例明显增大,D、E两级比例显著缩小,甚至接近于0。与尖峰岭热带山地雨林的原始林群落的频度级关系相似[28]。
(2)各频度级关系为:A>B>C>D>E
由图 1可知,群落P2、P3、P5的频度级关系与Raunkiaer频度规律A>B>C≥D<E有所不同,但与海南岛热带雨林的频度分布规律A>B>C>D>E[38]仍相似。在群落中频度级为A级的物种所占比例很大,这一现象与单种属及扩散种较丰富有关,说明群落中偶见种较多,来源丰富。E级物种数少则从一定程度上反映了群落的物种分布不甚均匀,优势种与建群种相对较明显。
3.9 其它生态特征在井冈山沟谷林群落中,层间植物较丰富。主要有附生的兰花、小型的蕨类和藤本,其中藤本以灌木状藤本占优势,如藤黄檀、瓜馥木、酒饼叶等;也有部分中、大型木质藤本,如木通、野木瓜、钩藤、鸡血藤、拓树、秤钩风等,高达10m以上,径可达15cm,木质茎在层间弯曲盘绕,总长度可达30m以上。
群落中的板根现象常有出现。主要见于栲、杜英、木莲等高大乔木,通常为长、高均小于1.5m的中小型板根。偶见较大的板根可在地面延伸2—3m,高达1.5m。
群落种类组成显示着起源上的古老性。建群种大果马蹄荷起源于第三纪,山胡椒属 (山橿)、樟属 (香桂、辣汁樟)、柯属 (东南石栎、滑皮柯、甜茶稠)、山茶属 (野茶树)、槭属 (青榨槭、长柄槭、五裂槭)等也是第三纪残遗成分。
4 结论与讨论(1)井冈山亚热带沟谷季雨林群落形成的原因
井冈山沟谷、低地季雨林各群落的物种组成丰富,各群落间随海拔高度的上升,物种丰富度稍有减少。热带性的优势科属、物种多样性指数、生态优势度、群落均匀度等,均远超同纬度或相近纬度地区的独立山体,而与南亚热带地区(北回归线地区)相似,甚至更丰富,还有其他一些重要的热带性雨林特征,如:群落区系起源古老,大型木质藤本、板根现象、寄生、附生以及兰科植物均较丰富。
井冈山地处中亚热带南缘,地质地貌复杂多样,相对海拔高差近1900m,导致热能和水分在时空上有明显差异,形成了中-南亚热带,中亚热带,北亚热带和暖温带等4个垂直气候亚带,发育出丰富的土壤和植被类型。受地貌和水文条件等的影响,在井冈山西南流向的湘洲河和行洲河的低海拔沟谷地段形成了温暖潮湿的沟谷环境,具有避风、湿度大和热量足的特点,土壤为具有丰富有机质的山地红壤或黄壤,为某些热带区系成分的生存提供了有利条件,从而发育形成了具有南亚热带性质的亚热带沟谷季雨林。
(2)井冈山地区沟谷季雨林群落的生态地理学意义
井冈山保存的亚热带沟谷季雨林是“印度-马来界”植被和群落类型向北延伸的“飞地”。在演替上常被称为侵入群落[2]或孑遗群落,这一类型显然是超地带性的,在植被或区系演替上具有重要意义。整体上,井冈山保存有冰期以来亚洲东部地区最为丰富完整的生物区系、生物多样性,是反映陆地生态系统、生物群落在冰期自“自北向南”退缩,以及间冰期“自南向北”重新扩张等重大地质历史演化事件和重要生态发育过程的重要例证。也是各类珍稀濒危物种、孑遗种,各类原始的维管植物,以及区域特有种、中国特有种的天然避难所。这也冲破了通道县(为湖南、贵州、广西三省交界)南部亚热带雨林为亚热带雨林的最北缘类型的界限[15],对研究我国植被的分布、类型和演替都有重要的价值。
| [1] | Wu Z Y. Chinese Vegetation, Beijing: Science Press, 1980: 1-1144. |
| [2] | Lin Y. Discussion of the vegetation types of Nanling Mountain and its position in the Chinese vegetation regionalization. Acta Phytoecologica et Geobotanica Sinica, 1965, 3(1): 65-67. |
| [3] | Wang B S. Discussion of the level regionalization of monsoon forests. Acta Phytoecologica et Geobotanica Sinica, 1987, 11(2): 154-158. |
| [4] | Zhang H D. Tropical Rainforest (Richards P G). Beijing: Science Press, 1959: 1-534. |
| [5] | Alexanderson C, Carlos A D. Ecology of the elfin woods warbler (Dendroica angelae). I. Distribution, habitat usage, and population densities. Caribbean Journal of Science, 1984, 20(1/2): 89-96. |
| [6] | Dodson J R, Myers C A. Vegetation and modern pollen rain from the Barrington Tops and Upper Hunter River Regions of New South Wales. Australian Journal of Botany, 1986, 34: 293-304. |
| [7] | Laidlaw M, Olsen M, Kitching R L, et al. Tree floristic and structural characteristics of one hectare of subtropical rainforest in Lamington National Park, Queensland. Proceedings of the Royal Society of Queensland, 2000, 109: 91-105. |
| [8] | Menzel F, Kitching RL, Sarah L. Boulter S.L. Host specificity or habitat structure? The epicortical beetle assemblages in an Australian subtropical rainforest. European Journal of Entomology, 2004, 101 (2): 251-259. |
| [9] | Chao W C, Chao K J, Guo-Zhang M. Song and Chang-Fu Hsieh. Species composition and structure of the lowland subtropical rainforest at Lanjenchi, Southern Taiwan. Taiwania, 2007, 52 (3): 253-269. |
| [10] | Chao W C, Wu S H, Lin H Y. Distribution patterns of tree species in the Lanjenchi lowland rain forest, Taiwania, 2007, 52 (4): 343-351. |
| [11] | Huang W L. Vegetation type classification system of Taiwan islands. Guizhou Science, 2003, 21 (1/2): 40-44. |
| [12] | Lin P, Qiu X Z. Subtropical rainforest of Hexi in Nanjing County, Fujian Province, China. Acta Phytoecologica et Geobotanica Sinica, 1987, 11 (3): 161-170. |
| [13] | Wei S Z. Discussion on subtropical rain forest of Letu, Nabjing County, Fujian Province. Journal of Fujian teacher's university (Natural Science), 1988, 4(2): 100-107. |
| [14] | He J. Discussion of the subtropical rain forests of southeast China, starting at the found of subtropical rain forest of Hexi, Nabjing County, Fujian Province. Journal of Xiamen University, 1955, 5: 31-34. |
| [15] | Duan W X. The subtropical rain forest and its reasons for the formation in the south of Tongdao county. Natural Science Journal of Huaihua Teacher's College, 1989, 8 (1): 70-73. |
| [16] | Song Y C. Tentative classification scheme of evergreen broad- leaved forests of China. Acta Phytoecologica Sinica, 2004, 28 (4): 435-448. |
| [17] | Song Y C, Xu G S, Chen W L, Wang X H, Da L J, Chen T C. Evergreen broad- leaved forest in Taiwan and its relationship with counterparts in mainland China. Acta Phytoecologica Sinica, 2003, 27(6): 719-732. |
| [18] | Wang B S, Yu S X, Peng S L. Nature Protocols of Phytocoenology. Guangzhou: Guangdong Higher Education Press, 1996:23-105. |
| [19] | Wu Z Y, Zhou Z K, Li D Z. Peng H, Sun H. The Areal-types of the world families of seed plants, Acta Botanica Yunnanica, 2003, 25 (3): 245-257. |
| [20] | Lin P. Phytosociology. Shanghai: Shanghai Scientific and Technology Press, 1986: 178-190. |
| [21] | Wu Z Y. The areal- types of Chinese genera of seed plants. Acta Botanica Yunnanica, 1991, 1: 1-139. |
| [22] | Wu Z Y. The areal- types of Chinese genera of seed plants. Acta Botanica Yunnanica, 1991, 1: 141-178. |
| [23] | Lu S G. Floristic of Pteridophytes in China. Flora of China. Beijing: Sciense Press, 2004: 80-94. |
| [24] | Lu S G. Floristic Outline of Pteridophytes in China. Advances in plant sciences. Beijing: Higher Education Press, 2004, 6: 29-41. |
| [25] | Wu Z Y, Zhou Z K, Sun H,Li D Z, Peng H. The Areal- types of Seed Plants and their Origin and Differentiation. Kunming: Yunnan Science and Technology Press, 2006: 146-451. |
| [26] | Lin Y. Scientific Survey and Research on Jinggangshan Nature Reserve. Beijing: Xinhua Publishing House, 1990: 1-544. |
| [27] | Liu R L, Tang G C. Study on flora of spermatophyte in Jinggang Mountain. Journal of Wuhan botanical research, 1995, 13 (3): 210-218. |
| [28] | Li Y D. Community characteristics of Tropical mountain rain forest in Jianfengling, Hainan island. Journal of Tropical and Subtropical botany, 1997, 5(1): 18-26. |
| [29] | Chen B Y, Li Z X, Zhang G C. Floristic analysis of Dinghu Shan. Research of Tropical and Subtropical forest ecological system, 1982, 1:48-57. |
| [30] | Shen R J, Lin S S, Fan Q, Jin J H, Liao W B, Zhong F H. Study on Dominant Community of Disanthus cercidifolius subsp.longipes from Mountain Sanqingshan of Jiangxi Province. Journal of Wuhan botanical research, 2009, 27(5): 501-508. |
| [31] | Wang X H. The Phytogeography and Species Diversity of Typical Evergreen Broad-leaved Forest in China[D]. Shanghai: East China Normal University, 2006, 1-121. |
| [32] | Peng S L, Zhou H C, Chen T X. The quantitative characters of organization of forest communities in Guangdong, Acta Phytoecologica et. Geobotanica Sinica, 1989, 13 (1): 10-17. |
| [33] | An S Q, Zhu X L, Wang Z F,Campbell D G, Li G Q, Chen X L. The plant species diversity in a tropical montane rain forest on Wuzhi Mountain, Hainan, Acta Ecologica Sinica, 1999, 19 (6): 803-809. |
| [34] | Fang J Y, Li Y D, Zhu B, Liu G H, Zhou G Y. Community structures and species richness in the montane rain forest of Jianfengling, Hainan Island, China. Biodiversity Science, 2004, 12 (1): 29-43. |
| [35] | Huang Z L, Kong G H, He D Q. Plant community diversity in Dinghushan Nature Reserve. Acta Ecologica Sinica, 2000, 20 (2): 193-198. |
| [36] | Lian Y W, Zhang Y H, Zhu X L, Xu J S, Li Z J. Analysis on species diversity and succession trend of subtropical rainforest in Hexi, Fujian. Journal of Xiamen University (Natural Science), 2000, 39 (3): 392-399. |
| [37] | Guo W, Shen R J, Wu J H,Liao W B, Peng S L, Chen H. Analysis on community composition and structure of Pseudotsuga gaussenii in Sanqing Mountain of Jiangxi Province. Journal of plant resources and environment, 2007, 16 (3): 46-52. |
| [38] | Hu Y J, Li Y X. Tropical Rain Forests of Hainan Island. Guangzhou: Guangdong Higher Education Press. 1992: 1-29. |
| [1] | 吴征镒主编. 中国植被. 北京: 科学出版社, 1980: 1-1144. |
| [2] | 林英. 论南岭山地植被的性质及其在中国植被区划中的位置问题. 植物生态学与地植物学丛刊, 1965, 3(1): 65-67. |
| [3] | 王伯荪. 论季雨林的水平地带性. 植物生态学与地植物学学报, 1987, 11(2): 154-158. |
| [4] | 理查斯. 热带雨林//张宏达等, 译. 北京: 科学出版社, 1959: 1-534. |
| [11] | 黄威廉. 台湾植被类型分类系统. 贵州科学, 2003, 21 (1/2): 40-44. |
| [12] | 林鹏, 邱喜昭. 福建南靖县和溪的亚热带雨林. 植物生态学与地植物学报, 1987, 11 (3): 161-170. |
| [13] | 魏守珍. 福建省南靖乐土亚热带雨林探讨. 福建师范大学学报(自然科学版), 1988, 2. |
| [14] | 何景. 从福建南靖县和溪镇“雨林”的发现谈到我国东南亚热带雨林区. 厦门大学学报, 1955, 5: 31-34. |
| [15] | 段帷泞. 通道县南部亚热带雨林及其成因. 怀化师专自然科学学报, 1989, 8 (1): 70-73. |
| [16] | 宋永昌. 中国常绿阔叶林分类试行方案. 植物生态学报, 2004, 28 (4): 435-448. |
| [17] | 宋永昌, 徐国士, 陈伟烈, 王希华, 达良俊, 陈添财. 台湾常绿阔叶林主要类型及其与大陆常绿阔叶林的关系. 植物生态学报. 2003, 27(6): 719-732. |
| [18] | 王伯荪, 余世孝, 彭少麟.植物群落学实验手册. 广州: 广东高等教育出版社, 1996:23-105. |
| [19] | 吴征镒, 周浙昆, 李德铢,彭华, 孙航. 世界种子植物科的分布区类型系统. 云南植物研究, 2003, 25 (3): 245-257. |
| [20] | 林鹏. 植物群落学. 上海: 上海科学出版社, 1986: 178-190. |
| [21] | 吴征镒. 中国种子植物属的分布区类型. 云南植物研究, 1991, 1 (增刊): 1-139. |
| [22] | 吴征镒. 中国种子植物属的分布区类型. 云南植物研究, 1991, 1 (增刊): 141-178. |
| [23] | 陆树刚. 中国蕨类植物区系. 中国植物志 (第1卷) 北京: 科学出版社, 2004: 80-94. |
| [24] | 陆树刚. 中国蕨类植物区系概论. 植物科学进展. 北京: 高等教育出版社, 2004, 6: 29-41. |
| [25] | 吴征镒, 周浙昆, 孙航, 李德铢, 彭华.中国种子植物属的分布区类型及其起源和分化. 昆明: 云南科技出版社, 2006: 146-451. |
| [26] | 林英. 井冈山自然保护区考察研究. 北京: 新华出版, 1990: 1-544. |
| [27] | 刘仁林, 唐赣成. 井冈山种子植物区系的研究. 武汉植物学研究, 1995, 13 (3): 210-218. |
| [28] | 李意德. 海南岛尖峰岭热带山地雨林的群落结构特征. 热带亚热带植物学报, 1997, 5(1): 18-26. |
| [29] | 陈邦余, 李泽贤, 张桂才. 鼎湖山植物区系的探讨. 热带亚热带森林生态系统研究, 1982, 1:48-57. |
| [30] | 沈如江, 林石狮, 凡强, 金建华, 廖文波, 钟方华,方华华. 江西省三清山长柄双花木优势群落研究. 武汉植物学研究, 2009, 27(5): 501-508. |
| [31] | 王希华. 中国典型常绿阔叶林植物地理与物种多样性研究[D]. 上海: 华东师范大学, 2006. |
| [32] | 彭少麟, 周厚诚, 陈天杏. 广东森林群落的组成结构数量特征. 植物生态学与地植物学学报, 1989, 13 (1): 10-17. |
| [33] | 安树青, 朱学雷, 王峥峰, David G. Campbell, 李国旗, 陈兴龙. 海南五指山热带山地雨林植物物种多样性研究. 生态学报, 1999, 19 (6): 803-809. |
| [34] | 方精云, 李意德, 朱彪, 刘国华, 周光义. 海南岛尖峰岭山地雨林的群落结构、物种多样性以及在世界雨林中的地位. 生物多样性, 2004, 12 (1): 29-43. |
| [35] | 黄忠良, 孔国辉, 何道泉. 鼎湖山植物群落多样性的研究. 生态学报, 2000, 20 (2): 193-198. |
| [36] | 连玉武,张宜辉, 朱小龙, 徐金森, 李振基. 福建南亚热带雨林物种多样性与群落演替趋势分析. 厦门大学学报 (自然科学版), 2000, 39 (3): 392-399. |
| [37] | 郭微, 沈如江, 吴金火, 廖文波, 彭少麟, 陈晖. 江西三清山华东黄杉群落的组成及结构分析. 植物资源与环境学报, 2007, 16 (3): 46-52. |
| [38] | 胡玉佳, 李玉杏. 海南岛热带雨林研究. 广州: 广东高等教育出版社, 1992: 1-29. |