 2014, Vol. 34
2014, Vol. 34文章信息
- 金艳强, 包维楷
- JIN Yanqiang, BAO Weikai
- 四川柏木人工林林下植被生物量与林分结构的关系
- Relationships of the understory biomass with stand structure of the Sichuan cypress plantation forests across Sichuan Basin, China
- 生态学报, 2014, 34(20): 5849-5859
- Acta Ecologica Sinica, 2014, 34(20): 5849-5859
- http://dx.doi.org/10.5846/stxb201301210127
-
文章历史
- 收稿日期:2013-1-21
- 网络收稿日期:2014-3-11






2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
林下植被一般指高度小于3 m的林下植物层片,主要由灌木、乔木幼苗以及草本组成,是森林的必要组分,在保育乡土动植物区系和生物多样性、为其它动物提供生境和食物来源、养分循环、促进林地生产力的长期持续性及促进森林更新和演替等方面发挥着重要作用[1, 2]。因此,林下植被一直是森林持续经营、生物多样性保护和森林功能维持的关键[2, 3, 4]。然而几十年来对森林生物量与生产力的研究主要聚焦于林木,对林下植被生物量与生产力的研究缺乏重视,与林分结构的关系认知严重不足[5]。林下植被总生物量虽然不高,但其细根生物量比例较高[6],作为森林的一个活跃的养分库,林下植被总生物量在森林养分循环以及林地生产力维持中起到关键作用。林下植被发育与森林冠层结构紧密相关[7],揭示林下植被生物量与森林结构的关系是认识森林结构与功能动态变化的基础。理论上,林下植被的生物量不仅受自身结构(盖度、基径、高度、体积(盖度与高的乘积)等)直接影响[8, 9],更受到乔木冠层的复杂制约。虽有一些研究揭示了灌丛或草地的生物量与群落结构性状的关系[10, 11],但林下植被的生物量与其结构性状的关系缺乏必要研究;而林下植被生物量与冠层结构的关系虽有少量研究,但结论并不统一。一些研究发现林下植被生物量受林分冠层影响严重[12, 13, 14],而另一些研究表明林下植被生物量的变化与林分密度没有明显相关关系[15, 16]。因此,进一步明确林下植被生物量与林分结构的关系是必要的。对柏木林的研究主要集中在柏木林的生态效应方面,其中柏木结构与林分生物量有不少研究,但大多数是调查林分生物量现状,涉及与林分结构因子(密度、胸高直径与高度)的关系探讨[17, 18, 19, 20]。已有研究发现,柏木人工林林下植被生物量占到群落总生物量的3.7%—23.6%[20, 21],不容忽视。而针对柏木人工林林下植被生物量与林分结构的关系至今缺乏研究。另外,估测林下植被生物量是森林功能及森林碳清算计量不可缺少的内容。传统而广泛采用的林下植被生物量测定方法是直接全收获法,很少有研究通过模型来估算林下植被生物量[17],在已有的少数林下植被生物量估测研究中,主要聚焦单优势灌木种群生物量估算模型(Species-specific models)的研究[22, 23, 24]。黎燕琼等[25]研究过柏木人工林林下优势灌木黄荆(Vitex negundo)的基径、均高等结构参数与其生物量的关系,采用基径和均高发展了黄荆生物量的单种估算模型,获得了较好的估算效果(R2=0.906)。然而,柏木人工林林下植被物种组成丰富,向成华等[26]的调查发现柏木人工纯林灌木层物种数为15,草本层28种。而且柏木人工林林下植被优势种地位也并不总是突出[20, 27],尤其是在中龄林阶段或密林状态。显然使用优势种单种模型不适用于估算区域林下植被生物量。针对每个种建立异速生长方程的工作量较大,且难以实现。因此,对区域性广泛分布的柏木林下植被生物量估算迫切需要发展基于林下植物总体易测结构参数的生物量估测混合模型(Mixed-species models),以提高估算效率。因此,本文选取四川盆地柏木人工林展开调查研究,主要目的是揭示林分结构参数与林下植被生物量关系,以及使用林分结构参数构建区域林下植被生物量的估测模型。研究结果可为四川柏木生物量估测以及碳计量体系建设提供直接的模型和方法依据。
1 研究区概况柏木(Cupressus funebris)是20世纪60年代以来四川盆地集体林区荒山绿化造林的主要树种,目前柏木人工林成为川中丘陵区的优势森林类型之一[28]。根据四川森林监测中心的数据,柏木林面积从1979年的49.4万hm2已增加到2007年的145.2万hm2,在长江上游防护林工程体系建设中发展迅速,以四川、湖北西部、贵州栽培最多。研究区域覆盖广元、巴中、绵阳、南充、遂宁等地市的12个县区,东经103°30′—109°30′,北纬28°40′—32°40′之间,海拔350—700 m,地貌以丘陵为主,是20世纪80年代长江上游防护林建设中柏木造林的重点区域。该区属亚热带湿润季风气候,四季分明,年降水量一般为1000—1200 mm,年均温16—18 ℃,全年日照时数1000—1400 h。土壤以紫色土为主,土层浅薄,肥力不高,因此发育的柏木林生产力(生物量)不高[20],群落结构比较简单,林下植被成丛明显。林下主要灌木种类主要有黄荆、马桑(Coriaria nepalensis)、铁仔(Myrsine africana)、火棘(Pyracantha fortuneana)等,而草本主要有白茅(Imperata cylindrica)、莎草(Cyrerus microiria)、荩草(Arthraxon hispidus)等[20, 27]。
2 研究方法 2.1 样方调查2011年在研究区针对柏木人工林设置了14个面积为1000 m2 (20 m×50 m)的典型样地,其中一块样地受地形限制,样地面积调整为800 m2 (20 m×40 m),然后把样地划分成100 m2 (10 m×10 m)的乔木样方,分别记录每一个样方的郁闭度Ct,然后进行每木检尺,测定大于3 m的乔木层每株的树高、胸径、冠幅。在样地中机械布点设置 3 个 2 m×2 m的灌木样方,布设于乔木亚样方。灌木样方调查了灌木物种组成,测定了总盖度,然后分种测定每一灌木群体的平均基径、平均高度后,将样方内所有灌木全部收获后,分枝茎、叶、根三部分就地称鲜重,然后各部分分别取约 300 g 混合样带回实验室测定含水率,以便将鲜生物量换算为干重。在3个灌木样方的右上角,分别设置 1 个 1 m × 1 m 的草本样方,估测总盖度后记录种类组成,然后分种测定各草群平均高度;然后将样方内所有草本植物全部收获,分地上部分和地下部分称鲜重,然后取约 300 g 混合样带回实验室测定含水率。把灌木和草本样品都在70 ℃烘3天后测定含水率,以含水率将灌木和草本各样方生物量鲜重按成分(叶、枝茎、根,地上、地下)换算成干重生物量(t/hm2)。野外调查了14块样地,样地基本概况见表 1,获取乔木样方的乔木层结构参数以及相应的灌草样方数据各42个,由于其中2个样方无灌草而没纳入模型构建,因此本文用40个样方数据进行分析。
| 样地Site | 经度Latitude | 纬度Longitude | 海拔/m Altitude | 坡向Aspect | 坡度/ (°) Slope | 坡位Slope position | 样地面积/m2 Stand area | 郁闭度Corerage | 林分密度 Density/(株/hm2) |
| P1 | 105°17.69′ | 31°22.16′ | 525 | ES10° | 27 | 中 | 1000 | 0.6 | 3470 |
| P2 | 105°25.21′ | 31°13.48′ | 540 | NW20° | 29 | 中 | 1000 | 0.7 | 4430 |
| P3 | 105°10.10′ | 31°04.85′ | 436 | ES40° | 29 | 中 | 1000 | 0.6 | 3110 |
| P4 | 105°22.57′ | 30°47.51′ | 396 | SW20° | 38 | 上 | 1000 | 0.6 | 2530 |
| P5 | 105°37.64′ | 30°48.45′ | 345 | NE15° | 41 | 下 | 1000 | 0.7 | 990 |
| P6 | 105°22.64′ | 30°38.89′ | 359 | NW5° | 52 | 中 | 1000 | 0.65 | 1370 |
| P7 | 105°40.43′ | 31°30.74′ | 600 | SW29° | 44 | 中 | 1000 | 0.6 | 1770 |
| P8 | 107°40.16′ | 32°03.76′ | 790 | SE20° | 40 | 下 | 1000 | 0.4 | 960 |
| P9 | 107°04.37′ | 31°55.76′ | 660 | SW9° | 39 | 中 | 800 | 0.9 | 3050 |
| P10 | 105°40.49′ | 31°35.05′ | 440 | NW39° | 31 | 上 | 1000 | 0.85 | 2460 |
| P11 | 106°46.05′ | 31°30.08′ | 595 | S | 49 | 中 | 1000 | 0.7 | 1860 |
| P12 | 106°03.21′ | 31°34.90′ | 488 | NW70° | 54 | 上 | 1000 | 0.65 | 3490 |
| P13 | 107°01.20′ | 31°25.57′ | 500 | SW33° | 43 | 中 | 1000 | 0.5 | 1650 |
| P14 | 106°53.87′ | 31°38.63′ | 540 | SE47° | 51 | 中 | 1000 | 0.78 | 690 |
根据野外调查获得各样方中林下灌木总盖度Cs(%)、平均高度Hs(cm),计算灌木群体平均基径Ds (cm)及体积 Vs (Vs=Cs×Hs)[29];根据草本群体的均高Hh(cm)与盖度Ch(%),然后计算出草群体积Vh (Vh =Ch×Hh);根据含水率和鲜重,计算获得每个小样方中的各类群以及总体各部分的干重,然后换算林下植被各部分生物量(t/hm2):林下植被总生物量 Wu、林下灌木生物量Ws及林下草本生物量Wh。根据乔木样方实测值胸径,计算平均胸径Da (cm) 、平均高度Ht (m)以及林分密度Du(株/hm2)。最后按圆柱计算出胸高断面积Bt(m2)。各参数基本范围见表 2。使用Shapiro-Wilk指数对林下植被生物量参数(Wu、Ws、Wh)以及林分结构参数(Hs、Cs、Vs、Hh、Ch、Vh、Ct、Bt、Da、Du、Ht)进行正态性检验,对不满足正态性检验的变量进行自然对数转换。然后使用Pearson相关分析去揭示林下植被生物量与林分结构的关系,根据相关系数判断紧密程度。
| Hs/cm | Ds/cm | Cs/% | Hh/cm | Ch/% | Cu/% | Ct | Du /(株/hm2) | Bt /m2 | Da /cm | Ht/m | |
| Hs: 灌木均高average shrub height; Ds: 灌木平均基径average diameter for shrub; Cs: 灌木盖度shrub cover; Hh: 草本均高average herbaceous height; Ch: 草本盖度herbaceous cover; Cu: 林下植被总盖度understory cover; Ct: 郁闭度canopy; Du: 林分密度stand density; Bt: 总胸高断面积total basal area; Da: 平均胸径average diameter at basal height; Ht: 乔木均高average height for overstory | |||||||||||
| 样方数N | 40 | 40 | 40 | 40 | 40 | 40 | 40 | 40 | 40 | 40 | 40 |
| 均值Mean | 82 | 0.91 | 29.4 | 20.8 | 52 | 84.6 | 0.6 | 1995.0 | 0.136 | 9.1 | 8 |
| 标准误SE | 5.7 | 0.14 | 3.5 | 1 | 4.4 | 1.9 | 0 | 208.3 | 0.018 | 0.6 | 0.4 |
| 最小值Minimum | 31.8 | 0.14 | 0.3 | 8 | 9 | 57 | 0.2 | 300.0 | 0.003 | 3.2 | 3.7 |
| 最大值Maximum | 176 | 4.97 | 83 | 34 | 100 | 99 | 0.9 | 5400.0 | 0.677 | 21.4 | 14.7 |
基于前面的相关分析,挑选与林下植被生物量(Wu、Ws、Wh)关系紧密的结构参数作为预测变量,分别对Wu、Ws、Wh进行多种模型的拟合。根据已有研究中对林下植被生物量与形态结构特性关系结果[30],选用的模型方程形式有
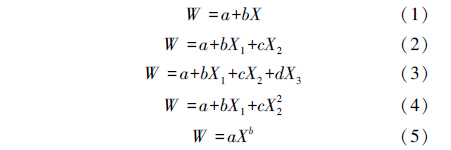
式中,X、X1、X2、X3即为模型预测变量,a、b、c为常系数。在拟合中,如模型有2个参数时就选取任意2个预测变量,如3个参数时即随机选择3个预测变量进行拟合,使用F检验对其进行模型显著性检验。采用调整的决定系数 (R2a)和模型估计误差(SEE)两个指标来评价显著性模型的优劣,R2a较大,且SEE较小的模型作为最佳估算模型[31]。为了增强模型在空间的延展性,选取与灌草样方相对应亚样方的林分结构性状因子包括Du、郁闭度、总胸高断面积作为变量,采用逐步回归方法对林下植被生物量进行模型拟合。使用AIC值(Akaike Information Criterion)作为模型选取的标准,取AIC值最低的模型作为最佳拟合模型[32]。AIC值计算通过方程:
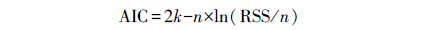
式中,k是参数数量,RSS为剩余平方和,n为样本数40。数据分析和模型的拟合检验均使用R.2.15.0软件[33]完成。
3 结果与分析 3.1 林下植被总生物量与林分结构的关系柏木Wu与林分各层片结构参数的关系差异较大(图 1)。Wu与灌木Ds (R=0.493,P<0.001),Cs(R=0.570,P<0.001)和 Vs(R=0.725,P<0.001)的关系紧密(图 1),与Hs(R=0.799,P<0.001)的关系最为紧密;而Wu与Vh(R=0.333,P=0.036)的关系紧密,与草本其它的结构参数Ch、Hh和Vh的关系,以及与乔木层的Ht、Ct、Dt、Bt、Da这5个结构参数间的相关关系都不显著(P > 0.05)。

|
| 图 1 四川柏木林下植被生物量 (Wu) 与林分结构的关系 Fig. 1 Pearson′s correlations between biomass with structural parameters of overstory and understory in the Cupressus funebris plantation forests from Sichuan Basin Hs,灌木均高average shrub height; Ds,灌木平均基径average diameter for shrub; Cs,灌木盖度shrub cover; Vs,灌木体积(Vs =Hs×Cs)shrub volume (Vs =Hs×Cs); Hh,草本均高average herbaceous height; Ch,草本盖度herbaceous cover; Vh,草本体积(Vh =Hh×Ch)herbaceous volume (Vh =Hh×Ch); Ct,郁闭度canopy; Du,林分密度stand density; Bt,胸高断面积basal area; Da,平均胸径average diameter at basal height; Ht,乔木均高average height for overstory; R,相关系数correlation coefficient;ns, P >0.05; *,P<0.05; **,P<0.01; ***,P<0.001 |
柏木林下灌木Ws与自身结构参数 Hs(R=0.725,P<0.001)、Ds(R=0.612,P<0.001)、Cs(R=0.593,P<0.001)和Vs(R=0.637,P<0.001)的关系也较为紧密(图 2);Ws与草本的Ch、Hh和Vh的关系都不显著(P > 0.05);然而Ws与林分结构参数中的Bt有显著负相关关系(R=-0.516,P<0.001),而与Ht、Ct、Dt和Da的关系都不显著(P > 0.05)。

|
| 图 2 四川柏木林林下灌木生物量 (Ws) 与上层乔木和自身结构参数的相关关系 Fig. 2 Correlations between the shrub biomass and structural parameters of overstory and understory by Pearson′s correlation under the Cupressus funebris plantation forests in Sichuan Basin |
林下草本生物量Wh与自身的 Ch的关系显著(R=0.377,P=0.016;图 3),而Vh的关系更紧密(R=0.464,P=0.003),但与Hh之间没有显著关系(P > 0.05)。而Wh与灌木结构参数Hs (R=0.444,P=0.004)和Vs (R=0.367,P=0.02)关系都较为紧密,但与灌木的Ds和Cs没有显著关系(P > 0.05)。Wh与乔木层结构参数(Ht、Ct、Bt、Dt和Da)间的关系都不显著(P > 0.05)。

|
| 图 3 四川柏木林林下草本生物量 (Wh) 与上层乔木和自身结构的相关关系 Fig. 3 Correlations between the herbaceous biomass and structural parameters of overstory and understory by Pearson′s correlation under the Cupressus funebris plantation forests in Sichuan Basin |
如前面的Pearson相关分析结果所示(图 1—3),林下植被及其各组分的生物量与其自身的结构参数关系最为紧密,而与林分特征因子间的关系大多数并不显著。据此,选取林下的灌木与草本的7个结构参数(Ds、Hs、Cs、Vs、Hh、Ch、Vh)作为预测变量,按照5种方程形式,分别拟合林下植被及其各组分生物量的估算模型。模型拟合发现,选用不同的预测变量及模型对柏木林下灌木Ws及其器官(叶Wsl、枝茎Wss、地上Wsa和地下Wsb)的估测效果差异大(表 3)。林下灌木叶、枝茎、地上、地下和总的生物量的最佳估算模型都是以Hs和Cs组合的参数Vs的幂函数形式,分别为Wsl=0.0001×V1.0268s(R2a=0.760,P<0.001)、Wss=0.0001×V1.1593s(R2a=0.754,P<0.001)、Wsa=0.0001×V1.1171s(R2a=0.784,P<0.001)、Wsb=0.0003×V0.9578s(R2a=0.651,P<0.001)和Ws=0.0005×V1.0411s(R2a=0.762,P<0.001)
| 层次Layer | 部位Components | 模型Model | 系数Coefficients | R2a | SEE | F | ||
| a | b | c | ||||||
| R2a,调整的决定系数adjusted R-square; Wsl,灌木叶生物量leaf biomass for shrubs; Wss,灌木枝茎生物量stem and branch biomass for shrubs; Wsa,灌木地上生物量aboveground biomass for shrubs; Wsb,灌木地下生物量belowground biomass for shrubs; Ws,灌木总生物量total shrubs biomass; Hs,灌木均高average shrub height; Cs,灌木盖度shrub cover; Vs,灌木体积(Vs =Hs×Cs)shrub volume (Vs =Hs×Cs); Wha,林下草本地上生物量aboveground biomass for herbs; Wh,林下草本总生物量total herbs biomass; Hh,草本均高average herb height; Ch,草本盖度herb cover; Wua,林下植被地上生物量aboveground biomass for understoy; Wub,林下植被地下生物量belowground biomass for understory; Wu,林下植被总生物量total understory biomass; SEE:模型的估计误差standard error of the estimate; **,P<0.01;***,P<0.001 | ||||||||
| 灌木Shrub | 叶 | Wsl=a×Vbs | 0.0001 | 1.0268 | 0.760 | 0.886 | 124.19*** | |
| 枝茎 | Wss=a×Vbs | 0.0001 | 1.1593 | 0.754 | 1.016 | 120.60*** | ||
| 地上 | Wsa=a×Vbs | 0.0001 | 1.1171 | 0.784 | 0.900 | 142.59*** | ||
| 地下 | Wsb=a×Vbs | 0.0003 | 0.9578 | 0.651 | 1.072 | 73.87*** | ||
| 总体 | Ws=a×Vbs | 0.0005 | 1.0411 | 0.762 | 0.892 | 125.96*** | ||
| 草本Herb | 地上 | lnWha=a+bHh+cCh | -1.9656 | 0.0337 | 0.0149 | 0.410 | 0.534 | 14.55*** |
| 总体 | lnWh=a+bHh+cCh | -0.8429 | 0.0322 | 0.0094 | 0.201 | 0.599 | 5.90** | |
| 林下植被 | 地上 | lnWua=a+bHs+cCs | -1.2839 | 0.0151 | 0.0151 | 0.700 | 0.484 | 46.52*** |
| Understory | 地下 | lnWub=a+bHs+cCs | -1.2714 | 0.0166 | 0.0069 | 0.538 | 0.614 | 23.70*** |
| 总体 | lnWu=a+bHs+cCs | -0.5358 | 0.0158 | 0.0111 | 0.695 | 0.463 | 45.51*** | |
在草本生物量的拟合中发现,使用草本的均高(Hh)、盖度(Ch)和体积(Vh)进行拟合,虽然通过了显著性检验,但模型精度都较低 (R2a<0.410,P<0.01,表 3)。林下草本总生物量的最佳估算模型为多元线性方程lnWh=a+bHh+cCh,但估算精度较低 (R2a=0.201,P=0.006);最佳估算林下草本地上生物量的模型同为多元线性方程lnWha=a+bHh+cCh,模型精度也不高 (R2a=0.410,P<0.001)。特别指出的是,在构建林下草本地下生物量估算模型中发现,草本的均高(Hh)、盖度(Ch)和体积(Vh)等指标在5种不同模型构建中,均未能通过显著性检验(P > 0.05)。进一步在拟合建立林下植被总生物量估算模型中发现,使用灌木Hs、Cs和Vs去估算Ws获得了较好的估算精度(表 3)。估算林下植被地上(R2a=0.700,P<0.001)、地下(R2a=0.538,P<0.001)和总体生物量(R2a=0.695,P<0.001)的最佳模型形式均为多元线性方程lnW =a+bHs+cCs。而林下草本体积Vh在模型拟合中贡献较小(R2a=0.088,P=0.04),草本Hh和Ch在模型拟合中无法通过显著性检验(P > 0.05)。逐步回归分析发现,采用林分密度(Du)、灌木均高(Hs)和灌木盖度(Cs)预测林下植被生物量获得了较高的估算精度(表 4)。与前述多元模型相比,林分密度Du的加入使得林下植被地上(R2a=0.732,P<0.001)、地下(R2a=0.558,P<0.001)及总生物量(R2a=0.721,P<0.001)估算模型的精度得到有效提高(R2a增大)。
| 部位Components | 模型 Model | 系数Coefficients | R2a | SEE | F | AIC | |||
| a | b | c | d | ||||||
| R2a: 调整的决定系数;Wua: 林下植被地上生物量;Wub: 林下植被地下生物量;Wu: 林下植被总生物量Hs: 灌木均高;Du: 林分密度;Cs: 灌木盖度;SEE:模型的估计误差;AIC:赤池信息量准则;*** P<0.001 | |||||||||
| 地上Wua | lnWua =a+bDu+cHs+dCs | -0.9894 | -0.0001 | 0.0155 | 0.0131 | 0.732 | 0.457 | 36.59*** | 60.85 |
| 地下Wub | lnWub =a+bDu+cHs | -0.9153 | -0.0001 | 0.0182 | 0.558 | 0.601 | 25.57*** | 84.13 | |
| 总体Wu | lnWu=a+bDu+cHs+dCs | -0.2813 | -0.0001 | 0.0161 | 0.0094 | 0.721 | 0.443 | 34.53*** | 59.22 |
林下植被盖度、高度、体积等结构参数与林下生物量相关关系突出[34],因而常作为林下植被生物量模型构建的基本预测变量使用[9, 35]。研究结果表明,林下植被生物量跟灌木层片结构参数相关更明显(图 1),但与草本层片结构参数并不相关。进一步发现,林下植被生物量与灌木平均高度的关系更为紧密,而不是灌木盖度与密度。这表明森林中林下灌木群体平均高度及体积能够更好反映林下灌木对林下异质性资源环境空间的生态适应能力,因为在林下更高的灌木生长意味着可更有效的获取限制性光资源[2]。但是,林下灌木生物量与其结构参数的关联程度与纯粹的灌丛是明显不同的[10, 36, 37, 38],这应该归因于两类环境中限制性资源类型、程度以及在不同的资源生态位下灌木形态性状的权衡关系可塑性的明显差异。一般缺乏森林林冠抑制作用下的灌木生长,通过分枝和萌蘖向水平空间拓展是明显的,因此其冠幅或冠直径及分枝程度可更好指示资源利用能力与生物量累积[36, 39, 40]。林下植被生物量与林分郁闭度、密度、胸高断面积等参数关系紧密,因为这些参数综合反映着林分拥挤程度,控制着林内光资源格局,这对林下植被的生长、生物量积累造成直接影响[14, 15, 41]。但是,单独使用各因子进行相关分析时,本研究没有发现存在明确的相关关系。这可能受到林分年龄的严重影响[16, 42],因为林龄反应出森林发育程度与环境的稳定性,能够一定程度上指示林分透光状况,处于不同年龄阶段的群落树种组成、冠型等都会改变林下的光环境,从而影响林下植被的盖度与生物量[2, 7]。干扰的影响也是不能排除的,林下植被对干扰的影响最为敏感[43]。研究区内的柏木林大多是荒山荒坡造林形成的,位于村庄周围,被农田包围,已成为农户承包山林,深受林下放牧、灌草收割积肥、及偷伐自用材等活动干扰,虽然调查时尽量排除这种有明显影响痕迹的林分,但这种干扰对林下植被生物量的影响不容忽视。Ares等[13]的研究就发现,疏伐可显著促进林下植被变化,改变林分结构与下木层的生产能力。因此,深入揭示林分结构调整及林下植被管理对林下生物量与林分结构特征因子关系的影响还需进一步研究。
4.2 柏木林下植被生物量模型本研究发现,柏木人工林林下灌木群体生物量优化混合模型是以体积Vs为变量的幂函数方程W = a×Vbs,比单一采用平均高、基径或盖度的精度更高,表明在柏木这样塔型的单优势种组成的林分环境中,灌木群体的性状关系复杂,性状权衡关系存在较大可塑性,采用综合指标能反映其资源利用生态位的复合性状,因而可获得更好估测效果。本文的研究结果与黎燕琼等[25]针对柏木人工林林下优势灌木黄荆建立的单种模型是不同的,柏木人工林林下优势灌木黄荆的最佳单种模型为以基径为参数建立的二次或三次曲线方程,其单种模型的估算精度(R2=0.906)稍高于本研究的混合模型(R2a=0.762),显然与本研究的大区域的空间异质性取样相关,因为植物性状权衡关系深受资源供给程度影响,而表现出对资源利用的可塑性差异。在混合模型构建中,群体基径仅对灌木枝生物量估算效果较好,相反,平均高度、盖度、体积在混合模型构建中获得了较为理想的估算效果[8, 37],这主要是由于灌木多分枝现象,致使基径并不充分指示灌木形态[36, 44]。Sah等[44]也指出,对于多分枝的灌木,把每个分枝当作一个个体对待,能够提高模型的精度。
在柏木人工林林下草本层混合模型构建中,使用高度、盖度、体积等因子及其组合去建立估算模型都未能获得较好的估算精度(R2a<0.410,P<0.01),尤其是草本地下生物量估算模型未能通过显著性检验(P > 0.05),这明显跟前述的林下灌木不同。这可能与冠层覆盖下林下草本群体的形态变异紧密相关,因为草本的形态直接影响着草本的盖度、高度、体积等属性变异,对草本层各因子的测算带来不确定性[34, 45]。MacDonald等[35]充分考虑草本植物形态,把草本植物细分为高草、矮草、禾草和非禾草几个功能群组,分别构建了草本植物不同功能群生物量的混合模型,就获得了较高的估算精度(R2> 0.81),为推测提供了佐证。另外,由于草本植物的盖度和生物量受到生长季的影响较为严重,不同生长季和不同年份之间其估算模型都不同[15],致使草本植物生物量模型的构建带来困难[46]。
针对林下植被总生物量的优化估算模型为多元线性模型lnWu=a+bHs+cCs (表 3),而在逐步回归中加入林分密度后,林下植被各部分生物量(地上、地下部分)的模型精度都有所提高,表明冠层结构显著左右着林下环境资源时空异质性,制约着林下生物量和生产量。各种模型中,使用多个因子参与拟合比单个因子作为参数的模型有更好的估算效果[22, 38, 40]。但是,如单独使用林分密度、郁闭度、胸高断面积等林分立地特征因子进行模型拟合时,模型未能通过显著性检验(P > 0.05;图 1—图 3)。这说明在林下植被生物量模型构建中,林下植被自身结构参数比林分特征因子更适用于其生物量模型构建,而林分因子是通过左右林下资源环境制约林下植被结构性状而间接去影响林下植被发育与生物量积累的[7]。不同模型之间其适用范围有所不同。
已有研究发现,单种模型比起混合模型有更高的估算精度[39, 47]。然而,曾慧卿等[38]在红壤丘陵区对比了不同林下物种生物量的单种模型和混合模型,发现白栎(Rhododendron simsii)、山莓(Rubus corchorifolius)和乌饭树(Vaccinium bracteatum)等树种的混合模型优于单种模型。在区域生物量估算中,无法对每一物种进行单种模型构建,使用混合模型比单种模型更高效、简便,在精度允许的情况下可作为区域林下植被生物量估算的首选方法。
总体来看,本研究揭示了四川柏木人工林林下生物量与林分结构的关系,发展了有效的柏木林下植被生物量与灌木生物量有效估测模型,为四川柏木林林下生物量估算与碳计量体系提供了依据。但依据调查获得的40个林下植被生物量数据建立估测模型,数据偏少,增加更多的数据进行更全面的数据拟合是必要的。此外,由于林下植被生物量受到多种因素影响,在应用各个混合模型前,进一步开展模型验证也是必要的。
致谢: 参加野外调查的还有杜忠、方志强、刘鑫、王喆、赵庆霞、冯德枫以及四川农业大学暑期实习学生;调查过程中承蒙四川省林勘院张文、赖长鸿、张炎周等及巴中、南充、广元、绵阳等相关市、县林业局的支持和帮助,特此致谢。
| [1] | Nilsson M C, Wardle D A. Understory vegetation as a forest ecosystem driver: evidence from the northern Swedish boreal forest. Frontiers in Ecology and the Environment, 2005, 3(8): 421-428. |
| [2] | Wagner S, Fischer H, Huth F. Canopy effects on vegetation caused by harvesting and regeneration treatments. European Journal of Forest Research, 2011, 130(1): 17-40. |
| [3] | Brockerhoff E G, Jactel H, Parrotta J A, Quine C P, Sayer J. Plantation forests and biodiversity: oxymoron or opportunity? Biodiversity and Conservation, 2008, 17(5): 925-951. |
| [4] | Bremer L, Farley K. Does plantation forestry restore biodiversity or create green deserts? A synthesis of the effects of land-use transitions on plant species richness. Biodiversity and Conservation, 2010, 19(14): 3893-3915. |
| [5] | Schmiedinger A, Kreyling J, Steinbauer M J, Macdonald S E, Jentsch A, Beierkuhnlein C. A continental comparison indicates long-term effects of forest management on understory diversity in coniferous forests. Canadian Journal of Forest Research, 2012, 42(7): 1239-1252. |
| [6] | Gonzalez M, Augusto L, Gallet-Budynek A, Xue J M, Yauschew-Raguenes N, Guyon D, Trichet P, Delerue F, Niollet S, Andreasson F, Achat D L, Bakker M R. Contribution of understory species to total ecosystem aboveground and belowground biomass in temperate Pinus pinaster Ait. forests. Forest Ecology and Management, 2013, 289: 38-47. |
| [7] | Strong W L. Tree canopy effects on understory species abundance in high-latitude Populus tremuloides stands, Yukon, Canada. Community Ecology, 2011, 12(1): 89-98. |
| [8] | Brown J K. Estimating shrub biomass from basal stem diameters. Canadian Journal of Forest Research, 1976, 6(2): 153-158. |
| [9] | Röttgermann M, Steinlein T, Beyschlag W, Dietz H. Linear relationships between aboveground biomass and plant cover in low open herbaceous vegetation. Journal of Vegetation Science, 2000, 11(1): 145-148. |
| [10] | Liu X L, Hao X D, Yang D S, Liu S R, Su Y M, Cai X H, He F, Ma Q Y. Aboveground biomass and its models of Quercus aquifolioides thicket community in Balangshan Mountain in Wolong Natural Reserve. Chinese Journal of Ecology, 2006, 25(5): 487-491. |
| [11] | Li G T, Qing F C, Jia S Y, Dong J L, Jirgele, Wang M G. Study on biomass estimation models of xerophyte shrub. Journal of Inner Mongolia Forestry College, 1998, 20(2): 24-31. |
| [12] | Pieper R D. Overstory-understory relations in pinyon-juniper woodlands in New Mexico. Journal of Range Management, 1990, 43(5): 413-415. |
| [13] | Ares A, Neill A R, Puettmann K J. Understory abundance, species diversity and functional attribute response to thinning in coniferous stands. Forest Ecology and Management, 2010, 260(7): 1104-1113. |
| [14] | Mitchell J E, Popovich S J. Effectiveness of basal area for estimating canopy cover of ponderosa pine. Forest Ecology and Management, 1997, 95(1): 45-51. |
| [15] | González-Hernández M P, Silva-Pando F J, Jiménez M C. Production patterns of understory layers in several Galician (NW Spain) woodlands: Seasonality, net productivity and renewal rates. Forest Ecology and Management, 1998, 109(1/3): 251-259. |
| [16] | Zhang W P, Huang J C, Li X B. Analysis of influencing factors of undergrowth biomass in Cunninghamia lanceolata plantations. Journal of Fujian Forestry Science and Technology, 2007, 34(3): 97-99, 149-149. |
| [17] | Zheng S W, Tang M, Zou J H, Mu C L. Summary of research on shrub biomass in China. Journal of Chengdu University: Natural Sciences, 2007, 26(3): 189-192. |
| [18] | Wu P F, Zhu B, Liu S R, Wang X G. Carbon storage and its allocation in mixed alder-cypress plantations at different age stages. Chinese Journal of Applied Ecology, 2008, 19(7): 1419-1424. |
| [19] | Zhu Y E, Yao D M. A study on the development and biomass of undergrowth vegetations in Cupressus funebris plantations in Yichang Suburbs. Forest Resources Management, 2007, (2): 52-56. |
| [20] | Wang J. Community characteristics and its biomass in alder and cypress mixed young forest. Journal of Sichuan Forestry Science and Technology, 1993, 14(1): 66-69. |
| [21] | Liu X L, Yan W X, Xiang C H, Jiang J M. Biomass and biomass models of secondary subtropical vegetation in Tuojiang river valley. Acta Phytoecologica Sinica, 1997, 21(5): 441-454. |
| [22] | Zeng Z Y, Liu Q J, Zeng H Q. Study of biomass models of some kinds of shrubs in Qianyanzhou, Jiangxi province. Journal of Fujian Forestry Science and Technology, 2005, 32(4): 68-72. |
| [23] | Dickinson Y L, Zenner E K. Allometric equations for the aboveground biomass of selected common eastern hardwood understory species. Northern Journal of Applied Forestry, 2010, 27(4): 160-165. |
| [24] | Zeng H Q, Liu Q J, Feng Z W, Ma Z Q. Biomass equations for four shrub species in subtropical China. Journal of Forest Research, 2009, 15(2): 83-90. |
| [25] | Li Y Q, Zheng S W, Gong G T, Chen J H, Tang B, Zhu Z F, Wu X X, Mu C L. Biomass and its allocation of undergrowth Vitex negundo L. in different age classes of mixed cypress forest. Acta Ecologica Sinica, 2010, 30(11): 2809-2818. |
| [26] | Xiang C H, Luo Z S, Chen J H, He F, Luo X H. Research on characteristics of community structure of the main forests in hilly regions of the Sichuan basin. Journal of Sichuan Forestry Science and Technology, 2005, 26(5): 25-29. |
| [27] | Zheng S W, Niu M, Zhang Q, Li Y Q, Mu C L, Gong G T, Chen J H, Zhu Z F, Wu X X. A study of the vegetation community structure and biodiversity of different Cupressus funebris types in hilly areas of central Sichuan province. Journal of Sichuan Forestry Science and Technology, 2011, 32(5): 20-28. |
| [28] | Li R W. Researches on Construction and Management of Protection Forest System in Upper Reach of Yangtze River. Chengdu: Sichuan Science and Technology Press, 2004. |
| [29] | Porté A J, Samalens J C, Dulhoste R, Teissier Du Cros R, Bosc A, Meredieu C. Using cover measurements to estimate aboveground understorey biomass in Maritime pine stands. Annals of Forest Science, 2009, 66(3): 307-307. |
| [30] | Lu Z L, Gong X S. Progress on the research of shrub biomass estimation. Forest Inventory and Planning, 2009, 34(4): 37-40, 45-45. |
| [31] | Zar J H. Biostatistical Analysis. 4th ed. New Jersey: Prentice Hall, 1999: 419-424. |
| [32] | Hurvich C M, Tsai C L. Regression and time series model selection in small samples. Biometrika, 1989, 76(2): 297-307. |
| [33] | R Development Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria, 2012. http://www.R-project.org/. |
| [34] | Wayne E R, Van Auken O W. Herbaceous cover and biomass of carex planostachys in Juniperus ashei woodlands of central texas. The Southwestern Naturalist, 2010, 55(3): 434-441. |
| [35] | MacDonald R L, Burke J M, Chen H Y H, Prepas E E. Relationship between aboveground biomass and percent cover of ground vegetation in Canadian boreal plain riparian forests. Forest Science, 2012, 58(1): 47-53. |
| [36] | Sun S C, Qian N B. Path analysis of morphological parameters of Convolvulus tragacuthoiedes population and individual biomass modeling of subshrubs. Chinese Journal of Applied Ecology, 1999, 10(02): 155-158. |
| [37] | Zeng H Q, Liu Q J, Ma Z Q, Zeng Z Y. The regression model of Loropetalum chinense biomass based on canopy diameter and plant height. Journal of Nanjing Forestry University: Natural Science, 2006, 30(4): 101-104. |
| [38] | Zeng H Q, Liu Q J, Feng Z W, Ma Z Q, Hu L L. Estimation models of understory shrub biomass and their applications in red soil hilly region. Chinese Journal of Applied Ecology, 2007, 18(10): 2185-2190. |
| [39] | Návar J, Nájera J, Jurado E. Biomass estimation equations in the Tamaulipan thornscrub of north-eastern Mexico. Journal of Arid Environments, 2002, 52(2): 167-179. |
| [40] | Wang L, Zhang H, Hasi, Zheng Q H, Liu L Y. A study on the estimating method of shrub upper biomass based on the crown diameter and plant height. Journal of Beijing Normal University: Natural Science, 2004, 40(05): 700-704. |
| [41] | Peek J M, Korol J J, Gay D, Hershey T. Overstory-understory biomass changes over a 35-year period in southcentral Oregon. Forest Ecology and Management, 2001, 150(3): 267-277. |
| [42] | Pan P, Mu C C, Sun Z H. Biomass of shrub and herb layers in Larix olgensis plantations. Journal of Northeast Forestry University, 2007, 35(04): 1-2, 6-6. |
| [43] | Hart S A, Chen H Y H. Understory vegetation dynamics of north american boreal forests. Critical Reviews in Plant Sciences, 2006, 25(4): 381-397. |
| [44] | Sah J P, Ross M S, Koptur S, Snyder J R. Estimating aboveground biomass of broadleaved woody plants in the understory of Florida Keys pine forests. Forest Ecology and Management, 2004, 203(1/3): 319-329. |
| [45] | Joshi M, Rawat Y S. Net primary productivity and species diversity of herbaceous vegetation in banj-oak (Quercus leucotrichophora A. Camus) forest in Kumaun Himalaya, India. Journal of Mountain Science, 2011, 8(6): 787-793. |
| [46] | Maybodi N B, Arzani H, Zare M T. Relationship between cover and yield of some range species in steppic region of Yazd province. BIABAN (Desert Journal), 2006, 11(2): 57-67. |
| [47] | Návar J, Méndez E, Nájera A, Graciano J, Dale V, Parresol B. Biomass equations for shrub species of Tamaulipan thornscrub of North-eastern Mexico. Journal of Arid Environments, 2004, 59(4): 657-674. |
| [10] | 刘兴良, 郝晓东, 杨冬生, 刘世荣, 宿以明, 蔡小虎, 何飞, 马钦彦. 卧龙巴郎山川滇高山栎灌丛地上生物量及其模型. 生态学杂志, 2006, 25(5): 487-491. |
| [11] | 李钢铁, 秦富仓, 贾守义, 董锦兰, 吉日格勒, 王满贵. 旱生灌木生物量预测模型的研究. 内蒙古林学院学报, 1998, 20(2): 24-31. |
| [16] | 张炜平, 黄聚聪, 李熙波. 杉木林林下植被生物量影响因素. 福建林业科技, 2007, 34(3): 97-99, 149-149. |
| [17] | 郑绍伟, 唐敏, 邹俊辉, 慕长龙. 灌木群落及生物量研究综述. 成都大学学报: 自然科学版, 2007, 26(3): 189-192. |
| [18] | 吴鹏飞, 朱波, 刘世荣, 王小国. 不同林龄桤-柏混交林生态系统的碳储量及其分配. 应用生态学报, 2008, 19(7): 1419-1424. |
| [19] | 朱元恩, 姚冬梅. 宜昌市郊柏木人工林林下植被发育及生物量研究. 林业资源管理, 2007, (2): 52-56. |
| [20] | 王江. 桤柏混交幼林群落特征及生物量调查. 四川林业科技, 1993, 14(1): 66-69. |
| [21] | 刘兴良, 鄢武先, 向成华, 蒋俊民. 沱江流域亚热带次生植被生物量及其模型. 植物生态学报, 1997, 21(5): 50-63. |
| [22] | 曾珍英, 刘琪璟, 曾慧卿. 江西千烟洲几种灌木生物量模型的研究. 福建林业科技, 2005, 32(4): 68-72. |
| [25] | 黎燕琼, 郑绍伟, 龚固堂, 陈俊华, 唐波, 朱自芳, 吴雪仙, 慕长龙. 不同年龄柏木混交林下主要灌木黄荆生物量及分配格局. 生态学报, 2010, 30(11): 2809-2818. |
| [26] | 向成华, 骆宗诗, 陈俊华, 何飞, 罗晓华. 四川盆地丘陵区主要森林群落结构特征研究. 四川林业科技, 2005, 26(5): 25-29. |
| [27] | 郑绍伟, 牛牧, 张琴, 黎燕琼, 慕长龙, 龚固堂, 陈俊华, 朱志芳, 吴雪仙. 川中丘陵区不同类型柏木林地植物群落结构和多样性研究. 四川林业科技, 2011, 32(5): 20-28. |
| [28] | 李荣伟. 长江上游防护林体系建设与经营利用. 成都: 四川科学技术出版社, 2004. |
| [30] | 卢振龙, 龚孝生. 灌木生物量测定的研究进展. 林业调查规划, 2009, 34(4): 37-40, 45-45. |
| [36] | 孙书存, 钱能斌. 刺旋花种群形态参数的通径分析与亚灌木个体生物量建模. 应用生态学报, 1999, 10(2): 155-158. |
| [37] | 曾慧卿, 刘琪璟, 马泽清, 曾珍英. 基于冠幅及植株高度的檵木生物量回归模型. 南京林业大学学报: 自然科学版, 2006, 30(4): 101-104. |
| [38] | 曾慧卿, 刘琪璟, 冯宗炜, 马泽清, 胡理乐. 红壤丘陵区林下灌木生物量估算模型的建立及其应用. 应用生态学报, 2007, 18(10): 2185-2190. |
| [40] | 王蕾, 张宏, 哈斯, 郑秋红, 刘连友. 基于冠幅直径和植株高度的灌木地上生物量估测方法研究. 北京师范大学学报: 自然科学版, 2004, 40(5): 700-704. |
| [42] | 潘攀, 牟长城, 孙志虎. 长白落叶松人工林灌丛生物量的调查与分析. 东北林业大学学报, 2007, 35(04): 1-2, 6-6. |