 2025, Vol. 45
2025, Vol. 45文章信息
- 王真, 刘任涛, 周磊, 徐红伟, 孙建财, 田颖
- WANG Zhen, LIU Rentao, ZHOU Lei, XU Hongwei, SUN Jiancai, TIAN Ying
- 柠条锦鸡儿灌丛排列方式对荒漠草原区地表节肢动物群落结构的影响
- Effects of Caragana korshinskii shrub arrangements on the community structure of ground-active arthropods in desertified steppe
- 生态学报. 2025, 45(7): 3444-3460
- Acta Ecologica Sinica. 2025, 45(7): 3444-3460
- http://dx.doi.org/10.20103/j.stxb.202406101345
-
文章历史
- 收稿日期: 2024-06-10
- 网络出版日期: 2025-01-02






2. 宁夏大学林业与草业学院, 银川 750021;
3. 宁夏大学西北土地退化与生态恢复国家重点实验室培育基地, 银川 750021;
4. 宁夏大学西北退化生态系统恢复与重建教育部重点实验室, 银川 750021;
5. 四川农业大学林学院碳汇研究中心, 成都 611130
2. College of Forestry and Pratacuture, Ningxia University, Yinchuan 750021, China;
3. Breeding Base for State Key Laboratory of Land Degradation and Ecological Restoration in Northwestern China, Ningxia University, Yinchuan 750021, China;
4. Key Laboratory of Restoration and Reconstruction of Degraded Ecosystems in Northwestern China of Ministry of Education, Ningxia University, Yinchuan 750021, China;
5. Carbon Sink Research Center, College of Forestry, Sichuan Agricultural University, Chengdu 611130, China
在气候条件改变和人类活动干扰的背景下, 荒漠草原生态系统易发生退化或沙化现象[1]。建植人工灌丛已成为荒漠草原区减缓荒漠化、提高土地生产力、增加生物多样性和维持生态系统稳定性的有效方式[2], 但如何从时间和空间上进行科学配置已成为当前荒漠化治理最迫切的现实问题[3]。近几十年来, 宁夏盐池县开展了大规模的人工灌丛植被建设, 多采取带状或点状为主的植被排列方式[4], 形成了不同的建植格局。不同植被排列方式的防风效应不同[4—5], 土壤理化性质也存在显著差异[6—8], 而地表节肢动物的栖居和繁殖环境与土壤理化性质密切相关[9]。因而, 不同排列方式的人工植被建设将对地表节肢动物数量特征、活动分布及多样性产生巨大影响。
地表节肢动物是荒漠草原区灌丛生态系统生物多样性的主要组成部分[10—11], 在维持生态系统生态服务功能、生物多样性和食物网结构等方面起着十分关键的作用[12—14]。土壤理化性质等环境因子的变化会直接或间接地影响地表节肢动物群落分布[15—16]及其生态功能[17—18]。已有研究表明, 地表节肢动物群落与土壤含水量、温湿度、pH和养分含量存在显著相关关系[19—20]。但因地域环境差异, 土壤理化性质不同, 地表节肢动物群落的响应模式也不尽相同[9]。目前的研究多集中在农田生态系统, 而对荒漠草原区灌丛生态系统地表节肢动物群落的认识还不足。因此, 在荒漠草原区开展灌丛排列方式对地表节肢动物群落分布和结构特征的影响研究, 对于掌握固沙灌丛林生物分布特性、干旱区流动沙地固定、荒漠生态系统有效恢复及人工灌丛林管理具有重要意义。
目前, 关于灌丛对地表节肢动物影响的研究主要集中在灌丛生境和年龄梯度上[21—23]。研究发现灌丛生境能够维持较高的大型节肢动物多样性和丰富度[24], 灌丛发育过程会对地表节肢动物聚集效应产生显著影响[25], 这表明灌丛时空异质性影响着地表节肢动物的多样性和时空分布特征。然而, 鲜见关于灌丛建植格局对地表节肢动物影响研究。刘晓丽等[26]和常海涛等[27]通过研究发现, 在不同灌丛配置模式下地表节肢动物群落组成会发生改变。综上, 人工植被建植模式会对土壤动物群落组成造成影响, 但是关于其排列方式对地表节肢动物的影响规律的研究较为薄弱, 这直接关系到人工植被排列方式的生态效应及其建植方式选择。
鉴于此, 在宁夏盐池县荒漠草原区选择人工种植的带状和点状排列灌丛为研究对象, 调查在2种灌丛排列方式下地表节肢动物的群落组成、多样性特征及功能群结构, 解析地表节肢动物对灌丛排列的响应规律, 旨在为宁夏荒漠草原人工灌丛建植模式选择、生物多样性保护和荒漠化防治提供依据。
1 研究区概况研究区域位于宁夏回族自治区吴忠市盐池县(37°49′ N, 107°27′ E, 海拔1348 m)。该区域属于典型的中温带大陆性季风气候。年平均气温8.5 ℃, 年均最高气温22.4 ℃, 年均最低气温-8.7 ℃。年均降雨量250—350 mm, 其中60%降雨量集中在7—9月, 年际变率大。年实际蒸散量2136 mm, 年潜在蒸散量1120 mm。地带性土壤类型主要为灰钙土, 非地带性土壤为风沙土, 土壤结构较为疏松, 肥力较低。
2 研究方法 2.1 试验设计于2023年5月、7月和10月, 选择带状和点状2种灌丛排列方式下的柠条锦鸡儿(Caragana korshinskii)人工植被区为研究样地, 间距1000 m以上;2种样地灌木高度和冠幅基本一致, 其中带状排列样地中柠条灌丛平均高度为(122.13±6.39)cm, 平均冠幅为(119.71±4.61)cm, 主要植被包括猪毛蒿(Artemisia scoparia)、艾草(Artemisia argyi)、冷蒿(Artemisia frigida)和牛枝子(Lespedeza potaninii)等, 点状排列样地中柠条灌丛平均高度为(119.48±3.94)cm, 平均冠幅为(120.44±3.11)cm, 主要植被包括艾草(Artemisia argyi)、地梢瓜(Cynanchum thesioides)和雾冰藜(Grubovia dasyphylla)等。
在每种样地设置3个重复, 面积为50 m×50 m, 间距30 m以上。在每个样区随机选择6个调查样点, 每个调查样点之间距离至少为10 m。
共计36个取样点, 即2样地× 3重复样区× 6样点。
2.2 地表节肢动物收集与鉴定采用陷阱诱捕法捕获地表节肢动物, 在每个调查样点布设1个陷阱杯(杯底直径为5 cm, 杯口直径为9 cm, 高度为10 cm), 杯口与地面齐平, 杯中加入防冻液, 以保证诱捕的有效性[28]。每次试验共布设36个陷阱杯, 14天后, 将陷阱杯收回, 并将捕获到的地表节肢动物标本收集置于装有75% 浓度酒精的小白瓶中, 写好标签, 带回实验室进行鉴定统计。
根据《中国土壤动物检索图鉴》[29]和《宁夏贺兰山昆虫》[30]等分类图鉴将采集到的地表节肢动物鉴定到科水平, 少数鉴定到目。根据不同地表节肢动物类群在群落总个体数中所占比例, 将其划分为优势类群(占群落总个体数的10% 以上)、常见类群(1%—10%)和稀有类群(< 1%)[31]。依据地表节肢动物取食类型, 将其划分为捕食性、植食性、杂食性、腐食性、寄生性和菌食性6个营养功能群。由于腐食性、寄生性和菌食性地表节肢动物个体数和类群数较少, 统一归为其他食性。
2.3 植被调查灌木:在每个调查样点附近随机设置1个10 m×10 m的样方, 调查该样方中5株灌丛的冠幅长度(cm)和宽度(cm), 植株高度(cm)。
草本:在每个调查样点附近随机设置1个0.5 m×0.5 m的样方, 调查该样方中草本种类、数量及高度(cm)。
2.4 土壤样品收集与分析在每个取样点按照五点取样法, 用铲子铲取0—15 cm混合土壤样品装入写好标签的自封袋中, 带回实验室进行阴干。将阴干后的土样过2 mm孔径筛, 一部分用于测定土壤pH、电导率和机械组成, 另一部分研磨后过0.25 mm孔径筛, 用于测定土壤有机碳、全氮、全磷和全钾。另外, 在取样点用铝盒采集0—15 cm土壤, 用于测定土壤含水量;用体积为100 cm3的环刀采集原状土壤, 用于测定土壤容重。
土壤pH和电导率(μS/cm)均按水土比为5 ∶ 1浸提后进行测定。土壤机械组成(%)采用Mastersizer 3000激光粒度仪测定[32], 根据美国农部(USDA)制划分土壤质地:黏粒(< 0.002 mm)、粉粒(0.002—0.05 mm)和砂粒(0.05—2 mm)[33]。土壤有机碳(g/kg)用重铬酸钾外加热法测定, 土壤全氮(g/kg)用半微量凯氏法测定, 土壤全钾(g/kg)用NaOH熔融法测定, 土壤全磷(g/kg)用钼锑抗比色法测定[33]。土壤含水量(%)和容重(g/cm3)采用烘干称量法进行测定。
2.5 数据处理地表节肢动物群落多样性以Shannon-Wienner多样性指数(H′)和Simpson优势度指数(C)表示, 计算公式如下:
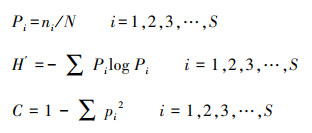
|
式中, Pi为类群i的个体数占群落总个体数的比例;N为群落总个体数;S为类群数。
利用SPSS Statistics 25对数据进行统计分析;分析前对所有数据进行正态分布检验, 不符合的数据采用log(x+1)进行转换;符合正态分布的数据采用独立样本T检验进行分析, 不符合的数据采用非参数检验进行分析。
利用R4.4.0计算地表节肢动物的多样性指数, 并进行主坐标分析(Principal Co-ordinates Analysis, PCoA)、环境因子分析(Distance-based redundancy analysis, db-RDA)和重复测量方差分析(Repeated measures ANOVA);利用Origin 2021绘制地表节肢动物群落指数特征和功能群结构特征图;利用SPSS Statistics 25进行Pearson相关性分析, 计算环境因子与地表节肢动物群落组成、多样性指数和功能类群的相关性。
3 结果与分析 3.1 环境因子由表 1可知, 在3个月份中, 土壤电导率、有机碳、全氮、全磷、黏粒和粉粒均表现为带状显著高于点状(P < 0.001);而土壤全钾和砂粒表现为带状显著低于点状(P < 0.001)。
| 环境指标Environmental factors | 5月May | 7月July | 10月October | ||||||||
| 带状Belt | 点状Point | T | 带状Belt | 点状Point | T | 带状Belt | 点状Point | T | |||
| 土壤含水量Soil moisture content/% | 0.01±0 | 0.02±0 | -4.61*** | 0.01±0 | 0.01±0 | 0.16 | 0.03±0 | 0.02±0 | 2.12* | ||
| 土壤容重Soil bulk density/(g/cm3) | 1.43±0.02 | 1.43±0.02 | 0.14 | 1.31±0.02 | 1.49±0.01 | -6.86*** | 1.39±0.02 | 1.43±0.02 | -1.66 | ||
| 土壤pH Soil potential of hydrogen | 7.73±0.06 | 7.74±0.14 | -0.08 | 7.88±0.02 | 8.16±0.02 | -9.22*** | 7.63±0.04 | 7.89±0.05 | -4.00*** | ||
| 土壤电导率Soil electrical conductivity/(μS/cm) | 88.44±1.71 | 67.82±2.09 | 7.64*** | 66.94±1.5 | 54.56±0.98 | 6.90*** | 76.93±1.86 | 61.02±1.02 | 7.49*** | ||
| 土壤有机碳Soil organic carbon/(g/kg) | 5.13±0.21 | 1.41±0.09 | 16.67*** | 4.25±0.15 | 1.34±0.09 | 16.59*** | 5.18±0.37 | 1.51±0.06 | 9.89*** | ||
| 土壤全氮Soil total nitrogen/(g/kg) | 0.28±0.01 | 0.06±0 | 16.36*** | 0.26±0.01 | 0.07±0 | 16.86*** | 0.33±0.03 | 0.08±0 | 8.89*** | ||
| 土壤全磷Soil total phosphorus/(g/kg) | 0.3±0.01 | 0.18±0.01 | 12.78*** | 0.27±0.01 | 0.2±0 | 10.85*** | 0.29±0.01 | 0.18±0.01 | 8.81*** | ||
| 土壤全钾Soil total kalium/(g/kg) | 18.94±0.06 | 19.48±0.1 | -4.45*** | 19.16±0.1 | 20.1±0.13 | -5.69*** | 19.33±0.11 | 20.57±0.12 | -7.58*** | ||
| 土壤黏粒Soil clay/% | 7.36±0.35 | 1.62±0.1 | 15.69*** | 6.51±0.51 | 1.6±0.16 | 9.17*** | 6.03±0.6 | 1.52±0.28 | 6.85*** | ||
| 土壤粉粒Soil silt/% | 23.99±0.85 | 1.97±0.15 | 25.45*** | 21.38±1.57 | 2.3±0.17 | 12.05*** | 19.75±1.71 | 1.99±0.29 | 10.22*** | ||
| 土壤砂粒Soil sand/% | 68.65±1.15 | 96.41±0.23 | -23.60*** | 72.11±2.07 | 96.1±0.32 | -11.48*** | 74.22±2.27 | 96.49±0.55 | -9.52*** | ||
| 草本丰富度Herb richness | 3.00±0.28 | 2.33±0.11 | 2.20* | 3.94±0.49 | 2.33±0.27 | 2.86** | 2.67±0.28 | 2.22±0.17 | 1.35 | ||
| 草本高度Herb height/m | 2.83±0.22 | 4.82±0.57 | -3.24** | 13.19±1.05 | 18.65±1.37 | -3.17** | 5.65±0.91 | 6.46±3.07 | -0.26 | ||
| 草本密度Shrub density/(株/m2) | 41.33±7.57 | 23.22±2.64 | 2.26* | 60.44±5.11 | 22.56±2.01 | 6.90*** | 38.67±8.37 | 55.67±10.92 | -1.24 | ||
| 灌木高度Shrub height/cm | 68.67±3.4 | 90.22±3.78 | -4.24*** | 148.00±6.43 | 132.67±4.58 | 1.94 | 149.72±8.75 | 135.56±5.9 | 1.34 | ||
| 灌木冠幅Shrubs crown width/cm | 128.58±10.44 | 117.83±4.81 | 0.94 | 119.72±5.37 | 122.5±5.45 | -0.36 | 110.83±7.17 | 121.00±6.07 | -1.08 | ||
| *表示P < 0.05, * *表示P < 0.01, * * *表示P < 0.001 | |||||||||||
另外, 在5月份, 草本丰富度和密度也表现为带状显著高于点状(P < 0.05);而土壤含水量、草本高度和灌木高度表现为带状显著低于点状(P < 0.01)。在7月份, 草本丰富度和密度也表现为带状显著高于点状(P < 0.01), 而土壤容重、pH和草本高度表现为带状显著低于点状(P < 0.01)。在10月份, 土壤含水量也表现为带状显著高于点状(P < 0.05), 而土壤pH表现为带状显著低于点状(P < 0.001)。
3.2 地表节肢动物群落组成由表 2可知, 在2种排列灌丛样地共捕获地表节肢动物3408只, 隶属于10目54科56个类群, 其中蚁科(Formicidae)、拟步甲科(Tenebrionidae)和步甲科(Carabidae)为优势类群, 其个体数分别占总捕获量的33.22%、14.52%、12.32%;其中长蝽科(Lygaeidae)、光盔蛛科(Liocranidae)、平腹蛛科(Gnaphosidae)、跳蛛科(Salticidae)、花蝽科(Anthocoridae)、象甲科(Curculioninae)、盾蝽科(Scutelleridae)、缘蝽科(Coreidae)、金龟科(Scarabaeidae)、叶蝉科(Cicadellidae)、狼栉蛛科(Zoridae)和吉丁甲科(Buprestidae)12个类群属于常见类群, 其个体数共占总捕获量的32.39%;其余41个类群属于稀有类群, 其个体数共占总捕获量的8.02%。
| 物种Species | 功能类群Functional groups | 5月May | 7月July | 10月October | 总计Total | |||||||||||||||||||||||||||||
| 带状Belt | 点状Point | 带状Belt | 点状Point | 带状Belt | 点状Point | |||||||||||||||||||||||||||||
| 个体数IN | % | 优势度Adv | 个体数IN | % | 优势度Adv | 个体数IN | % | 优势度Adv | 个体数IN | % | 优势度Adv | 个体数IN | % | 优势度Adv | 个体数IN | % | 优势度Adv | 个体数IN | % | 优势度Adv | ||||||||||||||
| 螯螋科Chelisochidae | O | 1 | 0.17 | + | 0 | 0.00 | + | 1 | 0.12 | + | 24 | 2.77 | ++ | 0 | 0.00 | + | 0 | 0.00 | + | 26 | 0.76 | + | ||||||||||||
| 步甲科Carabidae | Pr | 10 | 1.67 | ++ | 15 | 2.92 | ++ | 71 | 8.28 | ++ | 148 | 17.07 | +++ | 103 | 28.77 | +++ | 73 | 34.27 | +++ | 420 | 12.32 | +++ | ||||||||||||
| 蝽科Pentatomidae | Ph | 1 | 0.17 | + | 0 | 0.00 | + | 1 | 0.12 | + | 1 | 0.12 | + | 0 | 0.00 | + | 0 | 0.00 | + | 3 | 0.09 | + | ||||||||||||
| 粗螯蛛科Prodidomidae | Pr | 0 | 0.00 | + | 1 | 0.19 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.03 | + | ||||||||||||
| 盾蝽科Scutelleridae | Ph | 21 | 3.51 | ++ | 0 | 0.00 | + | 46 | 5.37 | ++ | 2 | 0.23 | + | 1 | 0.28 | + | 0 | 0.00 | + | 70 | 2.05 | ++ | ||||||||||||
| 粉蠹科Lyctidae | O | 1 | 0.17 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.03 | + | ||||||||||||
| 粪金龟科Geotrupidae | S | 1 | 0.17 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.03 | + | ||||||||||||
| 蜉金龟科Aphodiidae | S | 13 | 2.17 | ++ | 0 | 0.00 | + | 8 | 0.93 | + | 6 | 0.69 | + | 0 | 0.00 | + | 0 | 0.00 | + | 27 | 0.79 | + | ||||||||||||
| 管巢蛛科Clubionidae | Pr | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.12 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.03 | + | ||||||||||||
| 光盔蛛科Liocranidae | Pr | 27 | 4.51 | ++ | 63 | 12.26 | +++ | 44 | 5.13 | ++ | 17 | 1.96 | ++ | 3 | 0.84 | + | 4 | 1.88 | ++ | 158 | 4.64 | ++ | ||||||||||||
| 红蝽科Pyrrhocoridae | Ph | 0 | 0.00 | + | 0 | 0.00 | + | 5 | 0.58 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 5 | 0.15 | + | ||||||||||||
| 虎甲科Cicindelidae | Pr | 0 | 0.00 | + | 0 | 0.00 | + | 11 | 1.28 | ++ | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 11 | 0.32 | + | ||||||||||||
| 花蝽科Anthocoridae | Pr | 45 | 7.51 | ++ | 4 | 0.78 | + | 10 | 1.17 | ++ | 4 | 0.46 | + | 20 | 5.59 | ++ | 3 | 1.41 | ++ | 86 | 2.52 | ++ | ||||||||||||
| 花蚤科Mordellidae | Ph | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.12 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.03 | + | ||||||||||||
| 吉丁甲科Buprestidae | Ph | 27 | 4.51 | ++ | 8 | 1.56 | ++ | 1 | 0.12 | + | 1 | 0.12 | + | 2 | 0.56 | + | 0 | 0.00 | + | 39 | 1.14 | ++ | ||||||||||||
| 尖翅蝇科Lonchopteridae | O | 1 | 0.17 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.03 | + | ||||||||||||
| 金龟科Scarabaeidae | Ph | 5 | 0.83 | + | 1 | 0.19 | + | 1 | 0.12 | + | 0 | 0.00 | + | 32 | 8.94 | ++ | 20 | 9.39 | ++ | 59 | 1.73 | ++ | ||||||||||||
| 叩甲科Elaterinae | Ph | 0 | 0.00 | + | 2 | 0.39 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 2 | 0.06 | + | ||||||||||||
| 蜡蝉科若虫Fulgoridae | Ph | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.12 | + | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.03 | + | ||||||||||||
| 狼栉蛛科Zoridae | Pr | 6 | 1.00 | ++ | 16 | 3.11 | ++ | 4 | 0.47 | + | 18 | 2.08 | ++ | 3 | 0.84 | + | 0 | 0.00 | + | 47 | 1.38 | ++ | ||||||||||||
| 狼蛛科Lycosidae | Pr | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.12 | + | 0 | 0.00 | + | 1 | 0.28 | + | 0 | 0.00 | + | 2 | 0.06 | + | ||||||||||||
| 蝼蛄总科Gryllotalpoidea | Ph | 1 | 0.17 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.03 | + | ||||||||||||
| 蜢总科Eumastacoidea | Ph | 0 | 0.00 | + | 0 | 0.00 | + | 2 | 0.23 | + | 0 | 0.00 | + | 8 | 2.23 | ++ | 0 | 0.00 | + | 10 | 0.29 | + | ||||||||||||
| 膜蝽科Hebridae | Ph | 1 | 0.17 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.03 | + | ||||||||||||
| 泥蜂科Sphecidae | Pr | 1 | 0.17 | + | 0 | 0.00 | + | 3 | 0.35 | + | 4 | 0.46 | + | 0 | 0.00 | + | 0 | 0.00 | + | 8 | 0.23 | + | ||||||||||||
| 拟步甲科Tenebrionidae | Ph | 94 | 15.69 | +++ | 203 | 39.49 | +++ | 64 | 7.47 | ++ | 77 | 8.88 | ++ | 6 | 1.68 | ++ | 51 | 23.94 | +++ | 495 | 14.52 | +++ | ||||||||||||
| 瓢虫科Coccinellidae | Pr | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.28 | + | 1 | 0.47 | + | 2 | 0.06 | + | ||||||||||||
| 平腹蛛科Gnaphosidae | Pr | 6 | 1.00 | ++ | 52 | 10.12 | +++ | 13 | 1.52 | ++ | 62 | 7.15 | ++ | 12 | 3.35 | ++ | 9 | 4.23 | ++ | 154 | 4.52 | ++ | ||||||||||||
| 奇蝽科Enicocephalidae | Ph | 1 | 0.17 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.03 | + | ||||||||||||
| 鳃金龟科Melolonthidae | Ph | 8 | 1.34 | ++ | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 8 | 0.23 | + | ||||||||||||
| 食蚜蝇科Syrphidae | O | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 9 | 1.04 | ++ | 0 | 0.00 | + | 0 | 0.00 | + | 9 | 0.26 | + | ||||||||||||
| 硕螽科Bradyporidae | O | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 5 | 0.58 | + | 0 | 0.00 | + | 0 | 0.00 | + | 5 | 0.15 | + | ||||||||||||
| 跳蛛科Salticidae | Pr | 16 | 2.67 | ++ | 31 | 6.03 | ++ | 10 | 1.17 | ++ | 51 | 5.88 | ++ | 3 | 0.84 | + | 8 | 3.76 | ++ | 119 | 3.49 | ++ | ||||||||||||
| 蜈蚣目Scolopendromorpha | Pr | 1 | 0.17 | + | 1 | 0.19 | + | 2 | 0.23 | + | 8 | 0.92 | + | 1 | 0.28 | + | 0 | 0.00 | + | 13 | 0.38 | + | ||||||||||||
| 蟋蟀总科Grylloidea | O | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.12 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.03 | + | ||||||||||||
| 象虫科Curculionidae | Ph | 1 | 0.17 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.28 | + | 0 | 0.00 | + | 2 | 0.06 | + | ||||||||||||
| 象甲科Curculioninae | Ph | 22 | 3.67 | ++ | 26 | 5.06 | ++ | 1 | 0.12 | + | 0 | 0.00 | + | 23 | 6.42 | ++ | 10 | 4.69 | ++ | 82 | 2.41 | ++ | ||||||||||||
| 逍遥蛛科Philodromidae | Pr | 6 | 1.00 | ++ | 3 | 0.58 | + | 8 | 0.93 | + | 3 | 0.35 | + | 0 | 0.00 | + | 0 | 0.00 | + | 20 | 0.59 | + | ||||||||||||
| 小蠹科Scolytidae | Ph | 0 | 0.00 | + | 0 | 0.00 | + | 3 | 0.35 | + | 5 | 0.58 | + | 0 | 0.00 | + | 0 | 0.00 | + | 8 | 0.23 | + | ||||||||||||
| 小蕈甲科Mycetophagidae | F | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.28 | + | 0 | 0.00 | + | 1 | 0.03 | + | ||||||||||||
| 蟹蛛科Thomisidae | Pr | 0 | 0.00 | + | 0 | 0.00 | + | 2 | 0.23 | + | 0 | 0.00 | + | 11 | 3.07 | ++ | 7 | 3.29 | ++ | 20 | 0.59 | + | ||||||||||||
| 芫菁科Meloidae | Ph | 0 | 0.00 | + | 0 | 0.00 | + | 2 | 0.23 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 2 | 0.06 | + | ||||||||||||
| 阎甲科Histeridae | Pr | 1 | 0.17 | + | 5 | 0.97 | + | 1 | 0.12 | + | 2 | 0.23 | + | 1 | 0.28 | + | 0 | 0.00 | + | 10 | 0.29 | + | ||||||||||||
| 叶蝉科Cicadellidae | Ph | 21 | 3.51 | ++ | 1 | 0.19 | + | 12 | 1.40 | ++ | 0 | 0.00 | + | 17 | 4.75 | ++ | 0 | 0.00 | + | 51 | 1.50 | ++ | ||||||||||||
| 蚁科Formicidae | O | 94 | 15.69 | +++ | 64 | 12.45 | +++ | 456 | 53.21 | +++ | 403 | 46.48 | +++ | 92 | 25.70 | +++ | 23 | 10.80 | +++ | 1132 | 33.22 | +++ | ||||||||||||
| 蚁蛉科Myrmeleontidae | Pr | 0 | 0.00 | + | 4 | 0.78 | + | 1 | 0.12 | + | 0 | 0.00 | + | 0 | 0.00 | + | 3 | 1.41 | ++ | 8 | 0.23 | + | ||||||||||||
| 蚁形甲科Anthicidae | O | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.12 | + | 0 | 0.00 | + | 1 | 0.28 | + | 0 | 0.00 | + | 2 | 0.06 | + | ||||||||||||
| 隐翅甲科Staphylinidae | S | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 3 | 0.84 | + | 0 | 0.00 | + | 3 | 0.09 | + | ||||||||||||
| 缨甲科Ptiliidae | F | 0 | 0.00 | + | 1 | 0.19 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.03 | + | ||||||||||||
| 蚰蜒目Scutigeromorpha | Pr | 3 | 0.50 | + | 3 | 0.58 | + | 16 | 1.87 | ++ | 6 | 0.69 | + | 2 | 0.56 | + | 0 | 0.00 | + | 30 | 0.88 | + | ||||||||||||
| 圆蛛科Araneidae | Pr | 0 | 0.00 | + | 1 | 0.19 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.03 | + | ||||||||||||
| 缘蝽科Coreidae | Ph | 9 | 1.50 | ++ | 0 | 0.00 | + | 41 | 4.78 | ++ | 3 | 0.35 | + | 7 | 1.96 | ++ | 0 | 0.00 | + | 60 | 1.76 | ++ | ||||||||||||
| 蚱总科Tetrigoidea | Ph | 0 | 0.00 | + | 1 | 0.19 | + | 0 | 0.00 | + | 2 | 0.23 | + | 0 | 0.00 | + | 0 | 0.00 | + | 3 | 0.09 | + | ||||||||||||
| 长蝽科Lygaeidae | Ph | 154 | 25.71 | +++ | 6 | 1.17 | ++ | 11 | 1.28 | ++ | 4 | 0.46 | + | 3 | 0.84 | + | 1 | 0.47 | + | 179 | 5.25 | ++ | ||||||||||||
| 栉蝽科Ceratocombidae | Ph | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.12 | + | 0 | 0.00 | + | 0 | 0.00 | + | 1 | 0.03 | + | ||||||||||||
| 肿腿蜂科Bethylidae | Pa | 0 | 0.00 | + | 2 | 0.39 | + | 1 | 0.12 | + | 0 | 0.00 | + | 0 | 0.00 | + | 0 | 0.00 | + | 3 | 0.09 | + | ||||||||||||
| 总计Total | 599 | 100.00 | 514 | 100.00 | 857 | 100.00 | 867 | 100.00 | 358 | 100.00 | 213 | 100.00 | 3408 | 100.00 | ||||||||||||||||||||
| IN:个体数Individual number;Adv:优势度Advantage; +++:优势类群Dominant group(>10%);++:常见类群Common group(1%—10%);+:稀有类群Rare group(< 1%); Pr:捕食性Predatory;Ph:植食性Phytophagous;O:杂食性Omnivorous;S:腐食性Saprozoic;F:菌食性Fungivorous;Pa: 寄生性Parasitic | ||||||||||||||||||||||||||||||||||
在2种排列灌丛中所捕获到的地表节肢动物的数量和种类不同。其中带状排列灌丛共捕获地表节肢动物1814只, 隶属于10目45科47个类群;点状排列灌丛共捕获地表节肢动物1594只, 隶属于10目35科37个类群, 相较于带状, 其个体数减少了22只, 类群数减少了10个。
在3个月份中, 蚁科为2种排列灌丛中共有的优势类群, 但在2种排列灌丛中地表节肢动物优势类群的类别并不相同。5月份, 带状优势类群包括长蝽科、蚁科和拟步甲科, 点状优势类群包括拟步甲科、蚁科、光盔蛛科和平腹蛛科, 相较于带状, 其优势类群增加了1个;7月份, 带状优势类群包括蚁科, 点状优势类群包括蚁科和步甲科, 相较于带状, 其优势类群增加了1个;10月份, 带状优势类群包括步甲科和蚁科, 点状优势类群包括步甲科、拟步甲科和蚁科, 相较于带状, 其优势类群增加了1个。
由表 3可知, 优势类群的类群数在5、7月份均表现为带状显著低于点状(P < 0.05);常见类群的个体数和类群数在5、10月份均表现为带状显著高于点状(P < 0.05);稀有类群的个体数和类群数在7、10月份均表现为带状显著高于点状(P < 0.05)。
| 类群Group | 5月May | 7月July | 10月October | ||||||
| 带状Belt | 点状Point | 带状Belt | 点状Point | 带状Belt | 点状Point | ||||
| 优势类群Dominant group | 个体数IN | 19.00±2.5 | 21.22±2.34 | 25.33±0.34 | 30.61±6.43 | 10.83±2.37 | 8.17±0.95 | ||
| 类群数GR | 3.06±0.06b | 3.39±0.14a | 1.00±0b | 1.94±0.06a | 1.94±0.1 | 2.17±0.17 | |||
| 常见类群Common group | 个体数IN | 13.17±1.47a | 5.67±0.56b | 11.50±1.19 | 14.33±1.21 | 7.56±0.84a | 3.56±0.61b | ||
| 类群数GR | 6.83±0.46a | 3.22±0.22b | 3.72±0.31 | 4.33±0.24 | 4.11±0.35a | 2.17±0.33b | |||
| 稀有类群Rare group | 个体数IN | 1.11±0.2 | 1.67±0.35 | 10.78±1.13a | 3.22±0.53b | 1.50±0.42a | 0.11±0.08b | ||
| 类群数GR | 1.11±0.2 | 1.39±0.29 | 5.61±0.51a | 2.39±0.37b | 1.33±0.37a | 0.11±0.08b | |||
| IN:个体数Individual number;GR:类群数Group richness; 不同字母表示在相同月份2种排列方式间存在显著差异(P < 0.05) | |||||||||
由图 1可知, 带状拟步甲科个体数显著低于点状(P < 0.001);另外, 带状蚁科个体数高于点状, 步甲科个体数低于点状, 但在2种排列灌丛间并无显著差异。

|
| 图 1 蚁科、拟步甲科和步甲科个体数 Fig. 1 Individual number of Formicidae, Tenebrionidae and Carabidae *表示P < 0.05, * *表示P < 0.01, * * *表示P < 0.001 |
由表 4可知, 采样时间对优势类群个体数有显著影响(P < 0.001), 此外, 灌丛排列方式、采样时间及二者交互作用均对优势类群类群数、常见类群个体数和类群数、稀有类群个体数和类群数具有显著影响(P < 0.01)。
| 优势类群Dominant group | 常见类群Common group | 稀有类群Rare group | |||||||||||||||
| 个体数 Individual number |
类群数 Group richness |
个体数 Individual number |
类群数 Group richness |
个体数 Individual number |
类群数 Group richness |
||||||||||||
| F | P | F | P | F | P | F | P | F | P | F | P | ||||||
| 处理Treatment | 0.391 | 35.108 | *** | 11.71 | *** | 37.88 | *** | 36.73 | *** | 26.25 | *** | ||||||
| 月份Month | 17.264 | *** | 148.666 | *** | 25.38 | *** | 16.6 | *** | 73.28 | *** | 56.2 | *** | |||||
| 处理×月份T×M | 0.807 | 7.079 | ** | 12.92 | *** | 21.02 | *** | 28.08 | *** | 13.98 | *** | ||||||
| *表示P < 0.05, * *表示P < 0.01, * * *表示P < 0.001 | |||||||||||||||||
主坐标分析(图 2)显示, 两种排列灌丛间地表节肢动物群落结构不同, 且随季节动态变化。PCoA1和PCoA2分别解释了地表节肢动物群落变化的30.62%和26.18%。其中, 5月份带状和点状的地表节肢动物群落结构存在显著差异, 但在7月和10月显示出相似的群落结构。

|
| 图 2 地表节肢动物群落组成主坐标分析(PCoA) 排序图 Fig. 2 Principal co-ordinates analysis (PCoA) ranking diagram of the community composition of ground-active arthropods 不同字母表示在相同月份2种排列方式间存在显著差异(P < 0.05) |
由图 3可知, 地表节肢动物个体数在10月份表现为带状显著高于点状(P < 0.05), 类群数在3个月份均表现为带状显著高于点状(P < 0.05), Shannon-Wienner多样性指数在5、10月份均表现为带状显著高于点状(P < 0.01), 而Simpson优势度指数在5月份表现为带状显著低于点状(P < 0.01)。

|
| 图 3 地表节肢动物群落指数特征 Fig. 3 Community index of ground-active arthropods *表示P < 0.05, * *表示P < 0.01, * * *表示P < 0.001 |
由表 5可知, 灌丛排列方式对类群数和Shannon-Wienner多样性指数有显著影响(P < 0.01);采样时间对个体数、类群数、Shannon-Wienner多样性指数和Simpson优势度指数均有显著影响(P < 0.01);二者交互作用对Shannon-Wienner多样性指数和Simpson优势度指数有显著影响(P < 0.05)。
| 个体数 Individual number |
类群数 Group richness |
多样性指数 Shannon-Wienner index |
优势度指数 Simpson index |
||||||||
| F | P | F | P | F | P | F | P | ||||
| 处理Treatment | 2.097 | 38.996 | *** | 12.439 | ** | 2.059 | |||||
| 月份Month | 42.856 | *** | 35.885 | *** | 10.948 | *** | 5.38 | ** | |||
| 处理×月份T×M | 0.788 | 1.085 | 4.247 | * | 3.032 | * | |||||
| *表示P < 0.05, * *表示P < 0.01, * * *表示P < 0.001 | |||||||||||
将所捕获的地表节肢动物根据取食类型进行划分, 可分为捕食性、植食性、杂食性、腐食性、寄生性和菌食性6个功能类群, 由表 2可知, 其个体数分别占总个体数的32.60%、31.81%、34.54%、0.91%、0.09%和0.06%, 其中捕食性包含19个类群, 植食性包含23个类群, 杂食性包含8个类群, 腐食性包含3个类群, 寄生性包含1个类群, 菌食性包含2个类群。杂食性、捕食性和植食性的个体数量相对较高, 植食性和捕食性类群种类相对丰富, 而腐食性、寄生性和菌食性类群的个体数量和种类均相对较低, 将腐食性、寄生性和菌食性归为其他食性。
由图 4可知, 捕食性个体数在5、7月份均表现为带状显著低于点状(P < 0.05);而植食性个体数在5、7月份均表现为带状显著高于点状(P < 0.05);杂食性个体数在10月份表现为带状显著高于点状(P < 0.001);其他食性个体数在5月份表现为带状显著高于点状(P < 0.05)。

|
| 图 4 地表节肢动物功能类群个体数 Fig. 4 Individual number in functional groups of ground-active arthropods *表示P < 0.05, * *表示P < 0.01, * * *表示P < 0.001 |
由图 5可知, 捕食性类群数在5月份表现为带状显著低于点状(P < 0.01);而植食性类群数在5、7、10月份均表现为带状显著高于点状(P < 0.001);杂食性类群数在5、10月份均表现为带状显著高于点状(P < 0.01), 但在7月份表现为带状显著低于点状(P < 0.01);其他食性类群数在5月份表现为带状显著高于点状(P < 0.05)。

|
| 图 5 地表节肢动物功能类群类群数 Fig. 5 Group richness in functional groups of ground-active arthropod *表示P < 0.05, * *表示P < 0.01, * * *表示P < 0.001 |
由表 6可知, 灌丛排列方式对植食性个体数和类群数、其他食性个体数和类群数具有显著影响(P < 0.01);采样时间对捕食性个体数和类群数、植食性个体数和类群数、杂食性个体数和类群数、其他食性个体数具有显著影响(P < 0.05);二者交互作用对捕食性个体数和类群数、植食性类群数、杂食性类群数具有显著影响(P < 0.05)。
| 捕食性Predatory | 植食性Phytophagous | ||||||||||
| 个体数IN | 类群数GR | 个体数IN | 类群数GR | ||||||||
| F | P | F | P | F | P | F | P | ||||
| 处理Treatment | 3.560 | 0.729 | 14.572 | *** | 128.236 | *** | |||||
| 月份Month | 8.515 | *** | 20.570 | *** | 41.889 | *** | 27.198 | *** | |||
| 处理×月份T×M | 4.080 | * | 4.816 | * | 2.252 | 8.273 | *** | ||||
| 杂食性Omnivorous | 其他食性Other dietary habits | ||||||||||
| 个体数IN | 类群数GR | 个体数IN | 类群数GR | ||||||||
| F | P | F | P | F | P | F | P | ||||
| 处理Treatment | 1.024 | 2.663 | 7.423 | ** | 7.621 | ** | |||||
| 月份Month | 40.657 | *** | 20.946 | *** | 3.368 | * | 2.57 | ||||
| 处理×月份T×M | 0.15 | 16.868 | *** | 1.306 | 1.668 | ||||||
| IN:个体数Individual number;GR:类群数Group richness; *表示P < 0.05, * *表示P < 0.01, * * *表示P < 0.001 | |||||||||||
采用方差膨胀因子分析(Variance Inflation Factor)剔除VIF >2的环境因子, 以排除环境因子共线性的影响。由图 6可知, 排序轴1解释了33.17%的生境变化, 排序轴2解释了26.74%的生境变化。通过表 7可知, 影响带状和点状排列灌丛间物种组成差异的主要环境因子包括土壤容重、pH和草本丰富度。

|
| 图 6 环境因子分析(dbRDA) 排序图 Fig. 6 Distance-based redundancy analysis (dbRDA) ranking diagram Carabidae : 步甲科; Scutelleridae : 盾蝽科; Aphodiidae : 蜉金龟科; Liocranidae : 光盔蛛科; Anthocoridae : 花蝽科; Buprestidae : 吉丁甲科; Scarabaeidae : 金龟科; Zoridae : 狼栉蛛科; Tenebrionidae : 拟步甲科; Gnaphosidae : 平腹蛛科; Melolonthidae : 鳃金龟科; Salticidae : 跳蛛科; Cicadellidae : 叶蝉科; Formicidae : 蚁科; Coreidae : 缘蝽科; Lygaeidae : 长蝽科; BD : 土壤容重Soil bulk density; pH : 土壤酸碱度Soil potential of hydrogen; HR : 草本丰富度Herb richness; HH : 草本高度Herb height; HD : 草本密度Herb density; SC : 灌木冠幅Shrub crown width |
| 环境指标Environmental factors | R2 | P | 环境指标Environmental factors | R2 | P | |
| 土壤容重Soil bulk density | 0.45 | 0.001 | 草本高度Herb height | 0.22 | 0.001 | |
| 土壤pH Soil potential of hydrogen | 0.44 | 0.001 | 草本密度Shrub density | 0.11 | 0.002 | |
| 草本丰富度Herb richness | 0.38 | 0.001 | 灌木冠幅Shrubs crown width | 0.06 | 0.034 |
由表 8可知, 优势类群个体数与草本高度呈显著正相关(P < 0.01), 与土壤含水量和电导率呈显著负相关(P < 0.05)。常见类群个体数与土壤电导率、有机碳、全氮、全磷、黏粒和粉粒呈显著正相关(P < 0.01), 与土壤含水量、全钾、砂粒和灌木高度呈显著负相关(P < 0.01)。稀有类群个体数与草本高度和灌木高度呈显著正相关(P < 0.05)。
| 环境指标 Environmental factors |
优势类群 Dominant group |
常见类群 Common group |
稀有类群 Rare group |
优势物种 Dominant species |
||||||||
| 个体数 IN |
类群数 GR |
个体数 IN |
类群数 GR |
个体数 IN |
类群数 GR |
蚁科 Formicidae |
拟步甲科 Tenebrionidae |
步甲科 Carabidae |
||||
| 土壤含水量Soil moisture content | -0.24* | 0.06 | -0.53** | -0.51** | -0.15 | -0.23* | -0.28** | -0.28** | 0.02 | |||
| 土壤容重Soil bulk density | 0.15 | 0.21* | 0.09 | -0.07 | -0.06 | -0.14 | 0.10 | 0.11 | -0.01 | |||
| 土壤pH Soil potential of hydrogen | 0.14 | 0.24* | -0.03 | -0.11 | 0.15 | 0.10 | 0.09 | 0.02 | 0.08 | |||
| 土壤电导率Soil electrical conductivity | -0.29** | -0.53** | 0.38** | 0.51** | -0.18 | -0.12 | -0.10 | -0.06 | -0.27** | |||
| 土壤有机碳Soil organic carbon | -0.14 | -0.48** | 0.36** | 0.54** | -0.08 | -0.02 | 0.08 | -0.32** | -0.07 | |||
| 土壤全氮Soil total nitrogen | -0.15 | -0.47** | 0.28** | 0.46** | -0.06 | -0.02 | 0.07 | -0.40** | -0.05 | |||
| 土壤全磷Soil total phosphorus | -0.07 | -0.43** | 0.39** | 0.51** | -0.05 | -0.03 | 0.13 | -0.32** | -0.03 | |||
| 土壤全钾Soil total kalium | 0.05 | 0.44** | -0.44** | -0.59** | 0.00 | -0.08 | -0.09 | -0.09 | 0.19* | |||
| 土壤黏粒Soil clay | -0.15 | -0.49** | 0.39** | 0.50** | -0.03 | 0.04 | 0.11 | -0.31** | -0.18 | |||
| 土壤粉粒Soil silt | -0.14 | -0.52** | 0.42** | 0.56** | -0.01 | 0.06 | 0.11 | -0.33** | -0.13 | |||
| 土壤砂粒Soil sand | 0.15 | 0.52** | -0.42** | -0.55** | 0.02 | -0.05 | -0.11 | 0.33** | 0.14 | |||
| 草本丰富度Herb richness | -0.07 | -0.17 | 0.17 | 0.29** | 0.13 | 0.10 | 0.04 | -0.09 | -0.13 | |||
| 草本高度Herb height | 0.29** | 0.34** | -0.15 | -0.19* | 0.23* | 0.20* | 0.18 | -0.03 | 0.27** | |||
| 草本密度Shrub density | -0.09 | -0.29** | 0.06 | 0.09 | 0.03 | 0.09 | -0.07 | -0.19 | 0.21* | |||
| 灌木高度Shrub height | 0.10 | 0.19 | -0.40** | -0.38** | 0.20* | 0.16 | 0.05 | -0.41** | 0.37** | |||
| 灌木冠幅Shrubs crown width | 0.01 | 0.04 | 0.08 | 0.08 | 0.07 | 0.12 | -0.06 | 0.12 | 0.12 | |||
| IN:个体数Individual number;GR:类群数Group richness; *表示P < 0.05, * *表示P < 0.01, * * *表示P < 0.001 | ||||||||||||
优势类群的类群数与土壤容重、pH、全钾、砂粒和草本高度呈显著正相关(P < 0.05), 与土壤电导率、有机碳、全氮、全磷、黏粒、粉粒和草本密度呈显著负相关(P < 0.01)。常见类群的类群数与土壤电导率、有机碳、全氮、全磷、黏粒和粉粒呈显著正相关(P < 0.01), 与土壤含水量、全钾、砂粒和灌木高度呈显著负相关(P < 0.01)。稀有类群的类群数与草本高度呈显著正相关(P < 0.05), 与土壤含水量呈显著负相关(P < 0.05)。
蚁科个体数与土壤含水量呈显著负相关(P < 0.05)。拟步甲科个体数与土壤砂粒呈显著正相关(P < 0.01), 而与土壤含水量、有机碳、全氮、全磷、黏粒、粉粒和灌木高度呈显著负相关(P < 0.05)。步甲科个体数与土壤全钾、草本高度和密度以及灌木高度呈显著正相关(P < 0.05), 而与土壤电导率呈显著负相关(P < 0.01)。
由表 9可知, 地表节肢动物个体数、类群数和Shannon-Wiener指数均与土壤含水量和全磷呈显著负相关(P < 0.05);另外, 类群数还与土壤电导率、有机碳、全氮、全磷、黏粒、粉粒和草本丰富度呈显著正相关(P < 0.05), 与土壤砂粒和灌木高度呈显著负相关(P < 0.05);Shannon-Wiener指数还与土壤有机碳和粉粒呈显著正相关(P < 0.01)。Simpson指数与土壤含水量呈显著正相关(P < 0.05)。
| 环境指标 Environmental factors |
个体数 Individual number |
类群数 Group richness |
多样性指数 Shannon-Wiener index |
优势度指数 Simpson index |
| 土壤含水量Soil moisture content | -0.47** | -0.54** | -0.39** | 0.25* |
| 土壤容重Soil bulk density | 0.16 | -0.08 | -0.03 | -0.05 |
| 土壤pH Soil potential of hydrogen | 0.12 | 0.01 | 0.04 | -0.05 |
| 土壤电导率Soil electrical conductivity | -0.05 | 0.25* | 0.17 | -0.10 |
| 土壤有机碳Soil organic carbon | 0.07 | 0.33** | 0.20* | -0.08 |
| 土壤全氮Soil total nitrogen | 0.03 | 0.26** | 0.13 | -0.01 |
| 土壤全磷Soil total phosphorus | 0.16 | 0.31** | 0.15 | -0.02 |
| 土壤全钾Soil total kalium | -0.20* | -0.43** | -0.24* | 0.07 |
| 土壤黏粒Soil clay | 0.10 | 0.32** | 0.14 | 0.02 |
| 土壤粉粒Soil silt | 0.12 | 0.38** | 0.19* | -0.03 |
| 土壤砂粒Soil sand | -0.12 | -0.37** | -0.18 | 0.02 |
| 草本丰富度Herb richness | 0.07 | 0.25** | 0.15 | -0.04 |
| 草本高度Herb height | 0.18 | 0.02 | -0.01 | 0.04 |
| 草本密度Shrub density | -0.04 | 0.05 | 0.04 | -0.07 |
| 灌木高度Shrub height | -0.09 | -0.20* | -0.17 | 0.13 |
| 灌木冠幅Shrubs crown width | 0.06 | 0.14 | 0.14 | -0.13 |
| *表示P < 0.05, * *表示P < 0.01, * * *表示P < 0.001 | ||||
由表 10可知, 捕食性个体数与土壤砂粒和草本高度呈显著正相关(P < 0.05), 与土壤电导率、黏粒和粉粒呈显著负相关(P < 0.05)。植食性个体数与土壤电导率、有机碳、全磷、黏粒和粉粒呈显著正相关(P < 0.05), 与土壤含水量、全钾、砂粒、草本高度和灌木高度呈显著负相关(P < 0.05)。杂食性个体数与草本高度呈显著正相关(P < 0.05), 与土壤含水量呈显著负相关(P < 0.01)。其他食性个体数与土壤电导率、有机碳、全氮、全磷、黏粒和粉粒含量呈显著正相关(P < 0.05), 与土壤砂粒含量呈显著负相关(P < 0.01)。
| 环境指标 Environmental factors |
捕食性 Predatory |
植食性 Phytophagous |
杂食性 Omnivorous |
其他食性 Other dietary habits |
|||||||
| 个体数 IN |
类群数 GR |
个体数 IN |
类群数 GR |
个体数 IN |
类群数 GR |
个体数 IN |
类群数 GR |
||||
| 土壤含水量Soil moisture content | -0.17 | -0.31** | -0.45** | -0.44** | -0.29** | -0.23* | -0.12 | -0.18 | |||
| 土壤容重Soil bulk density | 0.05 | 0.00 | 0.12 | -0.15 | 0.12 | 0.08 | 0.02 | -0.05 | |||
| 土壤pH Soil potential of hydrogen | 0.18 | 0.21* | -0.12 | -0.29** | 0.12 | 0.27** | 0.01 | 0.01 | |||
| 土壤电导率Soil electrical conductivity | -0.29** | -0.22* | 0.37** | 0.58** | -0.12 | -0.04 | 0.21* | 0.21* | |||
| 土壤有机碳Soil organic carbon | -0.18 | -0.14 | 0.214* | 0.58** | 0.06 | 0.10 | 0.25** | 0.25** | |||
| 土壤全氮Soil total nitrogen | -0.18 | -0.16 | 0.12 | 0.50** | 0.05 | 0.09 | 0.21* | 0.20* | |||
| 土壤全磷Soil total phosphorus | -0.10 | -0.12 | 0.24* | 0.50** | 0.12 | 0.17 | 0.23* | 0.21* | |||
| 土壤全钾Soil total kalium | 0.10 | -0.11 | -0.42** | -0.53** | -0.06 | -0.07 | -0.18 | -0.17 | |||
| 土壤黏粒Soil clay | -0.23* | -0.14 | 0.25** | 0.53** | 0.09 | 0.16 | 0.28** | 0.26** | |||
| 土壤粉粒Soil silt | -0.19* | -0.11 | 0.27** | 0.59** | 0.09 | 0.17 | 0.31** | 0.28** | |||
| 土壤砂粒Soil sand | 0.20* | 0.12 | -0.26** | -0.58** | -0.09 | -0.17 | -0.31** | -0.28** | |||
| 草本丰富度Herb richness | -0.06 | 0.17 | 0.10 | 0.18 | 0.04 | 0.09 | 0.13 | 0.14 | |||
| 草本高度Herb height | 0.25** | 0.15 | -0.21* | -0.23* | 0.21* | 0.33** | -0.01 | -0.03 | |||
| 草本密度Shrub density | 0.13 | 0.07 | -0.08 | 0.07 | -0.08 | -0.13 | -0.05 | -0.01 | |||
| 灌木高度Shrub height | 0.17 | 0.00 | -0.51** | -0.37** | 0.06 | 0.12 | -0.05 | -0.06 | |||
| 灌木冠幅Shrubs crown width | 0.16 | 0.07 | 0.07 | 0.09 | -0.05 | 0.06 | 0.08 | 0.11 | |||
| IN:个体数Individual number;GR:类群数Group richness; *表示P < 0.05, * *表示P < 0.01, * * *表示P < 0.001 | |||||||||||
捕食性类群数与土壤pH呈显著正相关(P < 0.05), 与含水量和电导率呈显著负相关(P < 0.05)。植食性类群数与土壤电导率、有机碳、全氮、全磷、黏粒和粉粒含量呈显著正相关(P < 0.01), 与土壤含水量、pH、全钾、砂粒、草本高度和灌木高度呈显著负相关(P < 0.05)。杂食性类群数与土壤pH和草本高度呈显著正相关(P < 0.01), 与土壤含水量呈显著负相关(P < 0.05)。其他食性类群数与个体数和环境因子间的显著相关性保持一致。
4 讨论 4.1 灌丛排列方式对地表节肢动物群落组成的影响土壤动物群落组成是土壤动物生态学的基础[34], 对群落的形成演替、种群特性、物种更新规律的阐明具有重要作用, 可为揭示群落物种共存规律及维持机制提供重要信息[35]。本研究发现, 5、10月份, 带状排列灌丛中地表节肢动物常见类群的个体数和类群数显著高于点状, 7、10月份, 带状排列灌丛中稀有类群的个体数和类群数亦显著高于点状, 这是因为带状排列灌丛中土壤有机碳、全氮和全磷含量较高, 能够为地表节肢动物提供食物资源条件;其次带状排列灌丛中土壤黏粒和粉粒含量较高, 可将植物种子埋存起来[36], 为地表节肢动物提供充足的食物和适宜的栖息地, 提高节肢动物的数量和多样性[37];另外, 带状样地草本丰富度和草本密度显著高于点状, 为不耐高温的地表节肢动物提供了适宜的栖息地[38](表 1, 表 8)。
3个月份中, 2种排列灌丛的优势类群均包含蚁科, 说明蚁科在荒漠草原区人工灌丛林中具有较大优势, 与杨敏等[28]研究结果类似。由于不同灌丛排列方式土壤理化性质存在差异, 地表节肢动物优势类群组成也不尽相同。拟步甲科是干旱荒漠生态系统的重要组分之一, 在干旱荒漠生态系统物质分解和以碳氮等元素为核心的生物地球化学循环过程中发挥着关键作用[39—40]。带状排列灌丛中拟步甲科个体数显著低于点状, 这是因为拟步甲科主要分布于沙丘生境[41], 土壤质地通过直接影响拟步甲科对产卵地点和栖息生境的选择, 从而对它们的空间分布格局产生深刻影响[42], 而点状排列灌丛中土壤砂粒含量显著高于带状, 为拟步甲科提供了良好的生存居住环境;另外, 拟步甲科适应干旱少雨的环境[43], 本研究发现拟步甲科与土壤含水量呈显著负相关(表 8), 与刘继亮等[44]的研究保持一致。步甲科世代周期短, 能快速响应环境变化, 是较好的生态指示类群[45]。总体上看, 带状排列灌丛中步甲科个体数与点状无显著差异;从不同月份来看, 在点状排列灌丛中7和10月份优势类群均包含步甲科, 在带状排列灌丛中仅有10月份包含步甲科, 这是因为带状草本高度在10月份显著低于点状, 而步甲科与草本高度呈显著正相关(表 1, 表 8)。
4.2 灌丛排列方式对地表节肢动物群落多样性的影响土壤环境为地表节肢动物提供了栖息地, 地表植被为其提供了食物来源, 土壤动物类群数量的多少和组成的变化及其多样性的大小通常取决于土壤环境条件的优劣和食物资源的有效性[46], 不同灌丛排列方式对地表节肢动物多样性产生了深刻影响。
除了5、7月份地表节肢动物个体数在2种排列灌丛间无显著差异外, 10月份地表节肢动物个体数和5、7、10月份地表节肢动物类群数均表现为带状显著高于点状(图 3), 这是因为带状排列灌丛中土壤有机碳、粉粒含量和草本丰富度较高(表 1), 为不耐高温的地表节肢动物提供了充足的食物来源[37]和适宜的栖息地[38]。地表节肢动物类群分布与土壤养分和地表植被状况有关[47]。土壤养分影响地表植被生长状况, 如土壤磷和钾含量影响植物的初级生产力和对氮元素的吸收[48—49], 而植物作为地表节肢动物的食物来源[50], 其生长状况影响地表节肢动物类群分布。本研究发现, 地表节肢动物个体数和类群数与土壤全磷呈显著正相关, 而与土壤全钾呈显著负相关(表 9)。地表节肢动物类群数越多, 食物网结构越复杂, 越有利于土壤生态系统的物质循环和能量流动[51], 本研究结果表明灌丛带状排列相较于点状排列更适宜地表节肢动物的生存和定居。
多样性指数的高低可以体现生态系统食物链长短和共生现象的多少[52], 本研究发现, 5、10月份, 带状Shannon-Wienner多样性指数显著高于点状(图 3), 这说明带状排列灌丛的群落结构较点状更稳定。优势度指数是在群落类群组成基础上进一步推算出来以表示群落组成状况的指标, 优势度指数高就表明群落内不同类群的分布越不均匀, 优势类群就更能发挥自身的生态功能[53], 本研究中, 7、10月份Simpson优势度指数在2种灌丛排列间无显著差异, 而5月份表现为带状显著低于点状(图 3), 说明带状排列灌丛物种分布较点状更为均匀。土壤含水量在一定程度上会影响地表节肢动物的分布, 相关性分析发现Shannon-Wienner多样性指数与土壤含水量显著负相关, Simpson优势度指数则相反(表 9)。
4.3 灌丛排列方式对地表节肢动物群落功能群结构的影响地表节肢动物种类繁多, 个体大小、活动能力、活动方式各不相同[54]。各功能类群个体数量是反映环境条件的重要指标[55]。如杂食性个体数可以反映土壤环境限制因子变化幅度, 腐食性个体数可以反映土壤动物对环境的适应程度[56]。不同食性地表节肢动物对灌丛排列方式的敏感程度不同, 导致土壤动物功能群结构在2种排列灌丛间存在差异。
5、7月份, 捕食性个体数表现为点状显著高于带状, 相关性分析发现, 土壤电导率、质地和草本高度对其有显著影响(图 4, 表 10)。5、7月份, 植食性个体数表现为带状显著高于点状, 相关性分析发现, 草本高度和灌木高度与植食性个体数显著相关, 可能是因为土壤有机碳、土壤养分和土壤质地通过影响植被生长状况, 进而对植食性个体数造成影响(图 4, 表 10)。10月份, 带状排列灌丛杂食性个体数显著高于点状, 这可能与优势类群蚁科相关, 通过相关性分析发现蚁科和杂食性个体数均与土壤含水量呈显著负相关(图 4, 表 8, 表 10)。捕食性与植食性之间的数量比值, 反映了地表节肢动物食物网结构中的定量营养级关系[57]。本研究发现, 带状排列灌丛中植食性类群的数量比重高于捕食性类群, 在食物网结构中呈“金字塔”型, 更有利于维持地表节肢动物营养级关系及食物网结构的相对稳定性[58]。
功能群划分反映了不同节肢动物类群间营养级联关系[59], 本研究中, 带状排列灌丛中植食性类群表现出更大的优势地位, 点状排列灌丛中捕食性类群表现出更大的优势地位, 较杂食性和其他食性而言, 捕食性和植食性类群的占比较大(图 5), 反映了研究区域地表节肢动物区系以捕食性和植食性类群分布为主要特征。5、7、10月份植食性类群数和5、10月份杂食性类群数均表现为带状显著高于点状(图 5), 这是由于有些物种仅在带状出现, 如植食性蜢总科和鳃金龟科等;杂食性蚁形甲科和蟋蟀总科等。这反映了这些地表节肢动物类群对生境选择的严格性, 对环境变化起着重要的生物指示作用[10]。
5 结论带状排列灌丛中优势类群包括蚁科和步甲科, 点状排列灌丛中优势类群包括蚁科、拟步甲科和步甲科。地表节肢动物个体数在10月份表现为带状显著高于点状;类群数在5、7、10月份表现为带状显著高于点状;Shannon-Wienner多样性指数在5、10月份表现为带状显著高于点状;而Simpson优势度指数在5月份表现为带状显著低于点状。捕食性个体数在5、7月份表现为带状显著低于点状;而植食性个体数在5、7月份表现为带状显著高于点状;杂食性个体数在10月份表现为带状显著高于点状;其他食性个体数在5月份表现为带状显著高于点状。捕食性类群数在5月份表现为带状显著低于点状;而植食性类群数在5、7、10月份表现为带状显著高于点状;杂食性类群数在5、10月份表现为带状显著高于点状, 但杂食性类群数在7月份表现为带状显著低于点状;其他食性类群数在5月份表现为带状显著高于点状。影响地表节肢动物分布的主要环境因子包括土壤容重、pH和草本丰富度。综上, 带状排列灌丛更适合地表节肢动物生存和定居, 更利于维持地表节肢动物多样性。因此, 在未来荒漠生态系统种植固沙灌丛进行生态恢复时, 应根据灌丛在不同排列方式下的功能特征, 制定因地制宜的恢复方法, 关注生态恢复过程中土壤生物多样性的动态变化, 以促进荒漠生态系统长期稳定持续发展。
| [1] |
Schlesinger W H, Reynolds J F, Cunningham G L, Huenneke L F, Jarrell W M, Virginia R A, Whitford W G. Biological feedbacks in global desertification. Science, 1990, 247(4946): 1043-1048. DOI:10.1126/science.247.4946.1043 |
| [2] |
于钊, 李奇铮, 王培源, 蒋齐. 退化和恢复过程驱动的荒漠草地生态系统有机碳密度变化. 中国沙漠, 2022, 42(2): 215-222. |
| [3] |
赛希雅拉, 阿拉塔, 孙海莲. 浅谈华北驼绒藜灌木防护带建植模式及防护效益. 草原与草业, 2016, 28(1): 28-31. DOI:10.3969/j.issn.2095-5952.2016.01.008 |
| [4] |
王炜炜. 盐池县沙地灌丛人工植被配置对土壤风蚀影响机理研究[D]. 北京: 北京林业大学, 2008.
|
| [5] |
杨文斌, 赵爱国, 王晶莹, 姚建成, 田永桢, 胡小龙, 杨红艳. 低覆盖度沙蒿群丛的水平配置结构与防风固沙效果研究. 中国沙漠, 2006, 26(1): 108-112. DOI:10.3321/j.issn:1000-694X.2006.01.019 |
| [6] |
杜丽侠, 邢韶华. 北京八达岭地区灌木群落空间分布格局与土壤环境因子的关系分析. 西北植物学报, 2009, 29(3): 601-607. DOI:10.3321/j.issn:1000-4025.2009.03.027 |
| [7] |
马全林, 张德奎, 袁宏波, 郑庆钟, 丁峰, 张锦春, 魏怀东, 靳虎甲, 刘有军, 孙涛, 王继和. 乌兰布和沙漠植被数量分类及环境解释. 干旱区资源与环境, 2019, 33(9): 160-167. |
| [8] |
李锦荣, 崔崴, 王茹, 董雷, 张志杰, 张晓燕. 草原荒漠区红砂和珍珠猪毛菜群落分布特征分析. 中国水利水电科学研究院学报: 中英文, 2024, 22(2): 179-194. |
| [9] |
刘继亮, 赵文智, 李锋瑞, 巴义彬. 天然和人工固沙灌木林蜘蛛和甲虫分布与环境因子的关系. 生态学报, 2020, 40(21): 7987-7996. DOI:10.5846/stxb201911292590 |
| [10] |
张安宁, 常海涛, 陈蔚, 刘任涛. 干旱风沙区灌丛林地地面节肢动物群落对放牧管理的响应. 应用生态学报, 2019, 30(11): 3931-3941. |
| [11] |
冯永宏, 刘任涛, 刘纪贤, 蒋嘉瑜, 白燕娇, 郭志霞, 王文帆, 张安宁. 荒漠区油蒿(Artemisia ordosica)灌丛冠层中节肢动物群落结构特征. 中国沙漠, 2021, 41(5): 94-102. |
| [12] |
刘继亮, 赵文智, 李锋瑞. 黑河中游荒漠地面节肢动物分布特征. 干旱区研究, 2015, 32(6): 1192-1200. |
| [13] |
Piñero F S, Tinaut A, Aguirre-Segura A, Miñano J, Lencina J L, Ortiz-Sánchez F J, Pérez-López F J. Terrestrial arthropod fauna of arid areas of SE Spain: diversity, biogeography, and conservation. Journal of Arid Environments, 2011, 75(12): 1321-1332. DOI:10.1016/j.jaridenv.2011.06.014 |
| [14] |
McIntyre N E, Rango J, Fagan W F, Faeth S H. Ground arthropod community structure in a heterogeneous urban environment. Landscape and Urban Planning, 2001, 52(4): 257-274. DOI:10.1016/S0169-2046(00)00122-5 |
| [15] |
da Silva Santana M, Andrade E M, Oliveira V R, Costa B B, Silva V C, do Socorro Conceição de Freitas M, Cunha T J F, Giongo V. Trophic groups of soil fauna in semiarid: impacts of land use change, climatic seasonality and environmental variables. Pedobiologia, 2021, 89: 150774. DOI:10.1016/j.pedobi.2021.150774 |
| [16] |
Bestion E, Cote J, Jacob S, Winandy L, Legrand D. Habitat fragmentation experiments on arthropods: what to do next?. Current Opinion in Insect Science, 2019, 35: 117-122. DOI:10.1016/j.cois.2019.07.011 |
| [17] |
Li F R, Liu J L, Sun T S, Ma L F, Liu L L, Yang K. Impact of established shrub shelterbelts around oases on the diversity of ground beetles in arid ecosystems of NorthWestern China. Insect Conservation and Diversity, 2016, 9(2): 135-148. DOI:10.1111/icad.12152 |
| [18] |
Megías A G, Sánchez-Piñero F, Hódar J A. Trophic interactions in an arid ecosystem: from decomposers to top-predators. Journal of Arid Environments, 2011, 75(12): 1333-1341. DOI:10.1016/j.jaridenv.2011.01.010 |
| [19] |
娄巧哲, 徐养诚, 马吉宏, 吕昭智. 古尔班通古特沙漠南缘地表甲虫物种多样性及其与环境的关系. 生物多样性, 2011, 19(4): 441-452. |
| [20] |
刘志涛, 王莹, 高铸成, 丁敏, 叶董, 王京, 王瑞, 林英华. 环境因子对甘南高寒草甸地表大型节肢动物多样性的影响. 动物学杂志, 2023, 58(2): 237-249. |
| [21] |
刘任涛. 沙地灌丛的"肥岛" 和"虫岛" 形成过程、特征及其与生态系统演替的关系. 生态学杂志, 2014, 33(12): 3463-3469. |
| [22] |
常海涛, 刘佳楠, 赵娟, 张静, 罗雅曦, 刘任涛, 左小安. 乌拉特荒漠草原红砂(Reaumuria soongorica)灌丛林地地面节肢动物群落季节分布特征. 生态学报, 2020, 40(9): 3121-3130. DOI:10.5846/stxb201904150756 |
| [23] |
刘任涛, 张安宁. 固沙灌丛林营造初期地面节肢动物群落结构特征. 中国沙漠, 2020, 40(5): 190-199. |
| [24] |
Doblas-Miranda E, Sánchez-Piñero F, González-Megías A. Soil macroinvertebrate fauna of a Mediterranean arid system: composition and temporal changes in the assemblage. Soil Biology and Biochemistry, 2007, 39(8): 1916-1925. DOI:10.1016/j.soilbio.2007.02.009 |
| [25] |
刘任涛, 朱凡, 柴永青. 干旱区不同年龄灌丛斑块地面节肢动物的聚集效应. 应用生态学报, 2014, 25(1): 228-236. |
| [26] |
刘晓丽, 丁训欢, 宁杰, 张旭, 孙涛. 皖北石灰岩山地植被对地表节肢动物组成及多样性的影响. 生态学报, 2021, 41(15): 6115-6122. DOI:10.5846/stxb202001150117 |
| [27] |
常海涛, 刘任涛, 陈蔚, 张安宁, 左小安. 内蒙古乌拉特荒漠草原红砂灌丛林引入柠条后地面节肢动物群落结构分布特征. 草业学报, 2020, 29(12): 188-197. DOI:10.11686/cyxb2020019 |
| [28] |
杨敏, 刘任涛, 曾飞越, 吉雪茹, 方进, 赵文智. 腾格里沙漠东南缘人工固沙植被演替地面节肢动物群落多样性分布特征. 生态学报, 2024, 44(1): 428-439. DOI:10.20103/j.stxb.202302130244 |
| [29] |
尹文英. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998.
|
| [30] |
王新谱, 杨贵军. 宁夏贺兰山昆虫. 银川: 宁夏人民出版社, 2010.
|
| [31] |
刘任涛, 郗伟华, 刘佳楠, 赵娟, 常海涛. 沙地柠条(Caragana)灌丛微生境节肢动物群落特征. 中国沙漠, 2018, 38(1): 117-125. |
| [32] |
冯腾, 陈洪松, 张伟, 王克林. 激光粒度仪与沉降吸管法测定喀斯特地区土壤机械组成的对比研究. 农业现代化研究, 2013, 34(1): 100-103. |
| [33] |
鲍士旦. 土壤农化分析 (3版). 北京: 中国农业出版社, 2000.
|
| [34] |
尹文英. 土壤动物学研究的回顾与展望. 生物学通报, 2001, 36(8): 1-3. DOI:10.3969/j.issn.0006-3193.2001.08.001 |
| [35] |
Loreau M, Naeem S, Inchausti P, Bengtsson J, Grime J P, Hector A, Hooper D U, Huston M A, Raffaelli D, Schmid B, Tilman D, Wardle D A. Biodiversity and ecosystem functioning: current knowledge and future challenges. Science, 2001, 294(5543): 804-808. DOI:10.1126/science.1064088 |
| [36] |
李生宇, 雷加强. 草方格沙障的生态恢复作用——以古尔班通古特沙漠油田公路扰动带为例. 干旱区研究, 2003, 20(1): 7-10. |
| [37] |
王永珍, 林永一, 冯怡琳, 赵文智, 董六文, 刘继亮. 沙障对流动沙丘区地表节肢动物分布及多样性的影响. 生态学报, 2022, 42(16): 6768-6777. DOI:10.5846/stxb202108182294 |
| [38] |
Shelef O, Groner E. Linking landscape and species: Effect of shrubs on patch preference of beetles in arid and semi-arid ecosystems. Journal of Arid Environments, 2011, 75(10): 960-967. DOI:10.1016/j.jaridenv.2011.04.016 |
| [39] |
Shachak M, Pickett S T A, Gosz J R, Perevolotski A. Biodiversity in drylands: toward a unified framework. New York: Oxford University Press, 2005.
|
| [40] |
Whitford W G. The importance of the biodiversity of soil biota in arid ecosystems. Biodiversity & Conservation, 1996, 5(2): 185-195. |
| [41] |
董六文, 韩佳龙, 赵文智, 刘继亮, 巴义彬. 黑河流域湖泊湿地及毗邻沙丘地表节肢动物群落结构比较. 中国沙漠, 2020, 40(6): 250-258. |
| [42] |
Stapp P. Microhabitat use and community structure of darkling beetles (Coleoptera: Tenebrionidae) in shortgrass prairie: effects of season shrub and soil type. American Midland Naturalist, 1997, 137(2): 298. DOI:10.2307/2426849 |
| [43] |
Thomas D. Tenebrionid beetle diversity and habitat complexity in the eastern mojave desert. The Coleopterists Bulletin, 1983, 37(2): 135-147. |
| [44] |
刘继亮, 李锋瑞, 刘七军, 牛瑞雪. 黑河中游干旱荒漠地面节肢动物群落季节变异规律. 草业学报, 2010, 19(5): 161-169. |
| [45] |
Rainio J, Niemelä J. Ground beetles (Coleoptera: Carabidae) as bioindicators. Biodiversity & Conservation, 2003, 12(3): 487-506. |
| [46] |
赵哈林, 刘任涛, 周瑞莲, 赵学勇, 张铜会. 科尔沁沙地灌丛的"虫岛" 效应及其形成机理. 生态学杂志, 2012, 31(12): 2990-2995. |
| [47] |
林英华, 宋百敏, 韩茜, 王静, 张永, 于长青. 北京门头沟废弃采石矿区地表土壤动物群落多样性. 生态学报, 2007, 27(11): 4832-4839. DOI:10.3321/j.issn:1000-0933.2007.11.053 |
| [48] |
Sundquist E, Broecker W. The carbon cycle and atmospheric CO2: natural variations Archean to present. Washington: American Geophysical Union, 1985.
|
| [49] |
金月, 梁存柱, 崔利剑. 不同养分添加对草地群落优势植物叶片化学计量特征的影响. 北方农业学报, 2019, 47(2): 59-65. DOI:10.3969/j.issn.2096-1197.2019.02.10 |
| [50] |
孙彩彩, 董全民, 刘文亭, 冯斌, 时光, 刘玉祯, 俞旸, 张春平, 张小芳, 李彩弟, 杨增增, 杨晓霞. 放牧方式对青藏高原高寒草地土壤节肢动物群落结构和多样性的影响. 草业学报, 2022, 31(2): 62-75. |
| [51] |
刘任涛, 杨新国, 柴永青, 杨明秀, 朱凡, 李淑君. 荒漠草原区柠条林地地面节肢动物功能群对补播牧草和平茬措施的响应. 草业学报, 2013, 22(3): 78-84. |
| [52] |
刘新民, 门丽娜. 内蒙古武川县农田退耕还草对大型土壤动物群落的影响. 应用生态学报, 2009, 20(8): 1965-1972. |
| [53] |
崔丽巍, 刘世荣, 刘兴良, 吴鹏飞, 张洪芝, 何先进. 川西北冷杉林恢复过程中土壤动物群落动态. 生态学报, 2012, 32(15): 4772-4783. DOI:10.5846/stxb201111171746 |
| [54] |
刘洁, 高梅香, 吴东辉. 基于黑土农田生物多样性监测样地的地表大型节肢动物群落特征. 应用生态学报, 2017, 28(12): 3965-3975. |
| [55] |
Cole L, Bradford M A, Shaw P J A, Bardgett R D. The abundance, richness and functional role of soil meso- and macrofauna in temperate grassland—A case study. Applied Soil Ecology, 2006, 33(2): 186-198. DOI:10.1016/j.apsoil.2005.11.003 |
| [56] |
Brussaard L. Soil fauna, guilds, functional groups and ecosystem processes. Applied Soil Ecology, 1998, 9(1/2/3): 123-135. |
| [57] |
Zhao Z H, Shi P J, Men X Y, Ouyang F, Ge F. Effects of crop species richness on pest-natural enemy systems based on an experimental model system using a microlandscape. Science China Life Sciences, 2013, 56(8): 758-766. DOI:10.1007/s11427-013-4511-3 |
| [58] |
Li F R, Liu J L, Sun T S, Jin B W, Chen L J. Converting natural vegetation to farmland alters functional structure of ground-dwelling beetles and spiders in a desert oasis. Journal of Insect Conservation, 2014, 18(1): 57-67. DOI:10.1007/s10841-014-9614-z |
| [59] |
程建伟, 刘新民, 郝百惠, 张芯毓, 张宇平, 马文红. 氮沉降对内蒙古典型草原地表节肢动物的影响. 生态学杂志, 2017, 36(8): 2237-2245. |