 2025, Vol. 45
2025, Vol. 45文章信息
- 吴奇, 问青青, 庄前进, 陈建华, 傅佳琴, 谢家杰, 施怡珊, 张雨宁, 曾子君, 符蝶, 王云泉
- WU Qi, WEN Qingqing, ZHUANG Qianjin, CHEN Jianhua, FU Jiaqin, XIE Jiajie, SHI Yishan, ZHANG Yuning, ZENG Zijun, FU Die, WANG Yunquan
- 浙江婺城南山落叶阔叶林树冠互补性对群落多样性的影响
- Effects of crown complementarity on community diversity in the Nanshan deciduous broadleaf forest, Zhejiang Province, China
- 生态学报. 2025, 45(7): 3330-3340
- Acta Ecologica Sinica. 2025, 45(7): 3330-3340
- http://dx.doi.org/10.20103/j.stxb.202403120504
-
文章历史
- 收稿日期: 2024-03-12
- 网络出版日期: 2025-01-02






2. 金华婺城南山省级自然保护区管理中心, 金华 321004;
3. 浙江大学生命科学学院, 杭州 310058;
4. 浙江九龙山国家级自然保护区管理中心, 丽水 323300
2. The Administration Center of Jinhua Wucheng Nanshan Provincial Nature Reserve, Jinhua 321004, China;
3. College of Life Sciences, Zhejiang University, Hangzhou 310058, China;
4. The Administration Center of Zhejiang Jiulongshan National Nature Reserve, Lishui 323300, China
森林群落的生物多样性分布格局及其维持机制一直是群落生态学研究的核心科学问题, 其中的关键是理解群落中的物种如何实现稳定共存[1—3], 这对揭示森林生物多样性对全球气候变化的响应具有十分重要的意义[4—6]。生态位分化和生境过滤是决定群落内物种能否实现稳定共存和生物多样性维持的关键生态过程, 生态位分化通常被认为是减弱或避免局域尺度上物种的竞争排除从而实现群落内物种稳定共存的必要条件[2, 7—9]。鉴于森林群落物种组成的复杂性和生境异质性, 不同物种往往通过采取生存空间和资源的互补利用策略以占据不同的生态位[10—11]。例如, 密度制约效应使得同种个体之间存在更高的竞争强度和病虫害风险, 从而降低该物种的种间竞争优势, 为森林群落中其他物种提供生存空间[12—13];稀有树种在环境梯度上往往具有较窄的生态位宽度并处于边缘的生态位位置, 避免与优势树种竞争相似的资源[14];外生菌根和内生菌根树种在土壤养分获取方面具有不同的机制, 采用混合菌根策略的森林群落往往具有较高的生态位分化水平[15]。虽然生态学家意识到森林群落高度复杂的冠层结构对群落生物多样性维持至关重要[16—18], 但目前少有研究直接评估群落中树冠结构差异导致的树冠互补性对森林群落生物多样性的贡献。
森林群落内错落排布的树冠结构显著改变了群落的物种组成、群落结构乃至群落生产力[19—20]。可塑性的树冠构型对光辐射的拦截作用导致森林群落中光环境异质性的形成, 而森林光环境的分布特征对于树木的生长动态及空间分布格局起到决定性的作用[21—22]。因此, 森林群落共存物种之间树木冠层的分层现象促使不同个体沿着光可利用性的垂直梯度形成生态位分化, 从而显著提高了群落整体对光照的截获能力和对光能的利用效率[23], 并影响群落内的生物多样性[24]。换言之, 森林群落中共存物种间树冠结构的差异(即树冠互补性)反映了不同物种对冠层空间的互补利用, 从而减弱或避免群落中共存物种之间的竞争排除, 有利于群落生物多样性的维持, 可以作为衡量森林群落空间生态位分化程度的一个重要指标[25—26]。
除了树冠结构差异等生物因素导致的生态位分化对森林群落物种共存的影响外, 气候条件和土壤养分等非生物因素在生物多样性维持中也具有关键作用[27—29]。地形因子通过调节土壤养分、水分及太阳辐射等环境因子的空间分布, 成为主导群落内部生境异质性和植被格局的重要因素之一[30—31]。具体而言, 地形因子造成的生境过滤作用从区域物种库中筛选出能够适应特定环境的物种, 最终导致具有相似生境需求和功能性状的近缘种得以共存, 从而影响森林群落的物种组成、谱系结构和林分结构的形成[32—34]。此外, 地形因子还可以通过驱动森林冠层结构的精细变化影响林分小气候, 与森林冠层结构共同参与森林群落多样性的维持过程[35]。
综上所述, 森林群落中树冠在垂直空间的互补性可能导致空间生态位的分化, 并且与地形因子导致的生境过滤作用共同影响森林群落多样性的维持。同时监测森林群落物种树冠结构和地形变量的生物多样性长期监测样地为评估树冠互补性和环境变量对森林群落生物多样性的贡献提供了理想的研究平台。因此, 为了评估树冠互补性对群落多样性的影响, 本研究利用2023年浙江婺城南山落叶阔叶林1 hm2动态监测样地数据, 具体分析了树冠互补性和物种多样性、谱系多样性及结构多样性等群落多样性的关系, 主要探讨以下科学问题:(1)树冠互补性对森林群落多样性的影响;(2)地形因子对森林群落多样性的影响;(3)树冠互补性和地形因子对森林群落多样性的相对影响。
1 研究方法 1.1 研究地概况浙江婺城南山省级自然保护区(简称“南山保护区”)位于浙江金华西南山区白沙溪源头, 总面积9532.60hm2, 为仙霞岭山脉东北段, 属武夷山生物多样性保护优先区域, 是浙中地区南部生态屏障的重要组成部分, 也是浙中中心城市的重要水源地。南山保护区是浙江省面积最大的省级自然保护区, 是保护中亚热带中高海拔落叶阔叶林及穿山甲、黄腹角雉和永瓣藤等珍稀濒危野生动植物的森林生态系统类型自然保护区, 具有丰富的生物资源。
南山保护区的气候属于亚热带季风区, 四季分明, 雨量充沛。该区域年均温17℃, 最热月均温27.4℃, 最冷月均温4.2℃, 年均降雨量1449.8 mm, 土壤类型以黄壤和红壤为主。南山保护区主要植被类型有阔叶林、针叶阔叶混交林、针叶林、竹林和草丛等。其中, 中高海拔分布着大面积的落叶阔叶林和针阔混交林, 为保护区植被类型多样性的主要贡献者, 是南山保护区的主要保护对象。保护区内动植物资源丰富, 是浙中地区生物多样性最富集的地区之一。保护区内共有维管植物171科710属1495种, 珍稀濒危植物89种, 其中国家一级重点保护野生植物2种, 国家二级重点保护野生植物11种, 浙江省重点保护区野生植物18种(婺城南山省级自然保护区综合科学考察报告, 2019)。
1.2 样地设置与调查方法婺城南山落叶阔叶林1 hm2固定监测样地(简称“南山样地”)位于婺城南山省级自然保护区大西坑区域(图 1;119°24′30.6″E, 28°46′3.7″N), 于2023年完成样地建设和首次调查。样地东西水平距离100 m, 南北水平距离100 m, 最低海拔1012 m, 最高海拔1103 m, 总体呈东南高、西北低的趋势。南山样地建设参照全球森林生物多样性监测网络(Forest Global Earth Observatory Network, ForestGEO)和中国森林生物多样性监测网络(Chinese Forest Biodiversity Monitoring Network, CForBio)规范要求[5, 36], 将整个样地划分为25个20 m×20 m的样方, 再用样绳将每个20 m×20 m的样方划分成16个5 m×5 m的小样方。样地调查时以5 m×5 m的小样方为基本测树单元, 对样地内所有胸径≥1 cm的木本植物在离地面1.3 m处刷红色油漆标识并进行每木检尺, 调查记录内容包括立木序号、物种名、胸径、树高、冠幅、枝下高、坐标和立木状态等信息。由于林冠层截获了森林冠层空间中的大部分光资源[37], 本研究选取调查数据中2637株胸径≥5 cm的木本植物作为研究对象。

|
| 图 1 婺城南山1 hm2固定样地位置示意图 Fig. 1 Location of the Wucheng Nanshan 1 hm2 plot |
本研究利用样地调查期间由测高杆测定的树高、枝下高和冠幅数据, 通过将针叶和阔叶树种的树冠近似处理为圆锥形和椭球形的方法估测树冠体积, 计算公式如下所示[25]。
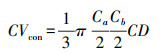
|
(1) |
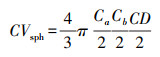
|
(2) |
式中, CVcon、CVsph分别表示圆锥形树冠和椭球形树冠的体积, Ca、Cb分别表示东西方向和南北方向的冠幅, CD表示树冠长度, 即树高与枝下高的差值。
为衡量森林群落中的树冠垂直分层和树木个体间的树冠尺寸分化情况, 本研究利用每对树木之间树冠体积差异的算数平均值衡量样方水平的树冠互补性指数(Crown complementarity index, CCI), 计算公式如下所示[26]:
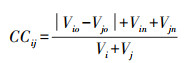
|
(3) |
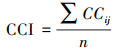
|
(4) |
式中, CCij表示树i和树j间的树冠互补性, Vio、Vjo分别表示树i和树j的树冠在垂直方向上的重叠体积, Vin、Vjn分别表示树i和树j的树冠在垂直方向上的非重叠体积, Vi、Vj分别表示树i和树j的树冠体积, n表示样方内的物种对数。
1.4 群落多样性计算 1.4.1 物种多样性计算本研究利用vegan软件包计算Shannon-Wiener指数(H′)和Pielou均匀度指数(J), 计算公式如下所示[38]:

|
(5) |

|
(6) |
式中, pi为物种i的多度, S为样方内的物种数。
1.4.2 谱系多样性计算本研究所采用的谱系树包含了所有现存的种子植物科和一些蕨类植物的科, 使用基于The Plant List (TPL;http://www.theplantlist.org)植物命名法的V.PhyloMaker2软件包构建[39—41]。利用picante软件包基于群落谱系树计算净亲缘关系指数(Net relatedness index, NRI)和最近亲缘关系指数(Nearest taxa index, NTI), 计算公式如下所示[42—43]:

|
(7) |
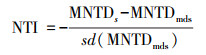
|
(8) |
式中, sd指标准差, MPD和MNPD分别表示物种间的平均成对谱系距离与平均最近种间谱系距离, MPDs和MNTDs表示实际观测值, MPDmds和MNTDmds表示随机模拟零模型的期望值[42]。
1.4.3 结构多样性计算本研究采用树高变异系数(Coefficient of variation of height)和胸径变异系数(Coefficient of variation of DBH)测度结构多样性, 计算公式如下所示[44]:
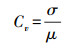
|
(9) |
式中, Cv指树高或胸径的变异系数, σ指树高或胸径的标准差, μ指树高或胸径的均值。
1.5 地形因子计算本研究选取了海拔、坡度、坡向和凹凸度4类地形因子。海拔指各样方中心点的相对高度, 坡度指各样方坡面与水平面所成的夹角, 坡向指各样方坡面与正北方向所成的顺时针方向的夹角, 凹凸度指目标样方的海拔与相邻样方的平均海拔之差[45]。
1.6 统计分析本研究所有数据分析和绘图均于R语言(版本4.2.1)中进行。为保证所有变量均通过Shapiro-Wilk正态性检验(P > 0.05), 对呈右偏分布的胸径变异系数进行了对数转换(处理前偏度=1.37)。使用线性或非线性回归模型分析树冠互补性对群落多样性的影响, 根据单项式或多项式回归模型的拟合度(R2)选择回归方式, 并采用Pearson相关性分析检验地形因子和群落多样性的相关性。构建多元回归模型分析树冠互补性和地形因子对群落多样性的相对影响, 将树冠互补性指数、树冠互补性指数的二次项与4种地形因子同时纳入全模型[46]。利用MuMIn包进行全子集回归, 根据小样本赤池信息准则(Akaike information criterion for small samples, AICc)筛选出同时含有树冠互补性和地形因子两类变量的最优模型。若不同模型间的ΔAICc≤2, 则选择变量最少的模型作为最优模型[47]。模型拟合过程中利用方差膨胀因子(Variance inflation factor, VIF)检查变量间的多重共线性, 保证所保留变量的VIF均<5[48]。通过计算某一变量的参数估计值与所有变量的参数估计值之和的比例, 评估树冠互补性和地形因子的相对贡献率[49]。
2 结果与分析 2.1 树冠互补性对群落多样性的影响随树冠互补性指数的增大, Shannon-Wiener指数、Pielou指数和树高变异系数显著增大(图 2), 但胸径变异系数无显著变化(图 2)。谱系结构和树冠互补性指数的关系较为复杂, 净谱系亲缘关系指数(NRI)与树冠互补性指数成反比(图 2), 但最近种间亲缘关系指数(NTI)随树冠互补性指数的增大表现出U型趋势(图 2)。就模型拟合效果而言, 树冠互补性指数与树高变异系数的拟合度最高(图 2), 与NRI的拟合度次之(图 2)。

|
| 图 2 树冠互补性与群落多样性间的双变量关系 Fig. 2 Bivariate relationships between crown complementarity and community diversity 图中灰色阴影代表 95%置信区间; 蓝色、红色和绿色部分分别表示物种多样性、谱系多样性和结构多样性与树冠互补性指数的关系; *P < 0.05, * * P < 0.01, * * * P < 0.001 |
海拔与Shannon-Wiener指数、Pielou指数、树高变异系数、胸径变异系数极显著负相关(图 3), 与NRI和NTI显著正相关。凹凸度与Shannon-Wiener指数、Pielou指数、树高变异系数、胸径变异系数显著负相关, 与NRI极显著正相关, 但与NTI不表现出相关性(图 3)。坡向和坡度与群落多样性大多不表现出相关性, 仅坡向与胸径变异系数呈显著正相关(图 3)。

|
| 图 3 地形因子与群落多样性的相关性 Fig. 3 Correlation between topographic factors and community diversity *P < 0.05, * * P < 0.01, * * * P < 0.001 |
树冠互补性对NRI、NTI和树高变异指数的相对贡献率与地形因子相近, 分别占可解释方差的50.16%、49.84%和53.60%(图 4), 但仅对NRI(P=0.0452, 图 4)和树高变异系数(P=0.0358, 图 4)造成显著负向和正向影响。地形因子对Shannon-Wiener指数、Pielou均匀度指数和胸径变异系数的相对贡献率大于树冠互补性, 分别占可解释方差的56.76%、68.78%和99.70%(图 4)。海拔是在最优模型中出现次数最多的地形因子, 对Shannon-Wiener指数(P=0.0255, 图 4)、Pielou均匀度指数(P=0.0018, 图 4)和胸径变异系数(P < 0.001, 图 4f)造成显著负向影响, 且对NRI造成显著正向影响(P=0.0465, 图 4)。

|
| 图 4 地形因子和树冠互补性对群落多样性的相对影响 Fig. 4 Relative effect of crown complementarity and topographic factors on community diversity CCI:树冠互补性指数;CON:凹凸度;ELE:海拔;ASP:坡向;SLO:坡度; 在NTI的最优模型中, CCI指树冠互补性指数的二次项; 堆叠柱形图的蓝色部分表示树冠互补性的相对贡献率, 红色部分表示地形因子的相对贡献率; *P < 0.05, * *P < 0.01, * * *P < 0.001 |
本研究发现, 树冠互补性对物种多样性和树高分化程度具有正向影响, 但与谱系多样性的关系较为复杂(图 2)。与前人研究结果相类似, 婺城南山落叶阔叶林中树冠互补性与物种多样性也存在明显的正相关关系(图 2), 这是因为树冠互补性的提高优化了森林冠层空间结构的分配方式, 提高了群落中不同植株间空间生态位的利用程度, 进而促进了物种共存和物种多样性的维持[50—51]。不同于物种多样性所提供的群落构建中关于生态过程的信息, 谱系多样性补充了群落构建中关于进化过程的信息[52]。NRI考虑了整个谱系树的平均谱系距离, 而NTI同时纳入了近缘种的谱系距离, 二者分别反映了深层和浅层的谱系结构[53]。本研究发现, 树冠互补性的提高可以使NRI发生明显的下降(图 2), 表明由树冠互补性表征的生态位分化可以使深层的谱系结构趋于发散。该结果与生态位保守理论的假设相符, 即物种在长时间演化过程中的生态位特征保持相对稳定, 且亲缘关系较远的物种间往往存在较低的生态位相似性[54]。然而, NTI与树冠互补性存在非线性的关系(图 2), 暗示了驱动浅层谱系结构维持的生态过程可能较为复杂。NTI在树冠互补性达到一定阈值后呈现出一段异常的上升趋势, 这可能是因为群落浅层的谱系结构响应了林冠结构复杂的生境中特殊的小气候格局[55]。本研究还关注了结构多样性这一相对不受重视的群落多样性维度[56], 发现树冠互补性的提升主要增加了树高而非胸径的变异(图 2)。这是因为树冠结构的差异主要反映森林群落垂直层面上的空间生态位分化, 使得树冠互补性主要影响植株高度的异质性或林分垂直结构的复杂性[57]。此外, 已有研究表明利用胸径衡量的森林结构难以反映异质性的光可利用性格局[58], 与本研究中树冠互补性几乎不影响群落水平结构分化程度的结论相符。总之, 本研究表明树冠互补性可以衡量森林群落中的垂直生态位分化, 在一定程度上促进了森林群落多样性的维持。
3.2 地形因子对婺城南山落叶阔叶林群落多样性的影响海拔被广泛认为是影响群落多样性的重要环境因素, 但针对群落多样性海拔梯度格局的研究并未得出统一的结论[59]。在全球范围内针对植物海拔梯度分布的研究中, 有超过半数的研究检验到了物种多样性的单峰格局, 但也有部分研究发现物种多样性与海拔呈线性相关[60]。在本研究选取的坡度、坡向、海拔和凹凸度4类地形因子中, 海拔和凹凸度与群落多样性表现出较高的相关性(图 3)。在海拔高度为1012 m至1103 m的婺城南山落叶阔叶林中, 物种多样性和结构多样性与海拔呈线性负相关, 谱系结构也随海拔升高而趋于聚集(图 3)。凹凸度也导致了类似的生物多样性格局, 但只能使深层的谱系结构趋于聚集(图 3)。与已有研究类似, 本研究也检测到了深层和浅层的谱系结构对同一环境因子的不同响应模式[52]。此外, 坡向和坡度对群落多样性的影响程度较小, 可能和本样地的不同样方所受的太阳辐射强度基本一致有关[61]。综上所述, 树冠互补性与地形因子主要代表了生态位分化和生境过滤两个不同的生态过程, 共同影响婺城南山落叶阔叶林群落多样性的维持。然而, 根据森林动态监测样地研究的惯例, 本研究仅选取了较易获得的地形因子数据[62]。海拔间接代表了光照和温度等环境因子, 凹凸度间接代表了土壤质地和含水量等土壤因子, 不能完全涵盖生物所处局域地区的小环境[63—64], 因此仍有必要进一步检验其他环境因素导致的生境过滤作用[65]。
3.3 树冠互补性和地形因子对婺城南山落叶阔叶林群落多样性的相对贡献树冠互补性和地形因子对不同维度群落多样性的影响存在一定差异, 二者对Shannon-Wiener指数、NRI、NTI和树高变异系数的相对贡献率相近, 但后者对Pielou均匀度指数和胸径变异系数的相对贡献率明显较高(图 4)。物种丰富度和均匀度是物种多样性的2个关键组成部分, 二者在响应同一生态过程时可能表现出不同的特征[66]。本研究发现, 地形因子对物种多样性的相对贡献率相较树冠互补性更大, 且对Pielou均匀度指数的相对贡献率最为突出(图 4), 表明婺城南山落叶阔叶林中的物种分布格局比物种组成情况所受到的生境过滤作用更为明显。树冠互补性和地形因子对NRI和NTI的相对贡献率基本相近, 以树冠互补性和NRI为变量的线性回归模型也存在较高的拟合度(图 4), 这意味着树冠互补性与谱系结构的构建密切相关, 并可能主要在长期的历史演化过程中影响谱系结构的构建[67]。此外, 树冠互补性和地形因子对不同层面结构多样性维持的相对重要性截然不同(图 4)。由于树高变异系数和胸径变异系数分别表示了森林群落垂直和水平空间的配置情况[68], 因此该结果进一步证实了树冠互补性在反映垂直生态位分化方面的重要作用, 同时也凸显出地形因子对森林群落水平空间配置情况不可忽视的影响。综上所述, 落叶阔叶林群落多样性的维持受多种生态过程的驱动, 由树冠互补性表征的生态位分化与地形因子导致的生境过滤作用在一定程度上相拮抗。值得注意的是, 除了本研究发现的树冠互补性在森林群落生物多样性维持方面的作用外, 也有研究表明树冠互补性可以驱动物种多样性和森林群落生产力之间的关系[25—26]。综合考虑树冠互补性作为生态位分化程度或者生态位互补性的度量方式, 不仅将为整合生物多样性维持机制和生物多样性效应的理论框架贡献一个可能的思路, 还有望为减缓气候变化负面影响的森林管理和保育过程提供基于自然的解决方案(Nature-based solution)[69—70]。
4 结论理解森林群落树冠空间结构的互补性对群落多样性的贡献有利于进一步加深对生物多样性维持机制的认识。本研究利用婺城南山落叶阔叶林群落调查数据和树木冠幅信息, 分析了树冠互补性和地形因子对婺城南山落叶阔叶林群落多样性的影响并评估了二者的相对贡献。本研究发现树冠互补性的增加有利于提高群落内物种多样性和树高变异程度, 同时导致深层谱系结构趋于发散, 这表明树冠互补性主要反应了森林群落垂直生态位的分化, 且倾向于在长期的历史演化过程中参与谱系结构的构建过程。物种多样性和结构多样性随海拔升高而显著降低, 谱系结构也随海拔升高而趋于聚集, 表明地形因子表征的生境过滤与树冠互补性表征的生态位分化共同影响婺城南山落叶阔叶林的群落多样性。研究结果表明树冠互补性是森林群落多样性维持的重要驱动因素, 有利于婺城南山落叶阔叶林群落生物多样性维持和物种共存。未来研究可以探讨叶习性差异引起的时间生态位分化对树冠互补性与生物多样性关系的影响, 这将有助于深化对树冠结构在维持森林群落多样性中作用的理解。
| [1] |
Chesson P. Mechanisms of maintenance of species diversity. Annual Review of Ecology and Systematics, 2000, 31: 343-366. DOI:10.1146/annurev.ecolsys.31.1.343 |
| [2] |
HilleRisLambers J, Adler P B, Harpole W S, Levine J M, Mayfield M M. Rethinking community assembly through the lens of coexistence theory. Annual Review of Ecology, Evolution, and Systematics, 2012, 43: 227-248. DOI:10.1146/annurev-ecolsys-110411-160411 |
| [3] |
马克平, 徐学红. 中国森林生物多样性监测网络有力支撑生物群落维持机制研究. 中国科学: 生命科学, 2020, 50(4): 359-361. |
| [4] |
Anderson-Teixeira K J, Davies S J, Bennett A C, Gonzalez-Akre E B, Muller-Landau H C, Wright S J, Abu Salim K, Almeyda Zambrano A M, Alonso A, Baltzer J L, Basset Y, Bourg N A, Broadbent E N, Brockelman W Y, Bunyavejchewin S, Burslem D F R P, Butt N, Cao M, Cardenas D, Chuyong G B, Clay K, Cordell S, Dattaraja H S, Deng X B, Detto M, Du X J, Duque A, Erikson D L, Ewango C E N, Fischer G A, Fletcher C, Foster R B, Giardina C P, Gilbert G S, Gunatilleke N, Gunatilleke S, Hao Z Q, Hargrove W W, Hart T B, Hau B C H, He F L, Hoffman F M, Howe R W, Hubbell S P, Inman-Narahari F M, Jansen P A, Jiang M X, Johnson D J, Kanzaki M, Kassim A R, Kenfack D, Kibet S, Kinnaird M F, Korte L, Kral K, Kumar J, Larson A J, Li Y D, Li X K, Liu S R, Lum S K Y, Lutz J A, Ma K P, Maddalena D M, Makana J R, Malhi Y, Marthews T, Serudin R M, McMahon S M, McShea W J, Memiaghe H R, Mi X C, Mizuno T, Morecroft M, Myers J A, Novotny V, de Oliveira A A, Ong P S, Orwig D A, Ostertag R, den Ouden J, Parker G G, Phillips R P, Sack L, Sainge M N, Sang W G, Sri-Ngernyuang K, Sukumar R, Sun I F, Sungpalee W, Suresh H S, Tan S, Thomas S C, Thomas D W, Thompson J, Turner B L, Uriarte M, Valencia R, Vallejo M I, Vicentini A, Vrška T, Wang X H, Wang X G, Weiblen G, Wolf A, Xu H, Yap S, Zimmerman J. CTFS-ForestGEO: a worldwide network monitoring forests in an era of global change. Global Change Biology, 2015, 21(2): 528-549. DOI:10.1111/gcb.12712 |
| [5] |
Davies S J, Abiem I, Abu Salim K, Aguilar S, Allen D, Alonso A, Anderson-Teixeira K, Andrade A, Arellano G, Ashton P S, Baker P J, Baker M E, Baltzer J L, Basset Y, Bissiengou P, Bohlman S, Bourg N A, Brockelman W Y, Bunyavejchewin S, Burslem D F R P, Cao M, Cárdenas D, Chang L W, Chang-Yang C H, Chao K J, Chao W C, Chapman H, Chen Y Y, Chisholm R A, Chu C J, Chuyong G, Clay K, Comita L S, Condit R, Cordell S, Dattaraja H S, de Oliveira A A, den Ouden J, Detto M, Dick C, Du X J, Duque Á, Ediriweera S, Ellis E C, Obiang N L E, Esufali S, Ewango C E N, Fernando E S, Filip J, Fischer G A, Foster R, Giambelluca T, Giardina C, Gilbert G S, Gonzalez-Akre E, Gunatilleke I A U N, Gunatilleke C V S, Hao Z Q, Hau B C H, He F L, Ni H W, Howe R W, Hubbell S P, Huth A, Inman-Narahari F, Itoh A, Janík D, Jansen P A, Jiang M X, Johnson D J, Jones F A, Kanzaki M, Kenfack D, Kiratiprayoon S, Král K, Krizel L, Lao S, Larson A J, Li Y D, Li X K, Litton C M, Liu Y, Liu S R, Lum S K Y, Luskin M S, Lutz J A, Luu H T, Ma K P, Makana J R, Malhi Y, Martin A, McCarthy C, McMahon S M, McShea W J, Memiaghe H, Mi X C, Mitre D, Mohamad M, Monks L, Muller-Landau H C, Musili P M, Myers J A, Nathalang A, Ngo K M, Norden N, Novotny V, O'Brien M J, Orwig D, Ostertag R, Papathanassiou K, Parker G G, Pérez R, Perfecto I, Phillips R P, Pongpattananurak N, Pretzsch H, Ren H B, Reynolds G, Rodriguez L J, Russo S E, Sack L, Sang W G, Shue J, Singh A, Song G Z M, Sukumar R, Sun I F, Suresh H S, Swenson N G, Tan S, Thomas S C, Thomas D, Thompson J, Turner B L, Uowolo A, Uriarte M, Valencia R, Vandermeer J, Vicentini A, Visser M, Vrska T, Wang X G, Wang X H, Weiblen G D, Whitfeld T J S, Wolf A, Wright S J, Xu H, Yao T L, Yap S L, Ye W H, Yu M J, Zhang M H, Zhu D G, Zhu L, Zimmerman J K, Zuleta D. ForestGEO: Understanding forest diversity and dynamics through a global observatory network. Biological Conservation, 2021, 253: 108907. DOI:10.1016/j.biocon.2020.108907 |
| [6] |
Mi X C, Feng G, Hu Y B, Zhang J, Chen L, Corlett R T, Hughes A C, Pimm S, Schmid B, Shi S H, Svenning J C, Ma K P. The global significance of biodiversity science in China: an overview. National Science Review, 2021, 8(7): nwab032. |
| [7] |
Zhao N, Liu H M, Wang Q F, Wang R L, Xu Z W, Jiao C C, Zhu J X, Yu G R, He N P. Root elemental composition in Chinese forests: implications for biogeochemical niche differentiation. Functional Ecology, 2018, 32(1): 40-49. DOI:10.1111/1365-2435.12938 |
| [8] |
陈磊, 米湘成, 马克平. 生态位分化与森林群落物种多样性维持研究展望. 生命科学, 2014, 26(2): 112-117. |
| [9] |
柴永福, 岳明. 植物群落构建机制研究进展. 生态学报, 2016, 36(15): 4557-4572. |
| [10] |
Potts M D, Davies S J, Bossert W H, Tan S, Nur Supardi M N. Habitat heterogeneity and niche structure of trees in two tropical rain forests. Oecologia, 2004, 139(3): 446-453. DOI:10.1007/s00442-004-1525-3 |
| [11] |
Zhang J, Zhang Z C, Lutz J A, Chu C J, Hu J B, Shen G C, Li B H, Yang Q S, Lian J Y, Zhang M H, Wang X H, Ye W H, He F L. Drone-acquired data reveal the importance of forest canopy structure in predicting tree diversity. Forest Ecology and Management, 2022, 505: 119945. DOI:10.1016/j.foreco.2021.119945 |
| [12] |
Chen L, Swenson N G, Ji N N, Mi X C, Ren H B, Guo L D, Ma K P. Differential soil fungus accumulation and density dependence of trees in a subtropical forest. Science, 2019, 366(6461): 124-128. DOI:10.1126/science.aau1361 |
| [13] |
Kalyuzhny M, Lake J K, Wright S J, Ostling A M. Pervasive within-species spatial repulsion among adult tropical trees. Science, 2023, 381(6657): 563-568. DOI:10.1126/science.adg7021 |
| [14] |
Mi X C, Sun Z H, Song Y F, Liu X J, Yang J, Wu J J, Ci X Q, Li J, Lin L X, Cao M, Ma K P. Rare tree species have narrow environmental but not functional niches. Functional Ecology, 2021, 35(2): 511-520. DOI:10.1111/1365-2435.13714 |
| [15] |
Luo S, Phillips R P, Jo I, Fei S L, Liang J J, Schmid B, Eisenhauer N. Higher productivity in forests with mixed mycorrhizal strategies. Nature Communications, 2023, 14(1): 1377. DOI:10.1038/s41467-023-36888-0 |
| [16] |
Hildebrand M, Perles-Garcia M D, Kunz M, Härdtle W, von Oheimb G, Fichtner A. Reprint of: tree-tree interactions and crown complementarity: the role of functional diversity and branch traits for canopy packing. Basic and Applied Ecology, 2021, 55: 53-63. DOI:10.1016/j.baae.2021.01.010 |
| [17] |
余秋伍, 杨菁, 沈国春. 浙江天童常绿阔叶林林冠结构与群落物种组成的关系. 植物生态学报, 2022, 46(5): 529-538. |
| [18] |
Nakamura A, Kitching R L, Cao M, Creedy T J, Fayle T M, Freiberg M, Hewitt C N, Itioka T, Koh L P, Ma K P, Malhi Y, Mitchell A, Novotny V, Ozanne C M P, Song L, Wang H, Ashton L A. Forests and their canopies: achievements and horizons in canopy science. Trends in Ecology & Evolution, 2017, 32(6): 438-451. |
| [19] |
Georgi L, Kunz M, Fichtner A, Reich K F, Bienert A, Maas H G, von Oheimb G. Effects of local neighbourhood diversity on crown structure and productivity of individual trees in mature mixed-species forests. Forest Ecosystems, 2021, 8: 26. DOI:10.1186/s40663-021-00306-y |
| [20] |
Jucker T, Bouriaud O, Coomes D A. Crown plasticity enables trees to optimize canopy packing in mixed-species forests. Functional Ecology, 2015, 29(8): 1078-1086. DOI:10.1111/1365-2435.12428 |
| [21] |
郑芬, 李兆佳, 邱治军, 赵厚本, 周光益. 广东南岭天然常绿阔叶林林下光环境对林下幼树功能性状的影响. 生态学报, 2020, 40(13): 4516-4527. |
| [22] |
Dormann C F, Bagnara M, Boch S, Hinderling J, Janeiro-Otero A, Schäfer D, Schall P, Hartig F. Plant species richness increases with light availability, but not variability, in temperate forests understorey. BMC Ecology, 2020, 20(1): 43. DOI:10.1186/s12898-020-00311-9 |
| [23] |
Ishii H, Azuma W, Nabeshima E. The need for a canopy perspective to understand the importance of phenotypic plasticity for promoting species coexistence and light-use complementarity in forest ecosystems. Ecological Research, 2013, 28(2): 191-198. DOI:10.1007/s11284-012-1025-6 |
| [24] |
Chai P T, Xie J J, Yang L S, Zheng R, Bian Y X, Fu J Q, Wang Y Q, Chen J H. Community vertical stratification drives temporal taxonomic and phylogenetic beta diversity in a mixed broadleaf-conifer forest. Frontiers in Ecology and Evolution, 2023, 11: 1170197. DOI:10.3389/fevo.2023.1170197 |
| [25] |
Zheng L T, Chen H Y H, Yan E R. Tree species diversity promotes litterfall productivity through crown complementarity in subtropical forests. Journal of Ecology, 2019, 107(4): 1852-1861. DOI:10.1111/1365-2745.13142 |
| [26] |
Williams L J, Paquette A, Cavender-Bares J, Messier C, Reich P B. Spatial complementarity in tree crowns explains overyielding in species mixtures. Nature Ecology & Evolution, 2017, 1(4): 63. |
| [27] |
Zhang J, Huang S, He F L. Half-century evidence from western Canada shows forest dynamics are primarily driven by competition followed by climate. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(13): 4009-4014. |
| [28] |
Chen L, Wang Y Q, Mi X C, Liu X J, Ren H B, Chen J H, Ma K P, Kraft N J B. Neighborhood effects explain increasing asynchronous seedling survival in a subtropical forest. Ecology, 2019, 100(11): e02821. DOI:10.1002/ecy.2821 |
| [29] |
Wang Y Q, Cadotte M W, Chen J H, Mi X C, Ren H B, Liu X J, Yu M J, Zhang J, Ma K P. Neighborhood interactions on seedling survival were greatly altered following an extreme winter storm. Forest Ecology and Management, 2020, 461: 117940. DOI:10.1016/j.foreco.2020.117940 |
| [30] |
杨永川, 达良俊, 由文辉. 浙江天童国家森林公园微地形与植被结构的关系. 生态学报, 2005, 25(11): 2830-2840. |
| [31] |
Chapman J, McEwan R. The role of environmental filtering in structuring Appalachian tree communities: topographic influences on functional diversity are mediated through soil characteristics. Forests, 2018, 9(1): 19. DOI:10.3390/f9010019 |
| [32] |
He R, Hu M, Shi H, Zhou Q, Shu X, Zhang K R, Zhang Q F, Dang H S. Patterns of species diversity and its determinants in a temperate deciduous broad-leaved forest. Forest Ecosystems, 2022, 9: 100062. DOI:10.1016/j.fecs.2022.100062 |
| [33] |
Gianoli E, Escobedo V M. Phenotypic plasticity may mediate habitat filtering in a forest edge community. Oikos, 2021, 130(10): 1788-1796. DOI:10.1111/oik.08455 |
| [34] |
Yuan Z Q, Ali A, Ruiz-Benito P, Jucker T, Mori A S, Wang S P, Zhang X K, Li H, Hao Z Q, Wang X G, Loreau M. Above- and below-ground biodiversity jointly regulate temperate forest multifunctionality along a local-scale environmental gradient. Journal of Ecology, 2020, 108(5): 2012-2024. DOI:10.1111/1365-2745.13378 |
| [35] |
Jucker T, Hardwick S R, Both S, Elias D M O, Ewers R M, Milodowski D T, Swinfield T, Coomes D A. Canopy structure and topography jointly constrain the microclimate of human-modified tropical landscapes. Global Change Biology, 2018, 24(11): 5243-5258. DOI:10.1111/gcb.14415 |
| [36] |
Feng G, Mi X C, Yan H, Li F Y, Svenning J C, Ma K P. CForBio: a network monitoring Chinese forest biodiversity. Science Bulletin, 2016, 61(15): 1163-1170. DOI:10.1007/s11434-016-1132-9 |
| [37] |
Onoda Y, Saluñga J B, Akutsu K, Aiba S I, Yahara T, Anten N P R. Trade-off between light interception efficiency and light use efficiency: implications for species coexistence in one-sided light competition. Journal of Ecology, 2014, 102(1): 167-175. DOI:10.1111/1365-2745.12184 |
| [38] |
马克平, 刘玉明. 生物群落多样性的测度方法Ⅰα多样性的测度方法(下). 生物多样性, 1994, 2(4): 231-239. |
| [39] |
Jin Y, Qian H. V. PhyloMaker2: an updated and enlarged R package that can generate very large phylogenies for vascular plants. Plant Diversity, 2022, 44(4): 335-339. DOI:10.1016/j.pld.2022.05.005 |
| [40] |
Smith S A, Brown J W. Constructing a broadly inclusive seed plant phylogeny. American Journal of Botany, 2018, 105(3): 302-314. DOI:10.1002/ajb2.1019 |
| [41] |
Zanne A E, Tank D C, Cornwell W K, Eastman J M, Smith S A, FitzJohn R G, McGlinn D J, O'Meara B C, Moles A T, Reich P B, Royer D L, Soltis D E, Stevens P F, Westoby M, Wright I J, Aarssen L, Bertin R I, Calaminus A, Govaerts R, Hemmings F, Leishman M R, Oleksyn J, Soltis P S, Swenson N G, Warman L, Beaulieu J M. Three keys to the radiation of angiosperms into freezing environments. Nature, 2014, 506(7486): 89-92. DOI:10.1038/nature12872 |
| [42] |
Webb C O, Ackerly D D, Kembel S W. Phylocom: software for the analysis of phylogenetic community structure and trait evolution. Bioinformatics, 2008, 24(18): 2098-2100. DOI:10.1093/bioinformatics/btn358 |
| [43] |
Kembel S W, Cowan P D, Helmus M R, Cornwell W K, Morlon H, Ackerly D D, Blomberg S P, Webb C O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics, 2010, 26(11): 1463-1464. DOI:10.1093/bioinformatics/btq166 |
| [44] |
Zhang Y, Chen H Y H. Individual size inequality links forest diversity and above-ground biomass. Journal of Ecology, 2015, 103(5): 1245-1252. DOI:10.1111/1365-2745.12425 |
| [45] |
姚志良, 温韩东, 邓云, 曹敏, 林露湘. 哀牢山亚热带中山湿性常绿阔叶林树种beta多样性格局形成的驱动力. 生物多样性, 2020, 28(4): 445-454. |
| [46] |
Le Provost G, Badenhausser I, Le Bagousse-Pinguet Y, Clough Y, Henckel L, Violle C, Bretagnolle V, Roncoroni M, Manning P, Gross N. Land-use history impacts functional diversity across multiple trophic groups. Proceedings of the National Academy of Sciences of the United States of America, 2020, 117(3): 1573-1579. |
| [47] |
Liu S G, Plaza C, Ochoa-Hueso R, Trivedi C, Wang J T, Trivedi P, Zhou G Y, Piñeiro J, Martins C S C, Singh B K, Delgado-Baquerizo M. Litter and soil biodiversity jointly drive ecosystem functions. Global Change Biology, 2023, 29(22): 6276-6285. DOI:10.1111/gcb.16913 |
| [48] |
Moura M R, Costa H C, Peixoto M A, Carvalho A L G, Santana D J, Vasconcelos H L. Geographical and socioeconomic determinants of species discovery trends in a biodiversity hotspot. Biological Conservation, 2018, 220: 237-244. DOI:10.1016/j.biocon.2018.01.024 |
| [49] |
Ren L, Huo J X, Xiang X, Pan Y P, Li Y Q, Wang Y Y, Meng D H, Yu C, Chen Y, Xu Z C, Huang Y M. Environmental conditions are the dominant factor influencing stability of terrestrial ecosystems on the Tibetan Plateau. Communications Earth & Environment, 2023, 4: 196. |
| [50] |
Xu Y Z, Chen H Y H, Xiao Z Q, Wan D, Liu F, Guo Y L, Qiao X J, Jiang M X. Species richness promotes productivity through tree crown spatial complementarity in a species-rich natural forest. Forests, 2022, 13(10): 1604. DOI:10.3390/f13101604 |
| [51] |
Pretzsch H. Canopy space filling and tree crown morphology in mixed-species stands compared with monocultures. Forest Ecology and Management, 2014, 327: 251-264. DOI:10.1016/j.foreco.2014.04.027 |
| [52] |
李梦佳, 何中声, 江蓝, 谷新光, 晋梦然, 陈博, 刘金福. 戴云山物种多样性与系统发育多样性海拔梯度分布格局及驱动因子. 生态学报, 2021, 41(3): 1148-1157. |
| [53] |
Starko S, Demes K W, Neufeld C J, Martone P T. Convergent evolution of niche structure in Northeast Pacific kelp forests. Functional Ecology, 2020, 34(10): 2131-2146. DOI:10.1111/1365-2435.13621 |
| [54] |
贾鹏, 杜国祯. 生态学的多样性指数: 功能与系统发育. 生命科学, 2014, 26(2): 153-157. |
| [55] |
Ishii H, Asano S. The role of crown architecture, leaf phenology and photosynthetic activity in promoting complementary use of light among coexisting species in temperate forests. Ecological Research, 2010, 25(4): 715-722. DOI:10.1007/s11284-009-0668-4 |
| [56] |
Ali A. Forest stand structure and functioning: current knowledge and future challenges. Ecological Indicators, 2019, 98: 665-677. DOI:10.1016/j.ecolind.2018.11.017 |
| [57] |
Marselis S M, Tang H, Armston J, Abernethy K, Alonso A, Barbier N, Bissiengou P, Jeffery K, Kenfack D, Labrière N, Lee S K, Lewis S L, Memiaghe H, Poulsen J R, White L, Dubayah R. Exploring the relation between remotely sensed vertical canopy structure and tree species diversity in Gabon. Environmental Research Letters, 2019, 14(9): 094013. DOI:10.1088/1748-9326/ab2dcd |
| [58] |
Montgomery R A, Chazdon R L. Forest structure, canopy architecture, and light transmittance in tropical wet forests. Ecology, 2001, 82(10): 2707. DOI:10.1890/0012-9658(2001)082[2707:FSCAAL]2.0.CO;2 |
| [59] |
He X R, Arif M, Zheng J, Ni X L, Yuan Z X, Zhu Q, Wang J F, Ding D D, Li C X. Plant diversity patterns along an elevation gradient: the relative impact of environmental and spatial variation on plant diversity and assembly in arid and semi-arid regions. Frontiers in Environmental Science, 2023, 11: 1021157. DOI:10.3389/fenvs.2023.1021157 |
| [60] |
Dani R S, Divakar P K, Baniya C B. Diversity and composition of plants species along elevational gradient: research trends. Biodiversity and Conservation, 2023, 32(8): 2961-2980. |
| [61] |
盘远方, 李娇凤, 姚玉萍, 姜勇, 利恒春, 王晓凤, 卢国琼, 杨晨, 黄诗雯, 蒋文平. 桂林岩溶石山青冈群落植物功能多样性和环境因子与坡向的关联研究. 生态学报, 2021, 41(11): 4484-4492. |
| [62] |
De Cáceres M, Legendre P, Valencia R, Cao M, Chang L W, Chuyong G, Condit R, Hao Z Q, Hsieh C F, Hubbell S, Kenfack D, Ma K P, Mi X C, Supardi Noor M N, Kassim A R, Ren H B, Su S H, Sun I F, Thomas D, Ye W H, He F L. The variation of tree beta diversity across a global network of forest plots. Global Ecology and Biogeography, 2012, 21(12): 1191-1202. |
| [63] |
Wang B W, Zhang G H, Duan J. Relationship between topography and the distribution of understory vegetation in a Pinus massoniana forest in Southern China. International Soil and Water Conservation Research, 2015, 3(4): 291-304. |
| [64] |
杨欣, 姚志良, 王彬, 温韩东, 邓云, 曹敏, 张志明, 谭正洪, 林露湘. 亚热带常绿阔叶林林分结构对物种组成变异的驱动作用: 从局域到区域尺度. 生物多样性, 2023, 31(2): 5-16. |
| [65] |
霍兵兵, 孙哲明, 欧文慧, 毛鸿志, 胡傲, 杨予静, 李中强. 环境筛选和扩散限制对长江流域湖北段湿地植物群落构建的共同影响. 生态学报, 2023, 43(5): 1804-1811. |
| [66] |
Yeboah D, Chen H Y H, Kingston S. Tree species richness decreases while species evenness increases with disturbance frequency in a natural boreal forest landscape. Ecology and Evolution, 2016, 6(3): 842-850. |
| [67] |
Mao L F, Dong Y R, Xing B B, Chen Y H, Dennett J, Bater C, Stadt J J, Nielsen S E. Maximum canopy height is associated with community phylogenetic structure in boreal forests. Journal of Plant Ecology, 2023, 16(4): rtac104. |
| [68] |
LaRue E A, Knott J A, Domke G M, Chen H Y, Guo Q F, Hisano M, Oswalt C, Oswalt S, Kong N, Potter K M, Fei S L. Structural diversity as a reliable and novel predictor for ecosystem productivity. Frontiers in Ecology and the Environment, 2023, 21(1): 33-39. |
| [69] |
Wang S P, Hong P B, Adler P B, Allan E, Hautier Y, Schmid B, Spaak J W, Feng Y H. Towards mechanistic integration of the causes and consequences of biodiversity. Trends in Ecology & Evolution, 2024, 39(7): 689-700. |
| [70] |
Verheyen K, Gillerot L, Blondeel H, De Frenne P, De Pauw K, Depauw L, Lorer E, Sanczuk P, Schreel J, Vanneste T, Wei L P, Landuyt D. Forest canopies as nature-based solutions to mitigate global change effects on people and nature. Journal of Ecology, 2024, 112(11): 2451-2461. |