 2025, Vol. 45
2025, Vol. 45文章信息
- 柴思帆, 宋献方, 卜红梅
- CHAI Sifan, SONG Xianfang, BU Hongmei
- 西藏中部河湖岸带湿地植物优势种碳氮化学计量特征
- Carbon and nitrogen stoichiometric characteristics of dominant wetland plants in riparian zones in Central Tibet
- 生态学报. 2025, 45(6): 2558-2570
- Acta Ecologica Sinica. 2025, 45(6): 2558-2570
- http://dx.doi.org/10.20103/j.stxb.202405161122
-
文章历史
- 收稿日期: 2024-05-16
- 网络出版日期: 2024-12-23





2. 中国科学院大学资源与环境学院, 北京 100049
2. College of Resources and Environment, University of Chinese Academy of Sciences, Beijing 100049, China
青藏高原具有独特的地理、气候和生态条件, 是中国湿地分布最集中的地区, 孕育了丰富多样的湿地生态系统[1]。这些湿地生态系统不仅是青藏高原生态安全的重要屏障, 也是全球气候变化响应的敏感区域。湿地植物作为湿地生态系统的重要组成部分, 其碳氮化学计量特征对于理解湿地生态系统的功能、稳定性和对环境变化的响应具有重要意义[2]。
碳(C)和氮(N)元素是形成植物组织结构和维持植物生长发育的基本元素, 也是影响湿地植物生长、代谢和生态功能的关键因素。在湿地生态系统中, 植物通过光合作用固定碳, 并通过生物地球化学循环过程参与氮的转化和循环。植物的碳氮化学计量特征(如C ∶ N比)不仅反映了植物对碳氮元素的吸收、利用和储存能力等生长策略, 也反映了植物对环境的适应性[2]。中国52个地区湿地植物N生态化学计量特征研究的结果表明, 中国大部分湿地植物叶片表现为N限制, 湿地类型决定着植物叶片的N生态化学计量特征, 河流类型的植物其N含量值最高, 沼泽湿地类型的植物其N含量值最低, 而植物碳氮化学计量特征之间的差异受到物种组成、植物生活型和环境等因素的影响[3]。在中国不同湖区中, 青藏高原湖区草本植物具有较高的N含量, 这可能与青藏高原独特的气候、地理条件(高海拔、低温、低纬度等)有关[4];青藏高原35种湿地植物测得的叶片养分浓度随着温度的降低而增加, 表明在青藏高原低温诱导下产生了养分积累, 与温度-植物生理假说(Temperature-plant physiological hypothesis, TPPH)一致[5]。
植物的碳氮同位素组成(δ13C和δ15N)是陆地生态系统碳氮循环的综合结果, 能够反映植物的新陈代谢和碳氮循环影响因子的综合作用[6—8], 其分布特征能较好的反映研究区域内的物源信息和生物地球化学特征[9]。植物的δ13C可以准确记录与植物生长过程相联系的气候环境信息。研究发现, 叶片δ13C与植物水分利用效率呈显著相关, 是反映植物水分利用的有效指标之一[10—11], 而植物水分利用效率又与叶片生态化学计量特征密切相关。植物δ15N可以反映外源输入, 污染物输入负荷越大, 植物的δ15N越高。太湖流域贡湖湾和梅梁湾沉水植物δ13C和δ15N的不同变化表明了两个湖区水体环境质量的明显差异[12]。城市河流水生植物的δ15N值也能反映植物对水体氮的利用策略[13], 其水分传输路径不同[14]、碳氮元素来源不同及吸收营养盐的效能差异, 导致不同生活型的湿地植物具有不同的碳氮同位素分布[15]。
近年来, 随着全球气候变化和人类活动的加剧, 青藏高原湿地生态系统面临着前所未有的压力和挑战。海拔梯度和经纬度的差异包含了水、热和光照等重要环境因子的变化, 可能改变湿地植物的碳氮化学计量特征, 进而影响湿地生态系统的结构和功能[16]。西藏中部地区, 从西部的阿里到那曲, 再到东部的拉萨、林芝和昌都, 平均海拔由4500 m以上到3040 m, 横跨了多个海拔梯度, 湿地植物具有显著的地理和生态多样性, 其地理信息的变化为湿地植物的生长提供了多样化的环境。湿地植物优势种在各自的生态位中占据了主导地位, 当地的优势种具有普遍性, 相对丰度高, 能够通过调节植物群落功能结构直接或间接对生态系统功能产生显著影响[17], 同时也能够通过对环境变化的敏感响应, 较好地反映生态系统的健康状况。因此, 本研究通过系统分析西藏中部河湖岸带湿地植物优势种的碳氮化学计量特征变化规律, 探讨植物碳氮化学计量特征与地理因子之间的关系, 明确湿地植物的生长策略和营养分配模式, 揭示地理因子对西藏湿地植物的影响。本研究将为青藏高原维持生态屏障功能和生态修复提供理论依据。
1 材料与方法 1.1 研究区概况西藏(78°24′E—99°06′E, 26°52′N—36°32′N)位于青藏高原西南部, 全区总面积122.84万km2, 地形复杂多样, 分为藏北高原、藏南谷地、藏东高山峡谷和喜马拉雅山地四个地带。本研究所调查的西藏中部河湖岸带湿地植物范围自西向东包括阿里、那曲、拉萨、林芝和昌都地区(图 1), 涉及面积共98.93万km2, 占全区总面积的80.5%, 平均海拔4000 m以上, 地势由西北向东南倾斜, 气候类型复杂, 呈现西北严寒干燥、东南温暖湿润的总趋向。各地降水的季节分配不均, 干季和雨季的分界明显, 年降水量自东南低地的5000 mm逐渐向西北递减到50 mm[18]。西藏湿地植物的种类主要有高草湿地植被、低草湿地植被、沉水型湿地植被、浮水型湿地植被和挺水型湿地植被。最新出版的《西藏湿地植物》详细记录了西藏345种湿地植物, 隶属63科202属[19]。其中, 禾本科、菊科、莎草科和豆科是最主要的植物类型, 而蕨麻、丝叶眼子菜、西伯利亚蓼、两栖蓼和华扁穗草等部分物种对维持西藏湿地生态平衡和生物多样性具有重要作用。

|
| 图 1 西藏中部河湖岸带湿地植物优势种采样点图 Fig. 1 Sampling sites of dominant wetland plants in riparian zones in Central Tibet |
分别于2019年和2023年8月在西藏中部地区开展河湖岸带湿地植物优势种采样(图 1)。2023年8月在西藏阿里地区的日土县、革吉县和改则县、那曲市的尼玛县、申扎县和班戈县以及拉萨市的当雄县, 沿河湖岸带分别布设9个采样点(图 2), 分别为班公湖(S1)、狮泉河上游(S2)、别若则错(S3)、洞错(S4)、当穹错(S5)、当惹雍错(S6)、色林错(S7)、巴木错(S8)和纳木错(S9)。2019年8月, 在林芝地区和昌都市布设4个采样点(表 1), 分别为尼洋河岸边(S10)、雅鲁藏布江岸边(S11)、米林县境内雅鲁藏布江岸边(S12)及邦达镇318国道西南(S13)。两次调查中, 共采集河湖岸带湿地植物优势种14种, 隶属于8科12属, 其中莎草科植物1种, 禾本科植物4种, 蓼科植物3种, 苋科植物1种, 菊科植物1种, 豆科植物2种, 报春花科1种, 木贼科1种(表 1)。

|
| 图 2 西藏中部河湖岸带湿地植物优势种采样点地理位置 Fig. 2 Geographic location of sampling sites for dominant wetland plants in riparian zones in Central Tibet |
| 植物名称Plant species | 科Family | 属Genus | 生活型Life form |
| 华扁穗草Blysmus sinocompressus | 莎草科 | 扁穗草属 | 多年生草本 |
| 中华草沙蚕Tripogon chinensis | 禾本科 | 草沙蚕属 | 多年生草本 |
| 牛筋草Eleusine indica | 禾本科 | 属 | 一年生草本 |
| 西伯利亚蓼Polygonum sibiricum | 蓼科 | 蓼属 | 多年生草本 |
| 平卧藜Chenopodium prostratum | 苋科 | 藜属 | 一年生草本 |
| 细叶西伯利亚蓼Polygonum sibiricum var. thomsonii | 蓼科 | 蓼属 | 一年生草本 |
| 麦薲草Elymus tangutorum | 禾本科 | 披碱草属 | 多年生草本 |
| 藏沙蒿Artemisia wellbyi | 菊科 | 蒿属 | 半灌木状草本 |
| 密花黄芪Astragalus membranaceus | 豆科 | 黄芪属 | 多年生草本 |
| 镰荚棘豆Oxytropis falcata | 豆科 | 棘豆属 | 多年生草本 |
| 海乳草Glaux maritima | 报春花科 | 海乳草属 | 多年生草本 |
| 白草Pennisetum flaccidum Griseb | 禾本科 | 狼尾草属 | 多年生草本 |
| 节节草Equisetum ramosissimum | 木贼科 | 木贼属 | 多年生草本 |
| 水蓼Polygonum hydropiper | 蓼科 | 蓼属 | 一年生草本 |
在采样区域, 观察植物群落, 选取个体数量多、生物量较大、枝叶覆盖地面程度大的优势种类进行采样。在采样过程中, 湿地植物整个植株连根被全部挖起, 再用湖水清洗掉附着物, 按种类分开, 称量鲜重, 所有样品放在保温箱内冷藏带回。在实验室内, 将湿地植物样品洗净后放入烘箱, 105 ℃下杀青30 min, 65 ℃烘干至恒重。在研磨机上, 将烘干的样品磨成粉末, 用200目筛网筛样品粉末, 用锡箔纸包裹后干燥保存, 以便进行后续测定。利用元素分析仪(Elementar vario MACRO cube, German)测定湿地植物土样的CN含量。CN含量采用质量含量(mg/g)表示, C ∶ N比为其两者之间的质量比。利用稳定同位素比质谱仪(Finnigan Mat 253; Thermo Fisher Scientific, Bremen, Germany)测定稳定碳氮同位素, 测得的碳氮稳定同位素值用‰表示, 分别表示为δ13C和δ15N(公式1)。
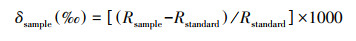
|
(1) |
其中, Rsample分别代表13C/12C或15N/14N比值, C同位素测定的标准物质为PDB(美洲拟箭石), N同位素的标准物质为空气中的N2, 仪器分析重复样品的精度为0.15‰。
1.3 数据分析与处理用单因素方差分析(One-way ANOVA)分别比较不同种类、不同采样点、不同生活型湿地植物C、N含量及其稳定同位素δ13C和δ15N值的空间差异, 显著性检验水平为P < 0.05或P < 0.01。在进行方差分析前, 用Shapiro-Wilk法检验原始数据的正态性, 用Bartlett法检验方差齐性。对于不满足正态分布的C含量和δ13C值, 选用非参数法Kruskal-Wallis秩和检验进行方差分析。采用相关分析(Correlation analysis), 分析湿地植物C和N变量之间、以及与植物含水率、地理因子之间的相关性, 显著性水平分别为P < 0.05或P < 0.01。采用线性回归分析方法, 构建湿地植物C和N变量之间、以及与植物含水率、地理因子之间的线性关系(P < 0.05或P < 0.01)。
利用含水率(η)测定西藏湿地植物体内水分含量占整株植物总重量的比例, 以百分比表示。利用种间变异系数(CVb)衡量不同种湿地植物个体之间在CN含量、C ∶ N比及其稳定同位素δ13C和δ15N值方面的变异程度, 反映不同种植物的差异程度。利用种内变异系数(CVw)衡量同一种湿地植物个体内部在CN含量、C ∶ N比及其稳定同位素δ13C和δ15N值方面的变异程度, 反映同一种植物个体间的差异程度。
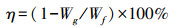
|
(2) |
其中, η为湿地植物含水率, Wg为湿地植物的干重, Wf为湿地植物的鲜重。
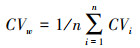
|
(3) |
其中, CVw为种内变异系数, n为西藏湿地植物物种数, i为湿地植物某物种, CVi为湿地植物物种i的变异系数。
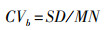
|
(4) |
其中, CVb为种间变异系数, SD为所有湿地植物物种平均值的标准差, MN为所有湿地植物物种平均值。
以上数据分析及制图过程在应用软件SPSS 26和origin 2021中完成。
2 结果与分析 2.1 湿地植物优势种C、N化学计量学特征对采集的湿地植物优势种的CN含量、C ∶ N比、及其δ13C和δ15N值进行Shapiro-Wilk正态性检验。检验结果表明, 湿地植物的N含量、C ∶ N比和δ15N值呈显著的正态分布(P>0.05)。湿地植物优势种的C含量范围为(105.97—433.25)mg/g(图 3), 均值为(325.01±83.54)mg/g(平均值±方差), 平均变异系数为0.26;N含量的范围为(42.40—4.87)mg/g, 均值为(20.76±8.54)mg/g, 平均变异系数为0.41;C ∶ N比在8.82—27.02之间, 均值为(17.20±5.01), 平均变异系数为0.29;δ13C和δ15N值的变化范围分别为(-29.15—-12.49)‰和(-0.612—24.22)‰, 均值分别为(-24.99±4.03)‰和(9.69±6.59)‰, 平均变异系数分别为0.16和0.68。湿地植物含水率的范围为(64—94.65)%, 均值为(85.27±7.43)%, 平均变异系数为0.09。

|
| 图 3 西藏中部河湖岸带湿地植物优势种和采样点C、N变量及含水率特征箱线图 Fig. 3 Boxplots of C and N variables and water content characteristics of dominant species and sampling sites of wetland plants in riparian zones in Central Tibet 图中实线的上下端代表最大值和最小值, 矩形的上下端代表上四分位数和下四分位数, 横线为中位数, 空心正方形代表平均值 |
方差分析结果表明, 不同种类湿地植物优势种的CN含量、C ∶ N比、δ13C和δ15N值均呈现显著差异(P < 0.05;图 3)。其中, 中华草沙蚕的C含量最小, 平均值为(166.3±52.95)mg/g;藏沙蒿C含量最高, 平均值达到(423.7±14.32)mg/g, 是中华草沙蚕的2.5倍。西伯利亚蓼的N含量最高, 为(39.47±4.64)mg/g, 中华草沙蚕的N含量最低, 为(6.57±1.54)mg/g。西伯利亚蓼的C ∶ N比最低, 平均值为(9.9±1.26);中华草沙蚕的C ∶ N比值最高, 为(24.94±2.81)。西伯利亚蓼δ13C的平均值最低, 为(-28.22±0.14)‰, 平卧藜δ13C的平均值最大, 为(-13.22±0.16)‰;平卧藜δ15N的平均值最高, 为(21.62±2.50)‰, 藏沙蒿的δ15N的平均值最低, 为(2.07±3.08)‰, 变异系数为1.48。
西藏中部河湖岸带湿地植物优势种的C、N变量呈现出种间差异大于种内差异的特征(表 2)。其中, δ13C的种内和种间变异系数均为最小, 分别为0.05和0.16;δ15N的种内和种间变异系数均为最大, 分别为0.44和0.68;N含量的种内和种间变异系数较为居中, 分别为0.20和0.41;C和C ∶ N比的变异系数相近, 种内变异系数分别为0.13和0.14;种间变异系数分别为0.26和0.29。
| C/(mg/g) | N/(mg/g) | C∶N比 C∶N ratio |
δ13C/‰ | δ15N/‰ | |
| 种内变异系数Coefficient of variation within species | 0.13 | 0.20 | 0.14 | 0.05 | 0.44 |
| 种间变异系数Coefficient of variation between species | 0.26 | 0.41 | 0.29 | 0.16 | 0.68 |
将所有植物按不同生活型进行分类, 分析不同生活型植物CN含量、C ∶ N比及其稳定同位素特征(表 3)。不同生活型植物的C含量、N含量、C ∶ N比以及δ13C值之间的差异不显著, 但一年生草本的δ15N值(15.45‰)显著高于多年生草本植物(8.68‰)和半灌木状草本植物(2.07‰)(P < 0.05)。多年生草本植物N含量(21.46 mg/g)高于一年生(18.67 mg/g)和半灌木状草本(19.89 mg/g)植物。同样, δ13C值呈现出与δ15N值一致的趋势:一年生草本(-22.82‰)>多年草本(-25.48‰)>半灌木状草本(-26.74‰)。
| 植物生活型 Life form |
样本数/个 Sample size/number |
C/(mg/g) | N/(mg/g) | C∶N比 C∶N ratio |
δ13C/‰ | δ15N/‰ |
| 多年生草本Perennial herbs | 34 | 320.53 | 21.46 | 16.95 | -25.48 | 8.68 |
| 一年生草本Annuals herbs | 10 | 310.63 | 18.67 | 16.63 | -22.82 | 15.45 |
| 半灌木状草本Semi-shrubbery herbs | 3 | 423.70 | 19.89 | 22.00 | -26.74 | 2.07 |
湿地植物优势种的C、N变量在不同采样点之间也表现出差异性(图 3)。其中N含量和δ13C值在各采样点之间无显著差异(P>0.05), 而C ∶ N比呈现出显著的空间差异(P < 0.05), C含量和δ15N值则呈现出极显著的空间差异性(P < 0.01)。在西藏中部湿地植物优势种不同采样点的CN含量中, C含量最高为406.03 mg/g, 出现在纳木错(S9), 最低为226.76 mg/g, 出现在雅鲁藏布江岸边(S11);N含量在别若则错(S3)最高, 为29.18 mg/g, 在雅鲁藏布江岸边(S11)最低, 为12.56 mg/g;C ∶ N比的最高比值和最低比值为23.31和13.09, 分别出现在班公湖(S1)和巴木错(S8)。δ13C值在不同采样点的范围为-27.33‰—-16.88‰, 最高值和最低值分别出现在雅鲁藏布江岸边(S11)和色林错(S7);δ15N值在不同采样点的范围为-0.61‰—17.32‰, 最高值和最低值分别出现在巴木错(S8)和尼洋河岸边(S10);在采样区内, 西部(S1—S4)、中部(S5—S9)和东部(S10—S13)的δ15N值具有极显著的空间差异(P < 0.01), 自西向东呈现出逐渐降低的趋势。
2.2 湿地植物优势种C、N化学计量学特征之间的关系相关分析和线性回归分析表明, 西藏中部采集的河湖岸带湿地植物碳氮化学计量之间存在显著的相关性(表 4)和线性关系(图 4)。其中, 湿地植物的C与N含量之间呈现极显著的线性回归关系, 决定系数R2为0.275(P < 0.001);C ∶ N比与N含量之间也存在极显著的负相关关系, 相关系数r值为0.776(P < 0.01), 且两者之间具有较强的线性回归关系(图 4), 决定系数R2为0.582(P < 0.001)。另外, δ15N值与C含量、C ∶ N比和δ13C之间也存在显著的相关性, 决定系数R2分别为0.105(P=0.015)、0.092(P=0.022)和0.072(P=0.038)。
| C/(mg/g) | N/(mg/g) | C∶N比 | δ13C/‰ | δ15N/‰ | 含水率/% | 经度/(°) | 纬度/(°) | 海拔/m | |
| C/(mg/g) | 1 | ||||||||
| N/(mg/g) | 0.539** | 1 | |||||||
| C∶N比C∶N ratio | 0.011 | -0.769** | 1 | ||||||
| δ13C/‰ | -0.227 | -0.200 | -0.007 | 1 | |||||
| δ15N/‰ | -0.353* | 0.040 | -0.334* | 0.303* | 1 | ||||
| 含水率Moisture content/% | 0.16 | 0.486** | -0.498** | -0.051 | 0.628** | 1 | |||
| 经度Longititude/(°) | 0.023 | 0.041 | -0.179 | 0.079 | -0.417** | -0.421** | 1 | ||
| 纬度Latitude/(°) | 0.038 | -0.034 | 0.219 | -0.142 | 0.421** | 0.400** | -0.944** | 1 | |
| 海拔Altitude/m | 0.231 | 0.237 | -0.116 | -0.279 | 0.321* | 0.570** | -0.388** | 0.512** | 1 |
| * * 表示在0.01级别(双尾)相关性显著;* 表示在0.05级别(双尾)相关性显著 | |||||||||

|
| 图 4 湿地植物优势种C、N化学计量特征之间及其与含水率和地理因子之间的线性关系 Fig. 4 Linear regressions between C and N stoichiometric characteristics of dominant wetland plant and water content and geographic factors 阴影部分为95%置信区间 |
相关分析和线性回归分析表明, 湿地植物优势种碳氮变量及含水率与采样点的经度、纬度和海拔梯度之间也存在显著的相关性(表 4)和线性关系(图 4)。湿地植物含水率与经纬度、海拔梯度之间均极显著相关(P < 0.01), 其中与海拔梯度的相关系数最高, 为0.570。此外, 在线性回归分析中, 海拔梯度解释了30.2%的植物含水率(P < 0.001)。湿地植物优势种的含水率与其N含量和δ15N值之间呈极显著正相关关系(P < 0.01), 与C ∶ N比之间呈极显著负相关(P < 0.01), 相关系数分别为0.486、0.628和-0.498。在线性回归分析中, 湿地植物N含量、C ∶ N比和δ15N值分别解释了22.2%、23.9%和38.3%的植物含水率(P < 0.001)。湿地植物优势种的δ15N值与采样点的经度和纬度之间分别呈极显著负相关和极显著正相关关系(P < 0.01), 与海拔梯度呈显著正相关(P < 0.05), 相关系数分别为-0.417、0.421和0.321;在线性回归分析中, 经度、纬度与海拔梯度分别解释了湿地植物δ15N值15.6%(P=0.004)、15.9%(P=0.003)和8.3%(P=0.028)的总体变异。
3 讨论 3.1 河湖岸带湿地植物优势种化学计量特征与其他地区比较植物的碳氮含量受到多种因素的影响, 如植物生活型、环境养分供应、气候和地形因素等[2], 因此在区域尺度上碳氮化学计量特征存在较大的空间差异。本研究区的湿地植物C、N化学计量学特征与其他地区相比存在显著差异。西藏中部湿地植物的C含量平均为(325.01±83.54)mg/g, 显著低于全球植物叶片总有机碳含量(464 mg/g)[20], 也低于长江中下游湖泊湿地植物(339.42—435.45 mg/g)[21—22]、贵州草海湿地植物(373.08 mg/g)[23]、内蒙古草原锡林河植物(411.35±1.35)mg/g[24]和银川平原湿地植物(410.41—424.11 mg/g)[25]的C含量, 说明西藏中部湿地植物的碳储存能力相对较弱。与荒漠带草原的草本植物叶片C含量(357 mg/g)[26]相比, 其值相近, 且研究区与荒漠带草原环境类似, 湿度低、气候干旱、蒸发量远大于降水量且日照时间较长。植物体内的营养成分是对其所在生长环境生态特性的反映[27], 因此非生物环境因素可能对其C含量产生了影响。
研究区湿地植物N的平均含量((20.76±8.54) mg/g)与青藏高原河湖岸带湿生植物N含量((20.51±0.8) mg/g)的研究结果较为一致[4], 与全球植被N平均含量值接近(20.10 mg/g), 超过了中国753种陆地植被叶片总氮平均含量((20.20±8.41) mg/g)及中国主要湿地植物的平均N含量((19.61±16.24) mg/g)[3, 28—29], 这可能与样本植物的生活型有关。相较于灌木和乔木, 草本植物的单位质量叶氮含量一般较高[30]。在本研究中, 样品均来自草本植物, 因此其N含量差异明显[31]。此外, 青藏高原高海拔、低温的地理和气候条件也可能是导致该区域植物含有较高N的原因[32]。研究发现, 青藏高原地区群落水平叶面氮含量随海拔升高而增加[33]。海拔升高通常伴随着温度的降低, 当环境温度相对较低时, 植被维持较高的叶片N含量以维持代谢。本研究区海拔在3000—5000 m范围内, 因此, 该地区的叶片N含量高于其他地区[34]。
本研究中δ13C值的范围(-29.15‰—-12.49‰)落在青藏高原东缘植物叶片δ13C值的范围内(-35.73‰—-12.82‰)[35];而δ15N值(-0.61‰—24.22‰)较高, 则可能是受到周围环境因素的影响。有研究指出, 干旱条件促进土壤反硝化作用, 导致大量15N贫化的气态氮损失, 使剩余的可利用氮富集15N, 造成叶片的δ15N增加[36]。因此, 干旱地区植物的δ15N值大于温润地区[37]。此外, 干旱条件下植物可能被迫利用含δ15N更多、更深层次的土壤水分, 从而导致叶片的δ15N增加[38—39]。
西藏中部湿地植物C含量和δ13C值的变异系数小于N含量和δ15N值的变异系数, 显示出C含量比N含量更具有稳定性, 而湿生植物在一年内经历频繁的干湿交替, 导致土壤中的养分, 尤其是N, 产生较大波动[40]。另一方面, 本研究中湿地植物中C与N元素之间呈现显著正相关关系, 显示出西藏湿地植物在固碳过程中对氮的利用效率的权衡策略与其他研究区域中的植物群体不同。一般认为, 高等陆生植物表现出C与N元素计量之间显著的负相关性, 如滇池流域植物叶片养分[41]和滨海盐地碱蓬叶片[42]的研究。此外, 本研究中湿地植物的N含量、C ∶ N比和δ15N值与含水率之间呈极显著相关性, 这一研究差异说明湿地植物在水的参与下, 其N含量呈现更大的波动性, 从而造成湿地植物的化学计量特征及其对环境变化的响应与陆生植物存在差异[43]。
3.2 湿地植物优势种的生长策略和营养分配模式植物资源利用对策随植物类型及地理分布不同存在较大差异, 而植物本身特性的差异, 也导致研究区不同生活型的湿地植物之间存在显著的资源利用差异(表 3)。本研究的多年生草本植物氮含量高于一年生和半灌木状草本植物, 说明多年生草本植物对N的吸收利用率较高。此外, 湿地植物优势种的碳氮稳定同位素值也显示出其生活型的差异。一年生草本的δ15N值最高, 与祁连山东段高寒草甸植物稳定同位素特性相符[44]。δ13C值呈现出与δ15N值一致的趋势:一年生草本>多年草本>半灌木状草本, δ13C反映了植物长期的水分利用效率[45], 因此本研究发现一年生草本植物的水分利用效率超过多年生草本植物和半灌木状草本植物。不同生活型湿地植物δ13C值的差异在区域尺度上的变化则更多受植物功能性状驱动[35]。
西藏中部河湖岸带不同种类湿地植物C、N含量、C ∶ N比、δ13C和δ15N值均呈现显著差异, 说明植物化学计量特征的差异受到系统发育、植被类型的重要影响。而在不同采样点中, 仅有C ∶ N比、C含量和δ15N值呈现出极显著的空间差异性, 而N含量和δ13C值无显著差异。一方面说明, 气候、土壤等环境因子对于N等限制性元素有一定的影响, 但非决定性因子[46]。另一方面, 由于忽略了不同功能群植物对环境变化的响应差异, 掩盖了N含量和δ13C值在植物功能群中随大尺度环境因子变化的差异[47], 而使其在空间上的生态化学计量学特征受到影响。
本研究的湿地植物C ∶ N比与N含量之间存在显著的负相关性(P < 0.01), 这表明植物的C ∶ N比主要受N含量的影响。湿地植物δ15N值与C ∶ N比呈显著负相关, 与δ13C之间存在显著正相关(P < 0.05), 这与青藏高原东缘草本植物叶片δ15N与C ∶ N比之间和δ13C与δ15N之间关系的研究结果相一致[48], 表明植物对氮素的利用受到植物获取水分能力的影响。干旱环境下, 植物为减少水分蒸腾损失而关闭部分气孔, 导致细胞间CO2浓度降低, δ13C值增加[49], 而由于降水减少, 最终导致植物δ15N增大[50]。因此, δ13C和δ15N值之间的紧密关联可能是由植物所生长环境的水分状况对这两种同位素信号的影响所导致的[51]。总之, 西藏湿地植物在C、N含量及其稳定同位素特征上呈现出与其他植物类群不同的特点, 这可能与其特殊的生境条件以及植物的生活型密切相关。
3.3 地理因子对湿地植物优势种C、N化学计量特征的影响海拔和经纬度梯度综合了温度、光照、水分和土壤等多种环境因子的变化, 直接控制着植物的化学计量特征及其分布格局[16, 52]。经纬度、海拔的改变能够引起温度、降水和蒸散发等气候、土壤及生物类的环境因子变化, 而土壤、水热状况的动态变化特征对植物的发育过程、元素迁移及物质循环具有重要作用[53]。本研究中, 湿地植物含水率与经度、纬度和海拔之间具有显著相关性(P < 0.01), 说明地理梯度改变了当地的水热条件, 而使其含水率发生变化。湿地植物的含水率与海拔梯度和N含量之间呈现正相关关系(P < 0.01), 说明随着海拔升高、N含量增加, 植物的含水率升高, 增强了其抗旱性[54]。相较于低海拔, 高海拔处降水量更少, 而在光照较为充足水分胁迫为主的条件下, 为了减少水分损失, 需要投资更多的N元素用于植物的保护和支撑, 从而降低植物的光合效率和生长速度, 以提高细胞渗透压来增强对水分的保护[55]。同时, 植物多发展出较大的叶厚度、叶组织密度以及较高含水量的性状特征来适应干旱环境, 因此植物具有更高的含水率[54]。
本研究中湿地植物δ15N值与经纬度、海拔均呈显著相关关系(P < 0.05或P < 0.01), 说明地理梯度影响了δ15N值的变化。研究表明, 地理梯度并不会直接对植物δ15N产生影响, 而是通过地理梯度的变化会引起温度、降雨量、大气CO2等环境因素的变化及土壤δ15N和土壤母质的变化, 而影响土壤N循环过程中各种氮源的δ15N值[56], 而最终影响植物中的δ15N值[57]。在地理分布上西藏地区大气降水量呈现出从西北向东南递增的趋势, 而本研究中西藏湿地植物δ15N值的变化呈现与之相反的趋势, 说明研究区湿地植物δ15N值的变化可能受降水影响。已有研究表明, 降水是青藏高原北部高寒草地氮循环最重要的驱动因素[58], 降水通过调控土壤N库[59], 进而对植物的δ15N产生影响。另一方面, 温度和降水也会在不同海拔阶段限制植物的δ15N值[60]。多数研究认为植物的δ15N值随海拔的升高而降低[57];也有研究表明植物的δ15N值在不同的海拔阶段随海拔梯度波动。如, 环青海湖区的植物在海拔2274 m以下其δ15N值呈明显的递减变化, 在海拔3195.83—4639.84 m范围内δ15N值先减小后增大[61]。本研究区域的海拔高度处于2800—4800 m之间, 湿地植物的δ15N值随海拔的升高而呈现缓慢增加趋势(图 4)。因此, 不同海拔阶段限制作用的因子不同[60], 湿地植物δ15N值对地理梯度的变化趋势响应也具有一定的差异性。
4 结论(1) 西藏中部湿地植物优势种的C、N化学计量学特征具有显著差异。C、N含量均值分别为(325.01±83.54)mg/g和(20.76±8.54)mg/g;δ13C和δ15N值的范围分别为-29.15‰—-12.49‰和-0.61‰—24.22‰。C含量大小受到当地非生物环境因素的影响, N含量则与本研究中植物生活型均为草本型植物且处于高海拔、低温度、干旱的环境有关。
(2) 西藏中部湿地植物优势种CN化学计量特征与其特殊的生境条件及植物的生活型密切相关。不同生活型湿地植物N含量和碳氮同位素值的变化说明, 不同生活型的湿地植物之间存在显著的资源利用差异。不同种类、不同生活型、不同样点的湿地植物C、N变量具有显著差异性, 表明植物化学计量特征的差异受到植物种类和植物生活型的影响, 但大尺度上环境条件的变化一定程度上掩盖了植物功能群中碳氮化学计量特征的差异。湿地植物C、N变量之间的相关及回归分析表明, 植物对氮素的利用受到植物获取水分能力的影响。
(3) 湿地植物优势种的C、N变量与含水率及地理因子的相关及回归分析表明, 经纬度、海拔梯度的变化通过水热条件的改变而影响了湿地植物的含水率和化学计量特征。湿地植物δ15N值在地理分布上的变化, 说明其值还可能受到降水的影响, 但不同海拔阶段限制作用的因子不同, 地理梯度变化对其影响也具有一定的差异性。
| [1] |
Zhao Z L, Zhang Y L, Liu L S, Liu F G, Zhang H F. Recent changes in wetlands on the Tibetan Plateau: a review. Journal of Geographical Sciences, 2015, 25(7): 879-896. DOI:10.1007/s11442-015-1208-5 |
| [2] |
Sterner R, Elser J. Ecological stoichiometry: the biology of elements from molecules to the biosphere. Princeton: Princeton University Press, 2003.
|
| [3] |
胡伟芳, 章文龙, 张林海, 陈晓艳, 林伟, 曾从盛, 仝川. 中国主要湿地植被氮和磷生态化学计量学特征. 植物生态学报, 2014, 38(10): 1041-1052. |
| [4] |
杨恒, 张丹, 李桂芳, 叶远行, 陈清飞, 王蓉. 中国河湖岸带草本植物氮磷化学计量学及内稳性特征. 生态科学, 2024, 43(2): 78-86. |
| [5] |
Wang Z, Xia C X, Yu D, Wu Z G. Low-temperature induced leaf elements accumulation in aquatic macrophytes across Tibetan Plateau. Ecological Engineering, 2015, 75: 1-8. DOI:10.1016/j.ecoleng.2014.11.015 |
| [6] |
姚凡云, 朱彪, 杜恩在. δ15N自然丰度法在陆地生态系统氮循环研究中的应用. 植物生态学报, 2012, 36(4): 346-352. |
| [7] |
于贵瑞, 高扬, 王秋凤, 刘世荣, 申卫军. 陆地生态系统碳氮水循环的关键耦合过程及其生物调控机制探讨. 中国生态农业学报, 2013, 21(1): 1-13. |
| [8] |
年世宇, 陈孝杨, 王校刚, 陈敏, 张凌霄. 碳同位素技术在陆地土壤碳循环中的应用. 安徽农业科学, 2020, 48(5): 9-13, 17. DOI:10.3969/j.issn.0517-6611.2020.05.003 |
| [9] |
罗绪强, 王世杰, 张桂玲, 王程媛, 杨鸿雁, 廖昕荣. 钙离子浓度对两种蕨类植物光合作用的影响. 生态环境学报, 2013, 22(2): 258-262. DOI:10.3969/j.issn.1674-5906.2013.02.014 |
| [10] |
O'Leary M H. Carbon Isotopes in Photosynthesis. BioScience, 1988, 38(05): 328-336. DOI:10.2307/1310735 |
| [11] |
殷树鹏, 张成君, 郭方琴, 李晓丽, 张云. 植物碳同位素组成的环境影响因素及在水分利用效率中的应用. 同位素, 2008, 21(1): 46-53. |
| [12] |
温周瑞, 徐军, 谢平. 太湖高等水生植物稳定碳、氮同位素特征. 湖泊科学, 2016, 28(1): 163-169. |
| [13] |
吴云, 刘学炎, 胡朝臣. 城市河流水生植物氮含量和氮同位素记录及其对水体氮污染的响应. 地球与环境, 2021, 49(3): 260-269. |
| [14] |
曾欢欢, 吴骏恩, 刘文杰. 丛林式橡胶林内植物水分利用效率与叶片养分含量. 亚热带植物科学, 2019, 48(2): 125-133. DOI:10.3969/j.issn.1009-7791.2019.02.005 |
| [15] |
Cloern J E, Canuel E A, Harris D. Stable carbon and nitrogen isotope composition of aquatic and terrestrial plants of the San Francisco Bay estuarine system. Limnology and Oceanography, 2002, 47(3): 713-729. DOI:10.4319/lo.2002.47.3.0713 |
| [16] |
拉琼, 扎西次仁, 朱卫东, 许敏, 钟扬. 雅鲁藏布江河岸植物物种丰富度分布格局及其环境解释. 生物多样性, 2014, 22(3): 337-347. |
| [17] |
Jiang M, Yang X, Wang T, Xu Y J, Dong K, He L Y, Liu Y L, Wang J L, Zhao N X, Gao Y B. A direct comparison of the effects and mechanisms between species richness and genotype richness in a dominant species on multiple ecosystem functions. Ecology and Evolution, 2021, 11(20): 14125-14134. |
| [18] |
贺桂芹. 西藏高寒湿地生态系统服务功能价值评估及湿地保护对策研究[D]. 杨凌: 西北农林科技大学, 2007.
|
| [19] |
刘文治, 卢蓓, 刘贵华. 西藏湿地植物. 武汉: 华中科技大学出版社, 2021.
|
| [20] |
Elser J J, Fagan W F, Denno R F, Dobberfuhl D R, Folarin A, Huberty A, Interlandi S, Kilham S S, McCauley E, Schulz K L, Siemann E H, Sterner R W. Nutritional constraints in terrestrial and freshwater food webs. Nature, 2000, 408(6812): 578-580. DOI:10.1038/35046058 |
| [21] |
李培玺. 巢湖湿地典型植物群落植物-土壤碳氮磷生态化学计量学特征[D]. 合肥: 安徽农业大学, 2020.
|
| [22] |
朱秋丽. 闽浙沪滨海湿地草本植物碳氮磷生态化学计量特征[D]. 福州: 福建师范大学, 2017.
|
| [23] |
杨羽. 贵州草海湿地土壤及植物碳氮磷化学计量学研究[D]. 贵阳: 贵州师范大学, 2020.
|
| [24] |
苑鹏云. 内蒙古草原区河流河滨带湿地植物性状对不同环境梯度的响应[D]. 呼和浩特: 内蒙古大学, 2018.
|
| [25] |
刘统高, 谢应忠, 席艳丽, 卜晓燕. 银川平原湿地芦苇碳氮磷分布及其生态化学计量特征研究. 黑龙江畜牧兽医, 2016(19): 22-26, 31. |
| [26] |
黄建军, 王希华. 浙江天童32种常绿阔叶树叶片的营养及结构特征. 华东师范大学学报: 自然科学版, 2003(1): 92-97. DOI:10.3969/j.issn.1000-5641.2003.01.016 |
| [27] |
李冬冬, 方昭, 杜好田, 姚静, 焦峰. 黄土高原不同植被带草本植物叶片与土壤碳氮分布特征. 植物营养与肥料学报, 2019, 25(5): 841-850. |
| [28] |
Reich P B, Oleksyn J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30): 11001-11006. |
| [29] |
Han W X, Fang J Y, Guo D L, Zhang Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytologist, 2005, 168(2): 377-385. DOI:10.1111/j.1469-8137.2005.01530.x |
| [30] |
田地, 严正兵, 方精云. 植物生态化学计量特征及其主要假说. 植物生态学报, 2021, 45(7): 682-713. |
| [31] |
Pan F J, Zhang W, Liu S J, Li D J, Wang K L. Leaf N: P stoichiometry across plant functional groups in the Karst region of southWestern China. Trees, 2015, 29(3): 883-892. DOI:10.1007/s00468-015-1170-y |
| [32] |
Körner, C. The nutritional status of plants from high altitudes. Oecologia, 1989, 81(3): 379-391. DOI:10.1007/BF00377088 |
| [33] |
Yu H L, Fan J W, Harris W, Li Y Z. Relationships between below-ground biomass and foliar N: P stoichiometry along climatic and altitudinal gradients of the Chinese grassland transect. Plant Ecology, 2017, 218(6): 661-671. DOI:10.1007/s11258-017-0719-9 |
| [34] |
Wang W, Zhao J M, Xing Z. Spatial patterns of leaf nitrogen and phosphorus stoichiometry across southeast to central Tibet. Journal of Mountain Science, 2022, 19(9): 2651-2663. DOI:10.1007/s11629-021-7194-4 |
| [35] |
Chen M, Shi Z M, Liu S, Xu G X, Cao X W, Chen J, Zhang M M, Feng Q H, Centritto M, Cao J H. Leaf functional traits have more contributions than climate to the variations of leaf stable carbon isotope of different plant functional types on the eastern Qinghai-Tibetan Plateau. Science of the Total Environment, 2023, 871: 162036. DOI:10.1016/j.scitotenv.2023.162036 |
| [36] |
Kahmen A, Wanek W, Buchmann N. Foliar δ15N values characterize soil N cycling and reflect nitrate or ammonium preference of plants along a temperate grassland gradient. Oecologia, 2008, 156(4): 861-870. DOI:10.1007/s00442-008-1028-8 |
| [37] |
刘贤赵, 张勇, 宿庆, 田艳林, 王庆, 全斌. 陆生植物氮同位素组成与气候环境变化研究进展. 地球科学进展, 2014, 29(2): 216-226. |
| [38] |
Jackson R B, Sperry J S, Dawson T E. Root water uptake and transport: using physiological processes in global predictions. Trends in Plant Science, 2000, 5(11): 482-488. DOI:10.1016/S1360-1385(00)01766-0 |
| [39] |
陈淼, 刘顺, 许格希, 史作民. 土壤剖面碳氮稳定同位素自然丰度的垂直分布模式及其影响机制. 应用生态学报, 2021, 32(6): 1919-1927. |
| [40] |
聂兰琴. 鄱阳湖典型湿地植物碳氮磷化学计量特征研究[D]. 南昌: 江西师范大学, 2015.
|
| [41] |
阎凯, 付登高, 何峰, 段昌群. 滇池流域富磷区不同土壤磷水平下植物叶片的养分化学计量特征. 植物生态学报, 2011, 35(4): 353-361. |
| [42] |
李征, 韩琳, 刘玉虹, 安树青, 冷欣. 滨海盐地碱蓬不同生长阶段叶片C、N、P化学计量特征. 植物生态学报, 2012, 36(10): 1054-1061. |
| [43] |
曾静, 侯志勇, 李旭, 耿明明, 谢永宏. 东洞庭湖洪水前后典型湿地植物化学计量特征动态变化. 环境生态学, 2024, 6(2): 65-70. |
| [44] |
王志鹏, 楚彬, 周睿, 花立民. 祁连山东段高寒草甸常见植物稳定性同位素特征. 甘肃农业大学学报, 2019, 54(1): 175-183, 189. |
| [45] |
Liu Y J, Zhang L R, Niu H S, Sun Y, Xu X L. Habitat-specific differences in plasticity of foliar δ13C in temperate steppe grasses. Ecology and Evolution, 2014, 4(5): 648-655. DOI:10.1002/ece3.970 |
| [46] |
Han W X, Fang J Y, Reich P B, Ian Woodward F, Wang Z H. Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in China. Ecology Letters, 2011, 14(8): 788-796. DOI:10.1111/j.1461-0248.2011.01641.x |
| [47] |
于海玲, 李愈哲, 樊江文, 钟华平. 中国草地样带不同功能群植物叶片氮磷含量随水热因子的变化规律. 生态学杂志, 2016, 35(11): 2867-2874. |
| [48] |
陈淼, 刘顺, 许格希, 陈健, 邢红爽, 李非凡, 张淼淼, 曹向文, 史作民. 青藏高原东缘不同植被类型下草本植物叶片碳氮稳定同位素差异及其驱动因素. 应用生态学报, 2024, 35(4): 877-885. |
| [49] |
Farquhar G D, Ehleringer J R, Hubick K T. Carbon isotope discrimination and photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology, 1989, 40: 503-537. DOI:10.1146/annurev.pp.40.060189.002443 |
| [50] |
Austin A, Sala O, Schulze E, Farquhar G, Miller J, Schulze W, Walker B, Williams R. Foliar δ15N is negatively correlated with rainfall along the IGBP transect in Australia. Australian Journal of Plant Physiology, 1999, 26(3): 293-295. |
| [51] |
Peri P L, Ladd B, Pepper D A, Bonser S P, Laffan S W, Amelung W. Carbon (δ13C) and nitrogen (δ15N) stable isotope composition in plant and soil in Southern Patagonia's native forests. Global Change Biology, 2012, 18(1): 311-321. DOI:10.1111/j.1365-2486.2011.02494.x |
| [52] |
Gaston K J. Global patterns in biodiversity. Nature, 2000, 405(6783): 220-227. DOI:10.1038/35012228 |
| [53] |
卢同平, 王艳飞, 王黎明, 林永静, 武梦娟, 张文翔, 牛洁. 西双版纳热带雨林土壤与叶片生态化学计量特征的干湿度效应. 生态学报, 2018, 38(7): 2333-2343. |
| [54] |
王剑博, 单立山, 解婷婷, 杨彪生, 师亚婷, 何彩. 环境异质性对红砂叶片功能性状的影响. 草地学报, 2023, 31(4): 1048-1056. |
| [55] |
周扬, 张丹桔, 宋思梦, 李勋, 张艳, 张健. 林窗大小对马尾松林下3种更新草本植物N、P化学计量研究. 植物研究, 2017, 37(6): 915-925. |
| [56] |
宋大伟, 李明财, 李来兴, 易现峰, 张晓爱. 高寒草甸消费者种群稳定碳、氮同位素组成的海拔分异. 生态学杂志, 2007, 26(1): 40-45. |
| [57] |
周咏春, 程希雷, 樊江文. 植物氮同位素组成与其影响因子的关系研究进展. 草地学报, 2012, 20(6): 981-989. |
| [58] |
Wu J S, Song M H, Ma W L, Zhang X Z, Shen Z X, Tarolli P, Wurst S, Shi P L, Ratzmann G, Feng Y F, Li M, Wang X T, Tietjen B. Plant and soil's δ15N are regulated by climate, soil nutrients, and species diversity in alpine grasslands on the northern Tibetan Plateau. Agriculture, Ecosystems & Environment, 2019, 281: 111-123. |
| [59] |
Lin L, Zhu B, Chen C R, Zhang Z H, Wang Q B, He J S. Precipitation overrides warming in mediating soil nitrogen pools in an alpine grassland ecosystem on the Tibetan Plateau. Scientific Reports, 2016, 6: 31438. DOI:10.1038/srep31438 |
| [60] |
刘贤赵, 王国安, 李嘉竹, 王庆. 北京东灵山地区现代植物氮同位素组成及其对海拔梯度的响应. 中国科学: D辑: 地球科学, 2009, 39(10): 1347-1359. |
| [61] |
李雪双, 贺世杰, 金秉福, 王传远. 环青海湖区域植物稳定碳氮同位素空间分布规律及影响因素研究. 鲁东大学学报: 自然科学版, 2018, 34(2): 167-172. DOI:10.3969/j.issn.1673-8020.2018.02.011 |